CHAPTER 2 Surgical Anatomy of the Brain
Cerebrum
Lateral Surface: Neural Structures
Superficial Anatomy
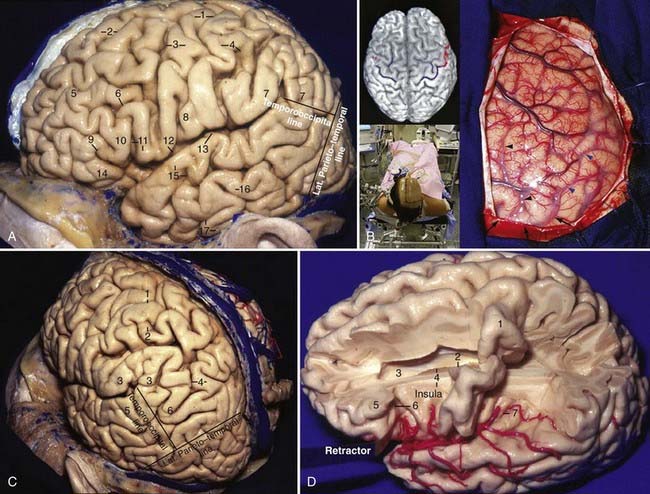
The cerebrum is arbitrarily divided into five lobes: frontal, parietal, temporal, occipital, and the hidden insula. On the lateral surface, they are limited by the central sulcus, the posterior ramus of the sylvian fissure, the lateral parietotemporal line (from the impression of the parieto-occipital sulcus to the preoccipital notch), and the temporo-occipital line (from the posterior end of the posterior ramus of the sylvian fissure to the midpoint of the lateral parietotemporal line). The cerebrum has four main sulci that are 100% continuous—the sylvian fissure and the callosal, parieto-occipital, and collateral sulci—and two almost continuous (92%) sulci—the central and calcarine sulci. There are two 100% interrupted sulci: the precentral and inferior temporal sulci.1 The central sulcus starts from the medial surface of the hemisphere above the cingulate sulcus and extends on the lateral surface of the hemisphere in a medial-to-lateral, superior-to-inferior, and posterior-to-anterior direction. It does not usually intercept the posterior ramus of the sylvian fissure and leaves a “bridge” connecting the precentral to the postcentral gyrus, known as pli de passage frontoparietal inferior, opercule rolandique, or the subcentral gyrus (Fig. 2-1A).
Frontal Lobe
The two main sulci are the superior and inferior frontal sulci, which are anteroposteriorly oriented and extend from the precentral sulcus to the frontal pole. At their posterior end, these two sulci are intercepted perpendicularly by the precentral sulcus, which has a direction very similar to that of the central sulcus. The precentral sulcus forms the anterior limit of the precentral gyrus. These two frontal sulci divide the lateral surface of the frontal lobe into three gyri: the superior, middle, and inferior frontal gyri (Fig. 2-1A). The anterior horizontal, the anterior ascending, and the posterior rami of the sylvian fissure divide the inferior frontal gyrus into three parts: the pars orbitalis, triangularis, and opercularis. The apex of the pars triangularis is usually retracted superiorly and leaves a space in the sylvian fissure that is generally the largest space in the superficial compartment of the sylvian fissure. The apex of the pars triangularis is directed inferiorly toward the junction of three rami of the sylvian fissure; this junctional point coincides with the anterior limiting sulcus of the insula in the depth of the sylvian fissure. It marks the anterior limit of the basal ganglia and the location of the anterior horn of the lateral ventricle. At the intercepting point between the superior frontal and precentral sulci, the precentral gyrus often has the morphology of the Greek letter “Ω” (omega), with its convexity pointing posteriorly. This is the most easily identifiable landmark of the motor strip and corresponds to the hand area (Fig. 2-1B).
Parietal Lobe
The parietal lobe is limited anteriorly by the central sulcus, medially by the interhemispheric fissure, inferolaterally by the sylvian fissure and the temporo-occipital line, and posteriorly by the lateral parietotemporal line. Its two main sulci are the postcentral and intraparietal sulci. The postcentral sulcus is very similar to the central sulcus, except for its variable continuity. The postcentral sulcus is the posterior limit of the postcentral gyrus, and it can sometimes be double. The intraparietal sulcus starts at the postcentral sulcus and is directed posteriorly and inferiorly toward the occipital pole; its direction is often parallel and 2 to 3 cm lateral to the midline. The bottom of the intraparietal sulcus is related to both the roof of the atrium and the occipital horn. The intraparietal sulcus divides the lateral surface of the parietal lobe into two parts: the superior and inferior parietal lobules. The superior parietal lobule, which is the superomedial and smaller part, continues as the precuneus on the medial surface of the parietal lobe. The inferior parietal lobule is constituted by the supramarginal and angular gyri. The supramarginal gyrus, the posterior continuation of the superior temporal gyrus, turns around the posterior ascending ramus of the sylvian fissure. The angular gyrus is the posterior continuation of the middle temporal gyrus and turns superiorly and medially behind the posterior ramus of the sylvian fissure up to the intraparietal sulcus; it is sometimes limited between the two posterior terminations of the superior temporal sulcus, the angular and anterior occipital rami (Fig. 2-1C).
Temporal Lobe
The temporal lobe is limited superiorly by the posterior ramus of the sylvian fissure and posteriorly by the temporo-occipital and lateral parietotemporal lines. It has two main sulci, the superior and inferior temporal sulci, that divide the lateral surface of the temporal lobe into three gyri, the superior, middle, and inferior temporal gyri. The inferior temporal gyrus occupies the lateral and basal surfaces of the cerebrum. The superior and inferior temporal gyri converge anteriorly to form the temporal pole (Fig. 2-1A).
Occipital Lobe
The “x-ray” vision concept can be demonstrated by the precentral gyrus, which begins on the medial surface of the cerebrum, above the level of the splenium of the corpus callosum, and passes above the body of the lateral ventricle, thalamus, posterior limb of the internal capsule, and posterior part of the lentiform nucleus to reach the sylvian fissure approximately midway between the anterior and posterior limits of the insula (Fig. 2-1D).
Sylvian Fissure
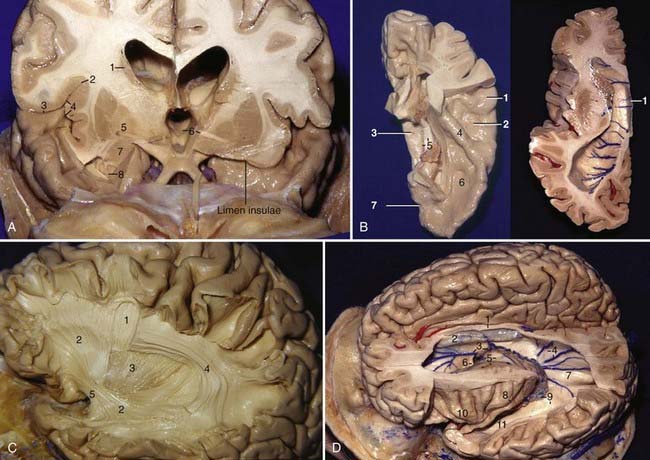
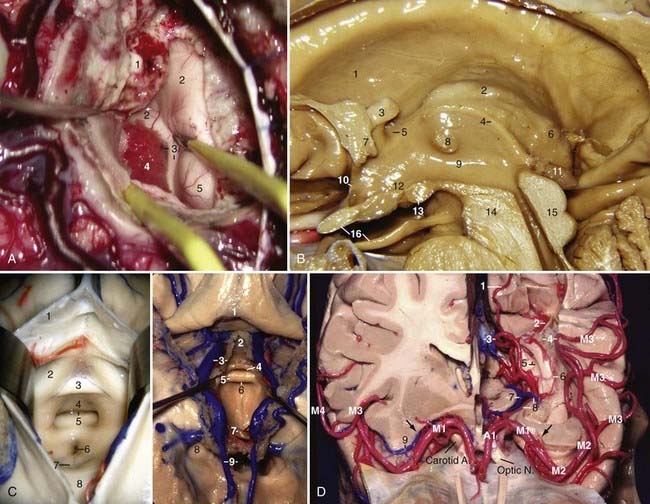
The sylvian fissure is the space between the frontal, parietal, and temporal opercula and the insula and extends from the basal to the lateral surface of the brain. It is composed of a superficial and a deep part. The superficial part has a stem and three rami; the stem extends medially from the semilunar gyrus of the uncus to the lateral end of the sphenoid ridge, where the stem divides into the anterior horizontal, anterior ascending, and posterior rami (Fig. 2-1A). The deep part is divided into a “sphenoidal compartment” and an “operculoinsular compartment.” The sphenoidal compartment, which arises in the region of the limen insulae lateral to the anterior perforated substance (APS), is a narrow space posterior to the sphenoid ridge between the frontal and temporal lobes that communicates medially with the carotid cistern, also called sylvian vallecula (see Fig. 2-4D).2 The operculoinsular compartment is formed by two narrow clefts, the opercular cleft between the opposing lips of the frontoparietal and temporal opercula and the insular cleft, which has a superior limb located between the insula and the frontoparietal opercula and an inferior limb between the insula and the temporal operculum (Fig. 2-2A).3 The gyri that constitute the frontal and parietal opercula of the sylvian fissure are, from posterior to anterior, the supramarginal, postcentral, and precentral gyri and the pars opercularis, triangularis, and orbitalis (see Fig. 2-1A); the gyri that constitute the temporal operculum of the sylvian fissure are, from posterior to anterior, the planum temporale, Heschl’s gyrus, and the planum polare (Fig. 2-2B, left). Each gyrus of the frontoparietal operculum is related to its counterpart on the temporal side; the supramarginal gyrus is in contact with the planum temporale, the postcentral gyrus is in contact with Heschl’s gyrus, and the precentral gyrus and pars opercularis, triangularis, and orbitalis are related to the planum polare. The site on the posterior ramus of the sylvian fissure where the postcentral gyrus meets Heschl’s gyrus is projected in the same coronal plane as the external acoustic meatus. The medial wall of the sylvian fissure is the insula or island of Reil, which can be seen only when the lips of the sylvian fissure are widely separated. The insula has the shape of a pyramid with its apex directed inferiorly and has an anterior and a lateral surface. The anterior surface is triangular in shape and is constituted by the transverse and accessory gyri and the insular pole. The medial portion of the insular pole is marked by an arched ridge of variable prominence, the limen insulae, which is composed of fibers of the uncinate fasciculus covered by a thin layer of gray matter that extends from the anterior end of the long gyrus, passes through the medial part of the insular pole, and ends at the middle of the posterior orbital gyrus. “Limen” means threshold, and the limen insulae is the threshold between the carotid cistern medially and the sylvian fissure laterally (Fig. 2-2A). The insula is encircled and separated from the opercula by a deep furrow called the circular or limiting sulcus of the insula, which has three parts, the superior, anterior, and inferior parts (see Fig. 2-1D). From the limen insulae, the sulci and gyri of the insula are directed superiorly in a radial manner. The deepest sulcus, the central sulcus of the insula, is a constant sulcus that extends upward and backward across the insula, in the general line of the central sulcus of the cerebrum. It divides the lateral surface of the insula into a large anterior zone that is divided by several shallow sulci into three to five short gyri and a posterior zone that is formed by the anterior and posterior long gyri (Fig. 2-2D). From microsurgical and radiologic viewpoints, the insula represents the external covering of the central core and is constituted by the extreme, external, and internal capsules, the claustrum, the basal ganglia, and the thalamus (Fig. 2-3A, left; also see Fig 2-2A). The anterior, inferior, and posterior limits of the insula on the lateral projection correspond to the anterior, inferior, and posterior limits of the central core. The upper limit of the central core (caudate nucleus) is higher than the upper limit of the insula (Fig. 2-2A).
Association Fibers of the Cerebrum
The association fibers are tracts of myelinated fibers that connect cortical areas of different lobes in the same hemisphere; they may be divided into short and long association fibers. The short association fibers connect adjacent gyri, whereas the long association fibers (fasciculi) connect distant gyri and form distinct compact bundles. The main fasciculi are (1) the superior longitudinal fasciculus, which is the largest, arches around the insula, and connects parts of the frontal, parietal, and temporal lobes; (2) the uncinate fasciculus, which lies in the depth of the limen insulae, has a marked curvature, and connects the basal parts of the frontal lobe with the temporal lobe; (3) the inferior occipitofrontal fasciculus, which connects the frontal and occipital lobes, as well as the posterior part of the temporal and parietal lobes; these fibers converge from the frontal lobe as a single bundle that runs lateral to the lentiform nucleus, where they are closely associated with the uncinate fasciculus (Fig. 2-2C); and (4) the cingulum, the fibers of which lie within the cingulate gyrus from below the rostrum of the corpus callosum to the parahippocampal gyrus.
Lateral Ventricles
Wrapping around the central core of the hemisphere are the lateral ventricles (Fig. 2-2D). Each ventricle has five components: a frontal horn, body, atrium, and occipital and temporal horns.4 The frontal horn is located in front of the foramen of Monro and has a roof, floor, and anterior, lateral, medial, and posterior walls. The transition between the genu and the body of the corpus callosum forms the roof, the rostrum of the corpus callosum forms the narrow floor, the septum pellucidum forms the medial wall, and the thalamus forms the posterior wall. The head of the caudate nucleus forms the majority of the lateral wall, but the most anterior part is constituted by the most anterior portion of the anterior limb of the internal capsule, and it is in close relation to the anterior limiting sulcus of the insula. The body of the lateral ventricle is located behind the foramen of Monro and extends to the point where the septum pellucidum, corpus callosum, and fornix meet. It has a roof, floor, and lateral and medial walls. The body of the corpus callosum forms the roof, the septum pellucidum above and the body of the fornix below form the medial wall, the body of the caudate nucleus forms the lateral wall, and the thalamus forms the floor. The caudate nucleus and the thalamus are separated by the striothalamic sulcus, the groove in which the stria terminalis and the thalamostriate vein course. The atrium has a roof, floor, and anterior, medial, and lateral walls. The roof is formed by the body, splenium, and tapetum of the corpus callosum. The floor is formed by the collateral trigone, a triangular area that bulges upward over the posterior end of the collateral sulcus. The medial wall is formed by two roughly horizontal prominences: the upper prominence, or the bulb of the callosum, is formed by a large bundle of fibers called the forceps major that connects the two occipital lobes; the lower prominence, or the calcar avis, overlies the deepest part of the calcarine sulcus. The lateral wall has an anterior portion formed by the caudate nucleus as it wraps around the lateral margin of the pulvinar, as well as a posterior portion formed by the fibers of the tapetum as they sweep anteroinferiorly along the lateral margin of the ventricle and separate the ventricular cavity from the optic radiation. The anterior wall has a medial part composed of the crus of the fornix as it wraps around the posterior portion of the pulvinar and a lateral part formed by the pulvinar of the thalamus. The occipital horn extends posteriorly into the occipital lobe from the atrium. It varies in size from being absent to extending far posterior in the occipital lobe. The bulb of the callosum and the calcar avis form its medial wall, the tapetum forms the roof and the lateral wall, and the collateral trigone forms the floor (Fig. 2-3A and B).4 The temporal horn extends forward and inferiorly from the atrium into the medial part of the temporal lobe and has a roof, floor, and anterior, lateral, and medial walls. The tapetum, the tail of the caudate nucleus, part of the retrolentiform and sublentiform components of the internal capsule, and the amygdaloid nucleus form the roof. The retrolentiform component is the posterior thalamic radiation that includes the optic radiation (Fig. 2-3C); the sublentiform component is formed mainly by the acoustic radiation. The amygdaloid nucleus constitutes the most anterior portion of the roof of the temporal horn and is located above and in front of the head of the hippocampus (Fig. 2-3D, right), anterior to the inferior choroidal point, which is the most anterior site of attachment of the choroid plexus in the temporal horn.5 There is no clear separation between the roof of the temporal horn and the thalamus because all fibers of the optic radiation come from the lateral geniculate body. Therefore, it is reasonable to consider the roof of the temporal horn a lateral extension of the thalamus.5 The attachment site of the choroid plexus can be a surgical landmark to separate the thalamus from the roof of the temporal horn (see Fig. 2-6D, right). The tapetum and the optic radiation form the lateral wall, the amygdaloid body forms the anterior wall, the head of the hippocampus forms the anterior third of the medial wall, and the choroidal fissure forms the posterior two thirds of the medial wall.5 The floor is formed medially by the hippocampus and laterally by the collateral eminence (Fig. 2-3D, left). The temporal horn is projected onto the middle temporal gyrus on the lateral view.
Foramen of Monro
The foramen of Monro is a passage through which the lateral ventricle communicates with the third ventricle. It usually has a crescent shape and is bounded anteriorly and superiorly by the columns of the fornix and posteriorly by the thalamus6; the elements that run close to the foramen of Monro are the anterior septal vein superiorly and medially, the choroidal plexus posterior and medially, and the thalamostriate vein laterally and posteriorly (see Figs. 2-2D and 2-3A).
Internal Capsule
The internal capsule has five parts: the anterior and posterior limbs, the genu, and the retrolentiform and sublentiform parts. The anterior limb is located between the head of the caudate nucleus and the anterior half of the lentiform nucleus and contains frontopontine fibers; the posterior limb is located between the thalamus and the posterior half of the lentiform nucleus and contains corticospinal tract, frontopontine, and corticorubral fibers and fibers of the superior thalamic radiation (somesthetic radiation). The genu comes to the ventricular surface immediately lateral to the foramen of Monro in the interval between the caudate nucleus and the thalamus, where the thalamostriate vein usually drains into the internal cerebral vein; the genu contains corticonuclear fibers and anterior fibers of the superior thalamic radiation. The retrolentiform part is located posterior to the lentiform nucleus and contains mainly parietopontine, occipitopontine, occipitocollicular, and occipitotectal fibers and the posterior thalamic radiation that includes the optic radiation. The sublentiform part is located below the lentiform nucleus and contains temporopontine and parietopontine fibers and acoustic radiation from the medial geniculate body to the superior temporal gyrus and the transverse temporal gyri (Fig. 2-3A, left, and B).
Corpus Callosum
The corpus callosum is the largest transverse commissure connecting the cerebral hemispheres. It contributes to the wall of each of the five parts of the lateral ventricle (see Fig. 2-2B, right). The corpus callosum is divided in four parts: rostrum, genu, body, and splenium. The rostrum is the floor of the frontal horn. The genu gives rise to a large fiber tract, the forceps minor, that forms the anterior wall of the frontal horn, and it connects the frontal lobes. The splenium gives rise to a large tract, the forceps major, that forms a prominence called the bulb in the upper part of the medial wall of the atrium and occipital horn as it sweeps posteriorly to connect the occipital lobes. Another fiber tract, the tapetum, arises in the posterior part of the body and splenium and sweeps laterally and inferiorly to form the roof and lateral wall of the atrium and the temporal and occipital horns.
Optic Radiation
The optic radiation is a bundle of fibers that extend from the lateral geniculate body to the visual area in the occipital lobe. The optic radiation may be divided into three parts: anterior, middle, and posterior. In the anterior part, the fibers initially take an anterior direction along the roof of the temporal horn, usually reach as far anteriorly as the tip of the temporal horn, and then loop backward in the lateral and inferior aspects of the atrium and the occipital horn to end in the lower lip of the calcarine fissure; this anterior loop is called Meyer’s loop. The anterior part represents the upper quadrants of the visual field. In the middle part, the fibers take a lateral direction initially, course along the roof of the temporal horn, and then proceed posteriorly along the lateral wall of the atrium and the occipital horn; the middle part contains the macular fibers. The fibers of the posterior part course directly backward along the lateral wall of the atrium and the occipital horn to end in the upper lip of the calcarine fissure; these fibers are responsible for the lower quadrants of the visual field (Fig. 2-3C).
Fornix
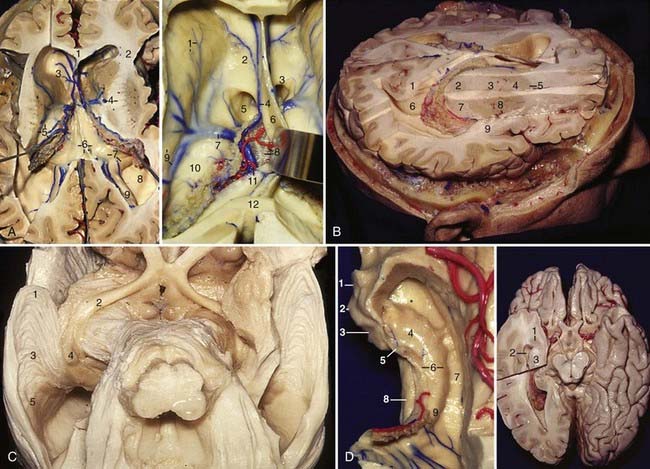
The fornix is a C-shaped structure that wraps around the thalamus in the wall of the lateral ventricle. The initial portion of the fornix, the fimbria, arises from the alveus, which is the subcortical white matter of the hippocampal allocortex, and thickens along the medial edge of the hippocampus; it is separated from the dentate gyrus by the fimbriodentate sulcus. The fimbria then passes posteriorly to become the crus of the fornix, which is the subcortical radiation of the hippocampal allocortex. In the atrium the crus wraps around the posterior surface of the pulvinar of the thalamus and arches superomedially toward the lower surface of the splenium of the corpus callosum; at the junction between the atrium and body of the lateral ventricle, the paired crura meet to form the body of the fornix. At the anterior margin of the thalamus, the body of the fornix separates into two columns that arch along the superior and anterior margins of the foramen of Monro. The columns of the fornix then split, pass predominantly posterior to the anterior commissure, and are directed inferiorly and posteriorly through the lateral wall of the third ventricle to reach the mamillary bodies at the floor of the third ventricle. In the area below the splenium, the two crura of the fornix are united by the hippocampal commissure (Fig. 2-4A; also see Fig. 2-3A and D, left).
Basal Ganglia
The caudate nucleus is a C-shaped structure that wraps around the thalamus; it has a head, body, and tail. The head and the body are the lateral walls of the frontal horn and the body of the lateral ventricle. The tail extends from the atrium into the roof of the temporal horn and is continuous with the amygdaloid nucleus (see Figs. 2-2A and 2-3A).
Thalamus
The thalamus is located in the center of the lateral ventricle. Each lateral ventricle wraps around the superior, inferior, and posterior surfaces of the thalamus. The anterior tubercle of the thalamus is the posterior limit of the foramen of Monro; the posterior part, called the pulvinar (pillow) of the thalamus, is the wall of three different compartments in the cerebrum. The posterolateral part of the pulvinar is the lateral half of the anterior wall of the atrium, the posteromedial part is covered by the crus of the fornix and is part of the superolateral wall of the quadrigeminal cistern, and the inferolateral part of the pulvinar is the roof of the wing of the ambient cistern. The medial part of the thalamus is the lateral wall of the third ventricle (see Figs. 2-3A and 2-4B).
Hippocampus
The hippocampus occupies the medial portion of the floor of the temporal horn and is divided into three parts: head, body, and tail. The head of the hippocampus, the anterior and largest part, is directed anteriorly and inferiorly and then medially. At the medial end of the tip of the temporal horn, it turns up vertically and bends over laterally to form the medial wall of the tip of the temporal horn, ahead of the choroidal fissure. The head of the hippocampus is free of the choroid plexus and features three or four hippocampal digitations; its overall shape resembles a feline paw, and it is directed toward the posterior segment of the uncus. Its posterior limit is the initial segment of the fimbria and the choroidal fissure. Superiorly, the head of the hippocampus is related to the posteroinferior portion of the amygdala. Anteriorly, it is related to the uncal recess of the temporal horn, which is the anterior continuation of the collateral eminence. The emergence of the choroid plexus, fimbria, and choroidal fissure marks the beginning of the body of the hippocampus. The body of the hippocampus takes an anteroposterior and inferosuperior direction and narrows as it approaches the atrium of the lateral ventricle. Posterior to the head of the hippocampus, the medial wall of the temporal horn is the choroidal fissure. At the atrium of the lateral ventricle, the body of the hippocampus changes direction and has its longitudinal axis oriented transversely to become the tail of the hippocampus. The tail of the hippocampus is slender and constitutes the medial part of the floor of the atrium; medially, the tail of the hippocampus fuses with the calcar avis. Histologically, the terminal segment of the hippocampal tail continues as the subsplenial gyrus, which covers the inferior splenial surface (see Figs. 2-3D, left, and 2-4A).
Amygdala
The amygdala and the hippocampus constitute the core of the limbic system. The temporal amygdala is composed of a series of gray matter nuclei classified into three main groups: basolateral, corticomedial, and central. From a neurosurgical viewpoint, the temporal amygdala can be considered to be located entirely within the boundaries of the uncus: superiorly, the amygdala blends into the globus pallidus; inferiorly, the temporal amygdala bulges inferiorly into the most anterior portion of the roof of the temporal horn above the hippocampal head and the uncal recess; and medially, it is related to the anterior and posterior segments of the uncus. It also forms the anterior wall of the temporal horn (see Figs. 2-2A, 2-3D, right, and 2-4A).
Choroidal Fissure
The choroidal fissure is a cleft located between the thalamus and the fornix and is the site of attachment of the choroid plexus in the lateral ventricle. It is a C-shaped arc that extends from the foramen of Monro through the body and atrium to the temporal horn.7 The body portion of the choroidal fissure lies between the body of the fornix and the thalamus,8 the atrial portion is located between the crus of the fornix and the pulvinar of the thalamus (see Fig. 2-3A), and the temporal portion lies between the fimbria of the fornix and the stria terminalis of the thalamus. The choroid plexus is attached to the fornix and the thalamus by an ependymal covering called the taenia fornicis and taenia choroidea, respectively; in the temporal part, the taenia fimbriae attaches the choroid plexus to the fimbria. The choroidal fissure is one of the most important landmarks in microneurosurgery involving the temporal lobe in that it separates temporal structures that can be removed from thalamic structures that should be preserved (Fig. 2-4A).
Third Ventricle
The third ventricle is a narrow, funnel-shaped, unilocular midline cavity. It communicates at its anterosuperior margin with each lateral ventricle through the foramen of Monro and posteriorly with the fourth ventricle through the aqueduct of Sylvius (Fig. 2-4B). It has a roof, a floor, and an anterior, posterior, and two lateral walls.9 The roof extends from the foramen of Monro anteriorly to the suprapineal recess posteriorly and is constituted superiorly to inferiorly by five layers (see Fig. 2-3A). The first layer is the fornix; the body of the fornix is the anterior portion of the roof of the third ventricle, and the crura and the hippocampal commissure are the roof of the posterior portion. The second layer is the superior membrane of the tela choroidea, which is the part of the tela choroidea that passes thorough the forniceal side of the choroidal fissure to cover the choroid plexus of the lateral ventricle. The third layer is a vascular layer located in a space between the superior and inferior membranes of the tela choroidea called the velum interpositum; it contains the internal cerebral veins and branches of the medial posterior choroidal arteries. The fourth layer, the inferior membrane of the tela choroidea, forms the floor of the velum interpositum. It is attached anterolaterally to the taenia thalami, a small ridge on the free edge of a fiber tract, the striae medullaris thalami, that extends along the superomedial border of the thalamus from the foramen of Monro to the habenular commissure (Fig. 2-4B). The posterior part of the inferior membrane of the tela choroidea is attached to the superior surface of the pineal body. The fifth layer is the choroidal plexus of the third ventricle and is usually represented by two parallel strands of choroid plexus projecting backward on each side of the midline. The floor extends from the optic chiasm, anteriorly, to the orifice of the aqueduct of Sylvius posteriorly, and it is constituted, from anterior to posterior, by the optic and infundibular recesses, the tuber cinereum, the mamillary bodies, the posterior perforated substance, the midbrain, and the aqueduct (Fig. 2-4B). The anterior wall is formed by the lamina terminalis and the posterior wall is represented, from inferior to superior, by the posterior commissure, pineal recess, habenular commissure, pineal gland, and suprapineal recess (Fig. 2-4C). At the inner angle formed by the roof and the anterior wall is the anterior commissure.10 Frequently, there is another commissure in the cavity of the third ventricle located posterior to the foramen of Monro called the massa intermedia, which connects both thalami. The lateral wall of the third ventricle is constituted by the thalamus above and by the hypothalamus below, both separated by the hypothalamic sulcus, a shallow groove extending from the foramen of Monro to the aqueduct. The hypothalamic sulcus is the rostral continuation of the sulcus limitans of the brainstem (Fig. 2-4B).
Lateral Surface: Arterial Relationships
The middle cerebral artery (MCA) is divided into four segments: the M1 or sphenoidal segment extends from the bifurcation of the internal carotid artery (ICA) to the limen insulae and is discussed in the section on the basal surface.11,12 The M2 or insular segment extends from the limen insulae to the superior and inferior circular sulci of the insula; it runs in the insular compartment of the sylvian fissure and is constituted by the superior and inferior trunks and their branches. After reaching the superior or inferior circular sulcus of the insula, the M2 branches enter the opercular compartment and are called the M3 segment. The M3 or opercular segment runs in the opercular compartment and is related to the frontal and parietal opercula superiorly and to the temporal operculum inferiorly. The loop of the most posterior M3 segment branch that exits from the sylvian fissure is called the M point or the sylvian point.13 Anatomically, the sylvian point is located behind the insula, above the medial end of Heschl’s gyrus (Fig. 2-4D). The angiographic sylvian point or M point is the location of the medial end of Heschl’s gyrus, the posterior end of the insula, and the central core, atrium, and pulvinar of the thalamus (Fig. 2-13A). On a lateral projection, the M2 and M3 segments form the “sylvian triangle,” which depicts the shape of the insula and represents the anterior, inferior, and posterior limits of the central core (Figs. 2-5A and 2-13B). The caudate nucleus is projected above the superior level of the sylvian triangle on a lateral projection (Figs. 2-5A and 2-13B). The fourth segment is the M4 or cortical segment; it extends from the sylvian fissure to the lateral surface of the cerebrum.
Lateral Surface: Venous Relationships
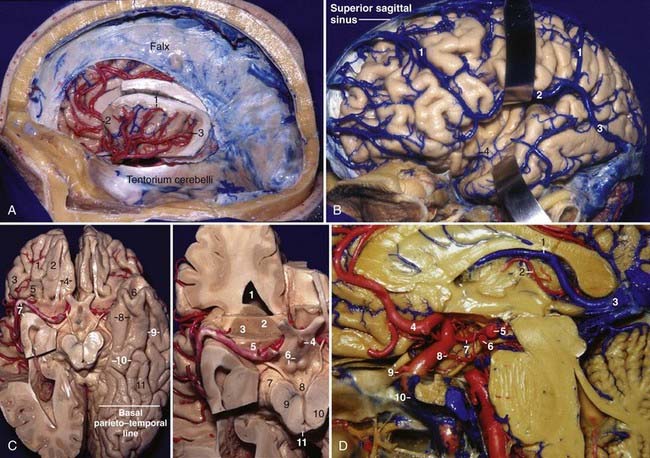
The superficial venous system drains the superficial fifth of the thickness of the cerebrum, whereas the deep venous system drains the remaining four fifths of the depth of the cerebrum. On the lateral surface of the cerebrum, the superficial venous drainage system is directed to venous channels adjacent to the lobes. On the frontal and parietal lobes, venous drainage may be directed superiorly toward the superior sagittal sinus or inferiorly toward the superficial sylvian vein. On the temporal lobe, the veins can drain superiorly toward the superficial sylvian vein or inferiorly toward the dural sinuses below the temporal lobe.14 There are three main anastomotic veins on the lateral surface of the cerebrum. The superficial sylvian vein begins at the posterior part of the posterior ramus of the sylvian fissure, runs inferiorly and anteriorly along the fissure, and commonly anastomoses with the veins of Trolard and Labbé. It may arise as two trunks or have several variations. In the region of the pterion, it enters the dura, runs along the lesser wing of the sphenoid in the sphenoparietal sinus or sinus of the lesser wing of the sphenoid, enters the anterior end of the cavernous sinus via the medial end of the superior orbital fissure, and then drains into the basilar sinus and the inferior petrosal sinus. The vein of Trolard, or the superior anastomotic vein, is the largest anastomotic vein crossing the lateral surface of the brain between the superior sagittal sinus and the sylvian fissure. It is more frequently located at the parietal lobe. The vein of Labbé, or the inferior anastomotic vein, is the largest anastomotic vein that crosses the temporal lobe between the sylvian fissure and the transverse sinus. It usually arises from the middle portion of the sylvian fissure and is directed posteriorly and inferiorly toward the anterior part of the transverse sinus, at the level of the preoccipital notch (Fig. 2-5B).
The deep part of the sylvian fissure is related to the deep sylvian or middle cerebral vein and its tributaries. The tributaries of the deep sylvian vein come mainly from the sulci of the insula. The deep middle cerebral vein begins as a vein in the central sulcus of the insula and runs anteriorly and inferiorly toward the limen insulae, where it joins other insular veins to form a common trunk (Fig. 2-6B).15
The deep venous system is divided into ventricular and cisternal groups; the cisternal group is discussed in the section on the basal surface. The ventricular veins are named mainly according to the location where they course: frontal horn veins—anterior caudate and anterior septal veins; veins of the body of the lateral ventricle—thalamostriate, thalamocaudate, posterior caudate, and posterior septal veins; atrium and occipital horn veins—medial and lateral atrial veins; temporal horn veins—inferior ventricular, amygdalar, and transverse hippocampal veins; deep thalamic veins—anterior and superior thalamic veins; superficial thalamic veins—anterior, superior, and posterior superficial thalamic veins; and choroidal veins—superior and inferior choroidal veins.15
Basal Surface: Neural Relationships
The basal surface is composed of the frontal, temporal, and occipital lobes. The olfactory tract and sulcus divide the basal surface of the frontal lobe into two uneven parts: the smaller and medial part is the rectus gyrus, whereas the larger and lateral part, the orbital surface, is located above the orbit and is composed of the orbital gyri. The orbital surface is divided by the orbital sulcus, a complex sulcus that exhibits a rough configuration of the letter “H,” into four quadrants: the anterior, medial, posterior, and lateral orbital gyri. The pars orbitalis of the inferior frontal gyrus is continuous with the posterior part of the lateral orbital gyrus and with the lateral part of the posterior orbital gyrus. The temporal lobe is separated posteriorly from the occipital lobe by the basal parietotemporal line (from the preoccipital notch to the junction between the parieto-occipital and calcarine fissures) and, laterally to medially, is composed of the inferior temporal gyrus, occipitotemporal sulcus, fusiform gyrus, collateral sulcus, and parahippocampal gyrus (Fig. 2-5C, left). The collateral sulcus is oriented inferiorly to superiorly and medially to laterally and bulges into the lateral part of the floor of the temporal horn (collateral eminence) and the atrium (collateral trigone). The collateral sulcus separates the allocortical parahippocampal gyrus medially from the mesocortical fusiform gyrus laterally. These gyri are kept separated anteriorly by the rhinal sulcus, which separates the uncus medially from the temporal pole laterally. The rhinal sulcus can be considered an anterior continuation of the collateral sulci, and it continues superiorly on the surface of the planum polare and separates it from the uncus medially (Figs. 2-2A and 2-5C, left).
The interpeduncular region is bounded by two oculomotor nerves and the posteromedial surface and apex of the uncus laterally; by the diencephalic membrane of the Liliequist membrane (the membrane that extends from the dorsum sellae to the mamillary bodies), pituitary stalk, and dorsum sellae anteriorly; by the tuber cinereum, mamillary bodies, and the posterior perforated substance superiorly; and by the inner surface of both crura cerebri posteriorly. The prepontine cistern forms the inferior limit of the interpeduncular fossa (Fig. 2-5C and D).
Anterior Perforated Substance
The APS is the entry site for perforating arteries from the ICA, the anterior choroidal artery (AChA), the anterior cerebral artery (ACA), and the MCA to the basal ganglia, the anterior portion of the thalamus, the genu, and the anterior and posterior limbs of the internal capsule. It is also the exit site for the inferior striate veins. The APS is a convex cavity extending upward at the posterior end of the basal surface of the frontal lobe; it is bounded anteriorly by the lateral and medial olfactory striae, posteromedially by the optic tract, posterolaterally by the anteromedial surface of the uncus, and laterally by the limen insulae. Medially, the APS extends above the optic chiasm to the interhemispheric fissure. The APS and the carotid bifurcation can be identified intraoperatively by following the olfactory tract posteriorly. The APS can be considered the “floor” of the anterior half of the basal ganglia (Fig. 2-5C, right).
Basal Surface: Arterial Relationships
The internal carotid artery is divided into five parts: the cervical, petrous, cavernous, clinoid, and supraclinoid portions. The supraclinoid portion has been divided into three segments based on the origin of its major branches: the ophthalmic segment extends from the origin of the ophthalmic artery to the origin of the posterior communicating artery (PCom), the communicating segment extends from the origin of the PCom to the origin of the anterior choroidal artery (AChA), and the choroidal segment extends from the origin of the AChA to the bifurcation of the ICA (Fig. 2-5D). The ophthalmic artery arises under the optic nerve, usually from the medial third of the superior surface of the ICA, passes anteriorly and laterally to become superolateral to the carotid, and enters the optic canal and the orbit. The perforating arteries from this segment arise from the posterior, medial, or posteromedial aspect of the ICA and are distributed to the stalk of the pituitary gland, the optic chiasm, and less commonly the optic nerve, premamillary portion of the floor of the third ventricle, and the optic tract. The superior hypophysial arteries, which can range from 1 to 5 in number, pass medially to supply the pituitary stalk and the anterior lobe of the pituitary gland. The inferior hypophysial artery from the meningohypophysial trunk of the cavernous ICA supplies the posterior lobe. The infundibular arteries are another group of arteries that arise from the PCom and supply the same area as the superior hypophysial artery. The PCom arises from the posteromedial or the posterior or posterolateral aspect of the ICA and passes posteromedially to join the posterior cerebral artery (PCA) (Fig. 2-6A, left). In the embryo, the PCom continues as PCA, but in adults the PCA becomes part of the basilar system. If the PCom remains the major origin of the PCA, the configuration of the PCom is termed fetal (Fig. 2-6A, right). In 60% of individuals there are no perforating arteries arising from the communicating segment of the ICA; when present, the perforating arteries from the PCom range from 4 to 14 in number, arise predominantly from the proximal half of the artery, course superiorly, and terminate in the floor of the third ventricle. The largest branch from the PCom is the premamillary artery or “anterior thalamoperforating artery” (Fig. 2-6B).
The anterior choroidal artery arises from either the posterolateral or posterior aspect of the ICA. The AChA courses posteriorly below the optic tract toward the temporal horn by passing through the choroidal fissure (Fig. 2-6A, left). The AChA sends off branches to the optic tract, crus cerebri, lateral geniculate body, and uncus and supplies the optic radiation, globus pallidus, midbrain, thalamus, and the retrolenticular and posterior portions of the posterior limb of the internal capsule.
The choroidal segment of the ICA is the most frequent site of perforating arteries (range, one to nine) arising from the posterior aspect of the ICA. They terminate in the posterior half of the central region of the APS, optic tract, and uncus.16
The anterior perforating arteries are those arising from the ICA, MCA, AChA, and ACA, and they enter the brain through the APS (Fig. 2-6B).
The M1, or sphenoidal segment of the MCA, extends from the bifurcation of the ICA to the limen insulae. It courses first in the carotid cistern and then continues in the sphenoidal compartment. The proximal half of M1 is related posteriorly and inferiorly to the anteromedial surface of the uncus, anteriorly to the lesser wing of the sphenoid, and superiorly to the APS; the distal half is related inferiorly to the planum polare, anteriorly to the lesser wing of the sphenoid, and superiorly and posteriorly to the insular pole. M1 has two types of branches: the lateral lenticulostriate arteries, which arise mostly from the superior or posterosuperior aspect of M1 and penetrate the middle and posterior portions of the lateral half of APS, and the early branches, which course toward the temporal lobe to supply the temporal pole. The bifurcation of the MCA occurs before the limen insulae in 86% of individuals (see Figs. 2-4D and 2-6B and C).3
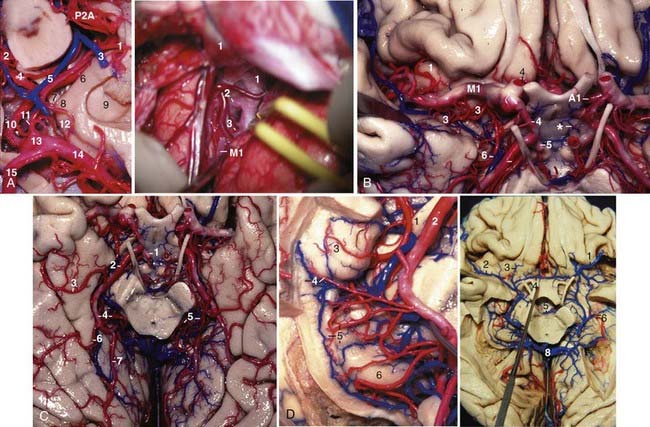
Embryologically, the posterior cerebral artery arises as a branch of the ICA, but up to birth its most frequent origin is the basilar artery.17 The PCA is divided into four segments: P1 extends from the basilar bifurcation to the site where the PCom joins the PCA. P2 extends from the PCom to the posterior aspect of the midbrain. P2 is further divided into P2A (anterior) and P2P (posterior) segments. P2A begins at the PCom and courses around the crus cerebri, inferior to the optic tract, AChA, and basal vein and medial to the posteromedial surface of the uncus, up to the posterior margin of the crus cerebri. P2P begins at the posterior margin of the crus cerebri; runs lateral to the tegmentum of the midbrain within the ambient cistern, parallel and inferior to the basal vein, inferolateral to the geniculate bodies and pulvinar, and medial to the parahippocampal gyrus; and enters the quadrigeminal cistern. P3 begins under the posterior part of the pulvinar in the lateral aspect of the quadrigeminal cistern and ends at the anterior limit of the anterior calcarine sulcus. P3 often divides into its major terminal branches, the calcarine and parieto-occipital arteries, before reaching the anterior limit of the anterior calcarine sulcus. The point where the PCAs from each side are closer to each other is called the collicular or quadrigeminal point. It marks the posterior limit of the midbrain on angiograms (see Fig. 2-14A). The P4 segment is the cortical branches of the PCA (Fig. 2-6C).
The main branches arising from the PCA are the posterior thalamoperforating, the direct perforating, the short and long circumflex, the thalamogeniculate, the medial and lateral posterior choroidal, the inferior temporal, the parieto-occipital, the calcarine, and the posterior pericallosal arteries. The posterior thalamoperforating arteries, which arise from P1 and enter the brain through the posterior perforated substance, interpeduncular fossa, and medial crus cerebri, supply the anterior and part of the posterior thalamus, hypothalamus, subthalamus, substantia nigra, red nucleus, oculomotor and trochlear nuclei, oculomotor nerve, mesencephalic reticular formation, pretectum, rostromedial floor of the third ventricle, and the posterior portion of the internal capsule. The direct perforating arteries to the crus cerebri arise mainly from the P2A segment and supply the crus cerebri. The short and long circumflex arteries to the brainstem arise mainly from P1 and less frequently from P2A; the short circumflex artery courses around the midbrain and terminates at the geniculate bodies, whereas the long circumflex artery courses around the midbrain and reaches the colliculi. The thalamogeniculate arteries arise equally from the P2A or P2P segments, perforate the inferior surface of the geniculate bodies, and supply the posterior half of the lateral thalamus, posterior limb of the internal capsule, and the optic tract (Fig. 2-6C). The medial posterior choroidal arteries (MPChAs) arise mainly from P2A and less frequently from the P2P and P1 segments, course around the midbrain medial to the main trunk of the PCA, turn around the pulvinar of the thalamus and proceed superiorly at the lateral side of the colliculi and pineal gland, enter the roof of the third ventricle through the velum interpositum, and finally course through the foramen of Monro to enter the choroid plexus in the lateral ventricle (see Figs. 2-3A, right, and 2-5D). The MPChA supplies the crus cerebri, tegmentum, geniculate bodies (mainly the medial one), colliculi, pulvinar, pineal gland, and medial thalamus. Angiographically on a lateral projection, the MPChA describes the shape of the number “3.” The inferior curve of the “3” is the point where it turns around the pulvinar, and the superior curve is the point where it contours the colliculi before entering the roof of the third ventricle (Fig. 2-14B). The lateral posterior choroidal arteries (LPChAs) arise mainly from P2P and less frequently from the P2A segment and pass laterally and enter the ventricular cavity directly through the choroidal fissure to supply the choroid plexus in the atrium and the temporal horn. It anastomoses with the AChA (see Figs. 2-4D and 2-6D, left). The inferior temporal arteries are distributed to the basal surface of the temporal and occipital lobes. They include the hippocampal artery and three groups of temporal arteries, namely, the anterior, middle, and posterior temporal arteries (Fig. 2-6C). The anterior temporal artery arises mainly from P2A, whereas the middle and posterior temporal arteries arise mainly from the P2P segment. The parieto-occipital and calcarine arteries are usually terminal branches of the PCA; they arise predominantly from P3 but may sometimes also arise from the P2P segment and course into the parieto-occipital and calcarine fissures, respectively. As the calcarine fissure reaches laterally and bulges into the medial wall of the atrium and the occipital horn, the calcarine artery also follows laterally into the depth of the calcarine fissure (see Fig. 2-4D). The splenial or posterior pericallosal artery supplies the splenium of the corpus callosum and arises from the parieto-occipital artery in 62% of individuals, but it can also arise from the calcarine artery, MPChA, posterior temporal artery, P2P, P3, and LPChA.
Basal Surface: Venous Relationships
The most important deep venous channel on the basal surface is the basal vein of Rosenthal. The basal vein originates below the APS and is divided into three segments (Fig. 2-6D, right): the first, or anterior or striate segment, originates from the junction of the anterior cerebral, inferior striate, olfactory, fronto-orbital, and deep middle cerebral veins under the APS and runs posteriorly under the optic tract, medial to the anterior portion of the crus cerebri. This point corresponds to the most medial (before its termination into the vein of Galen) and usually most inferior part of the basal vein and laterally indicates the location of the apex of the uncus. The second, or middle or peduncular segment, starts from the most medial point in the course of the basal vein, usually corresponding to the site where the peduncular vein joins the basal vein. It runs laterally between the upper part of the posteromedial surface of the uncus and the upper part of the crus cerebri and under the optic tract to reach the most lateral part of the crus cerebri, which corresponds to the most lateral point of the vein as it turns around the crus cerebri, generally where the inferior ventricular vein joins the basal vein; this is called the anterior peduncular segment by Huang and Wolf.18 It then turns medially, superiorly, and posteriorly to the plane of the lateral mesencephalic sulcus behind the crus cerebri to constitute the posterior peduncular segment. The main tributaries of the second segment are the peduncular or interpeduncular, inferior ventricular, inferior choroidal, hippocampal, and anterior hippocampal veins. The third, or posterior or posterior mesencephalic segment, runs medially, superiorly, and posteriorly from the lateral mesencephalic sulcus and under the pulvinar of the thalamus to penetrate the quadrigeminal cistern and generally drains into the vein of Galen. The main tributaries of the third segment are the lateral mesencephalic, posterior thalamic, posterior longitudinal hippocampal, medial temporal, and medial occipital veins. Sometimes, the precentral cerebellar, superior vermian, internal occipital, splenial, medial atrial, and direct lateral and lateral atrial subependymal veins may drain into the third segment of the basal vein. In the angiographic frontal view, the overall shape of both basal veins resembles the legs of a frog lying on its back with its toes directed anterolaterally. The foot corresponds to the striate segment and is related superiorly to the APS, laterally to the anterior segment of the uncus, medially to the optic tract, and inferiorly to the contents of the carotid cistern. The ankle corresponds posteriorly to the anterior aspect of the crus cerebri, laterally to the apex of the uncus, and superiorly to the optic tract; the leg corresponds to the anterior peduncular segment and is related superiorly to the optic tract, laterally to the upper portion of the posteromedial surface of the uncus, and medially to the upper portion of the crus cerebri. The knee corresponds to the most lateral aspect of the crus cerebri and to the posterior edge of the posterior segment of the uncus. It is related laterally to the inferior choroidal point, superiorly to the optic tract just before it reaches the lateral geniculate body, and inferiorly to the contents of the ambient cistern. The thigh, which includes the posterior peduncular and the posterior mesencephalic segments, is related medially to the tegmentum of the midbrain, laterally to the parahippocampal gyrus, superiorly to the medial aspect of the pulvinar of the thalamus, which is the roof of the wing of the ambient cistern, and inferiorly to the contents of the wing of the ambient cistern (see Fig. 2-6D).17 In the angiographic lateral view, the basal and the internal cerebral veins delimit the thalamus and hypothalamus (Fig. 2-7A; also see Fig. 2-13D).
Medial Surface: Neural Relationships
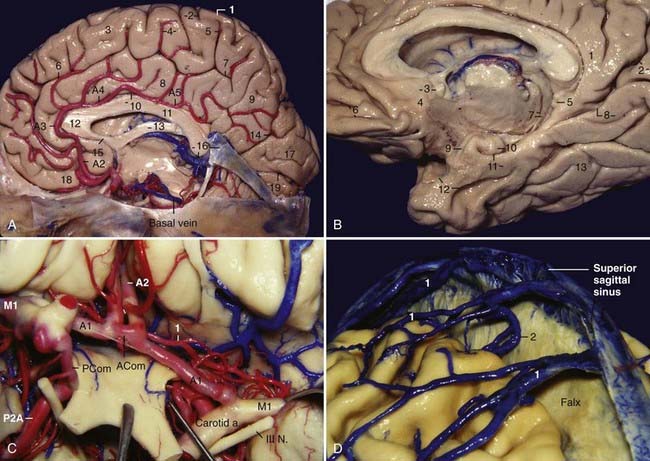
The medial surface of the cerebrum contains the sulci and gyri of the frontal, parietal, occipital, and temporal lobes. The general organization of the gyri of the frontal, parietal, and occipital lobes on this surface can be compared with that of a three-layer roll: the inner layer is represented by the corpus callosum, the intermediate layer by the cingulate gyrus, and the outer layer by the medial frontal gyrus, paracentral lobule, precuneus, cuneus, and lingual gyrus. The cingulate gyrus is separated inferiorly from the corpus callosum by the callosal sulcus and superiorly from the outer layer by the cingulate sulcus. Several secondary rami ascend from the cingulate sulcus in a radiating pattern and divide the outer layer into several sections. There are two secondary rami of particular importance: the paracentral ramus, which ascends from the cingulate sulcus at the level of the midpoint of the corpus callosum and separates the medial frontal gyrus anteriorly from the paracentral lobule posteriorly, and the marginal ramus, which ascends from the cingulate sulcus at the level of the splenium of the corpus callosum and separates the paracentral lobule anteriorly from the precuneus posteriorly. The marginal ramus intercepts the postcentral gyrus in almost 100% of individuals and is an important landmark to determine the location of the sensory or motor areas in the lateral convexity on midsagittal magnetic resonance images. The parieto-occipital sulcus separates the precuneus superiorly from the cuneus inferiorly, and the calcarine sulcus separates the cuneus superiorly from the lingual gyrus inferiorly. The paracentral ramus and the marginal ramus form the paracentral lobule, which is concerned with movements of the contralateral lower limb and perineal region and is involved in voluntary control of defecation and micturition. The paracentral lobule comprises the anterior portion of the postcentral and precentral gyri and the posterior portion of the superior frontal gyrus. The precuneus and the part of the paracentral lobule behind the central sulcus form the medial part of the parietal lobe; the precuneus corresponds to the superior parietal lobule on the lateral surface. The precuneus presents the subparietal sulcus, a vaguely H-shaped sulcus where the vertical arm of the H tends to align with the marginal ramus, and the parieto-occipital sulcus, which separates the precuneus above from the cingulate gyrus below (Fig. 2-7A). The parieto-occipital and calcarine sulci define the cuneus; the cuneus and medial part of the lingual gyrus are the medial portion of the occipital lobe. The calcarine sulcus starts at the occipital pole and is directed anteriorly; it has a slightly curved course with a characteristic upward convexity. The calcarine sulcus joins the parieto-occipital sulcus (only superficially) at an acute angle behind the isthmus of the cingulate gyrus and continues anteriorly to intercept the isthmus of the cingulate gyrus. The portion of the calcarine sulcus anterior to the junction is called anterior calcarine sulcus; it is crossed by a buried anterior cuneolingual gyrus and bulges into the medial wall of the atrium of the lateral ventricle as the calcar avis. It contains the visual cortex only on its lower lip. The part of the calcarine sulcus posterior to the union is called the posterior calcarine sulcus and includes the striate (visual) cortex on its upper and lower lips (Fig. 2-7A and B). Anteriorly, the cingulate and medial frontal gyri wrap around the genu and rostrum of the corpus callosum. At the inferior end of these two gyri, under the rostrum of the corpus callosum and in front of the lamina terminalis, is a narrow triangle of gray matter, the paraterminal gyrus, separated from the rest of the cortex by a shallow posterior paraolfactory sulcus. Slightly anterior to this sulcus, a short vertical sulcus may occur, the anterior paraolfactory sulcus; the cortex between the posterior and anterior paraolfactory sulci is the subcallosal area or paraolfactory gyrus. Frequently, two anteroposteriorly directed sulci, the superior and inferior rostral sulci, which are parallel to the floor of the anterior fossa, divide the inferior portion of the medial frontal gyrus into three parts. Posteriorly, the cingulate gyrus continues inferiorly with the parahippocampal gyrus through the isthmus of the cingulate gyrus. The mesial portion of the temporal lobe contains intraventricular and extraventricular elements. The intraventricular elements are the hippocampus, fimbria, amygdala, and choroidal fissure; the extraventricular elements are the parahippocampal gyrus, uncus, and dentate gyrus. The parahippocampal gyrus extends anteriorly to posteriorly, and at its anterior extremity, it deviates medially and bends posteriorly to constitute the uncus. Posteriorly, just bellow the splenium of the corpus callosum, the parahippocampal gyrus is often intersected by the anterior calcarine sulcus, which divides the posterior portion of the parahippocampal gyrus into the isthmus of the cingulate gyrus superiorly and the parahippocampal gyrus inferiorly; the parahippocampal gyrus continues posteriorly as the lingual gyrus. Superiorly, the parahippocampal gyrus is separated from the dentate gyrus by the hippocampal sulcus. Laterally, the parahippocampal gyrus is limited by the collateral sulcus posteriorly and the rhinal sulcus anteriorly. The rhinal sulcus marks the lateral limit of the entorhinal area of the parahippocampal gyrus; the parahippocampal gyrus is separated from the inferior surface of the posterior segment of the uncus by the uncal notch. Medially, the parahippocampal gyrus is related to the edge of the tentorium and to the contents of the ambient cistern. The various components of the parahippocampal gyrus are the subiculum, presubiculum, parasubiculum, and entorhinal area; the subiculum is the medial round edge of the parahippocampal gyrus. The name uncus means “hook.” It is formed by the anterior portion of the parahippocampal gyrus, which has deviated medially and folded posteriorly. Inferiorly, the uncus is separated from the parahippocampal gyrus by the uncal notch. Anteriorly, the uncus continues with the anterior portion of the parahippocampal gyrus without a sharp boundary; superiorly, the uncus is continuous with the globus pallidus. At the basal surface, the uncus is separated laterally from the temporal pole by the rhinal sulcus, and its medial part is normally herniated medially to the edge of the tentorium. When viewed from its basal surface, the uncus has the shape of an arrowhead with its apex pointing medially; it features an apex, an anterior segment, and a posterior segment (see Fig. 2-3D). The anterior segment of the uncus has one surface, the anteromedial surface, whereas the posterior segment has two surfaces, the posteromedial and inferior surfaces. Both segments converge superiorly at the junction between the amygdala and the globus pallidus. The uncus is composed of five small gyri and a small part of the entorhinal area, which occupies the anterior portion of the anteromedial surface. The anterior segment or anteromedial surface is part of the parahippocampal gyrus and contains the semilunar and ambient gyri. The semilunar gyrus occupies the superior portion of the anteromedial surface and is bordered inferiorly by the sulcus annularis, and the ambient gyrus is medial and inferior to the semilunar gyrus; the anteroinferior area of this surface is occupied by the entorhinal area, which continues anteriorly and inferiorly with the entorhinal area of the parahippocampal gyrus (Fig. 2-7B). The anteromedial surface is related to the proximal sylvian fissure and carotid cistern and is the posterolateral limit of the APS. The posterior segment is related to the hippocampus and has two surfaces: a posteromedial and inferior surface (see Figs. 2-3D, left, and 2-6D, left). The posterior segment is occupied by three small gyri; from anterior to posterior, they are the uncinate gyrus, the band of Giacomini, and the intralimbic gyrus. The superior and inferior portions of the posteromedial surface of the uncus are related, respectively, to the crural and ambient cisterns. Posterior and superior to the uncus is the inferior choroidal point, where the choroid plexus of the temporal horn begins. The inferior choroidal point corresponds to the site where the AChA enters and the inferior ventricular vein leaves the temporal horn through the choroidal fissure (see Fig. 2-6A, left, and 2-6D, right). The inferior surface is the superior lip of the uncal notch, and it is visible only from below when the parahippocampal gyrus is removed. The dentate gyrus bears this name because of its characteristic tooth-like elevations; the margo denticulatus is prominent mainly in its anterior and middle portions. The dentate gyrus continues anteriorly with the band of Giacomini, also called the tail of the dentate gyrus, and continues posteriorly with the fasciolar gyrus, a smooth grayish band that is located posterior to the splenium of the corpus callosum; the fasciolar gyrus continues above the corpus callosum as the indusium griseum and finally ends as the paraterminal gyrus. The fimbriodentate and hippocampal sulci separate the dentate gyrus, respectively, from the fimbria superiorly and the parahippocampal gyrus inferiorly (Fig. 2-7B).
The extraventricular and intraventricular structures of the mesial temporal lobe are intimately related. The anterior segment of the uncus is related to M1, the carotid artery, and the amygdala. The apex of the uncus passes above the oculomotor nerve and is related to the uncal recess and the amygdala laterally (see Figs. 2-5D and 2-6A); the posterior segment is related to the head of the hippocampus and the amygdala laterally, to P2A inferomedially, and to the AChA superomedially.
Medial Surface: Arterial Relationships
The anterior cerebral artery has five segments. The A1 segment extends from the bifurcation of the ICA to the anterior communicating artery (ACom). The A2 segment extends from the ACom to the junction between the rostrum and the genu of the corpus callosum. The A3 segment extends from the genu of the corpus callosum to the point where the artery turns sharply and posteriorly above the genu of the corpus callosum. The A2 and A3 segments together are also called the ascending segment. The A4 and A5 segments extend above the corpus callosum, from the genu to the splenium. These two segments together are also called the horizontal segment, and a point bisected in the lateral view close behind the coronal suture separates them. The segment of the ACA distal to the ACom (A2 to A5) has also been called the pericallosal artery (see Fig. 2-7A). The junction of the ACom with the A1 segment occurs above the chiasm in 70% of individuals and above the nerve in 30%. The shorter A1 segments are usually stretched tightly over the chiasm; the longer ones pass anteriorly over the optic nerve and can be elongated and tortuous and reach either the tuberculum sellae or the planum sphenoidale (Fig. 2-7A). The medial lenticulostriate perforators, ranging from 1 to 11 branches (average of 6.4), arise from the superior, posterior, or posterosuperior aspect of the proximal half of the A1 segment and pursue a direct posterior and superior course to enter the medial half of the APS.19 Embryologically, the ACom develops from a multichanneled vascular network that coalesces to a variable degree by the time of birth.19 Only in 20% of individuals does the ACom communicate with two A1 segments of equal size. The ACom complex probably exists as a single channel in about 75% of individuals.19 The perforators from the ACom, ranging from 0 to 4 (average of 1.6), usually arise from its posteroinferior aspect and supply the infundibulum, the APS, the optic chiasm, the subcallosal area, and the preoptic areas of the hypothalamus. The recurrent artery of Heubner of the ACA arises in 78% of individuals from the proximal A2 segment, and it doubles back on its parent vessel, courses anterior to the A1 segment in 60% of individuals, and can be seen on elevating the frontal lobe before visualization of the A1 segment; it is the largest and longest branch directed to the APS. After its origin, it passes above the carotid bifurcation and accompanies the M1 segment into the medial part of the sylvian fissure before entering the anterior and middle portions of the full mediolateral extent of the APS (see Fig. 2-7C). The A2 segment is also the source of the central or the basal perforating arteries, which pass posteriorly and enter the optic chiasm, the lamina terminalis, and the anterior forebrain, below the corpus callosum. The two first cortical branches of the ACA supplying the medial surface, the orbitofrontal and the frontopolar arteries, usually arise from the A2 segment. The segments A3 to A5 give rise to other cortical branches and supply the medial surface of the hemisphere. All the cortical branches arise more frequently from the pericallosal than from the callosomarginal artery.
Medial Surface: Venous Relationships
The medial frontal veins drain the medial surface of the frontal lobe. They can empty either superiorly into the superior sagittal sinus or inferiorly into the inferior sagittal sinus or into the veins that pass around the corpus callosum and drain into the anterior end of the basal vein. The medial parietal veins drain the medial surface of the parietal lobe. They can either empty superiorly into the superior sagittal sinus or course around the splenium of the corpus callosum and drain inferiorly into the vein of Galen or its tributaries. On both lobes, the veins commonly curve over the superior margin of the hemisphere onto the upper part of the lateral surface, where they join the terminal end of the veins from the lateral surface before emptying into the superior sagittal sinus (see Fig. 2-7D). The posterior pericallosal veins, one on each side, arise from tributaries that drain the posterior part of the cingulate gyrus and the precuneus and course side by side around the splenium of the corpus callosum to terminate in either the vein of Galen or the internal cerebral vein. The anterior and posterior calcarine veins drain the occipital lobe. The anterior calcarine or internal occipital vein arises from tributaries that drain the anterior portion of the cuneus and lingual gyrus and passes forward to join the posterior pericallosal vein near the splenium before terminating in either the internal cerebral vein or the vein of Galen. The posterior calcarine vein arises from tributaries that drain the area bordering the posterior part of the calcarine fissure and then curves sharply upward on the cuneus to reach the superior sagittal sinus.
The deep venous system of the mesial temporal region drains into the basal vein of Rosenthal.
Posterior Fossa
Brainstem
The brainstem is divided into three parts: midbrain, pons, and medulla.
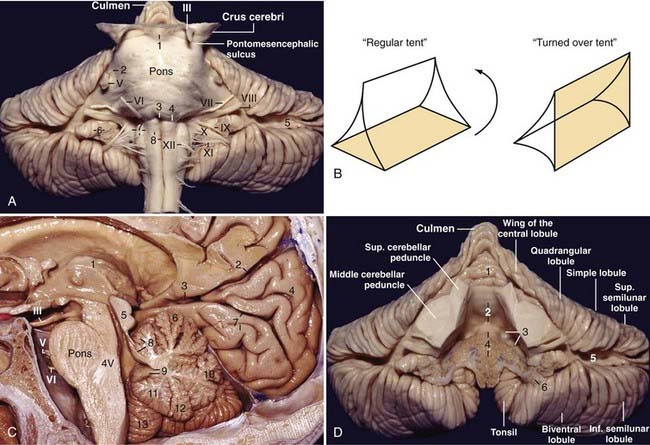
The midbrain is divided by a midline sagittal plane into two cerebral peduncles. Each peduncle is further divided into three parts: an anterior part, the crus cerebri or basis pedunculi; an intermediate part, the tegmentum; and a posterior part located behind the aqueduct, the tectum. The substantia nigra and lateral mesencephalic sulcus separate the crus cerebri from the tegmentum. The oculomotor nerves emerge from the medial side of the crura cerebri in the interpeduncular fossa (see Fig. 2-5C). The pontomesencephalic sulcus, which separates the midbrain from the pons, originates in the depth of the interpeduncular fossa and runs around the inferior margin of the crus cerebri to join the lateral mesencephalic sulcus behind the crus cerebri. The posterior aspect of the midbrain contains the superior and inferior colliculi (quadrigeminal plate). The trochlear nerve exits the brainstem below the inferior colliculus.
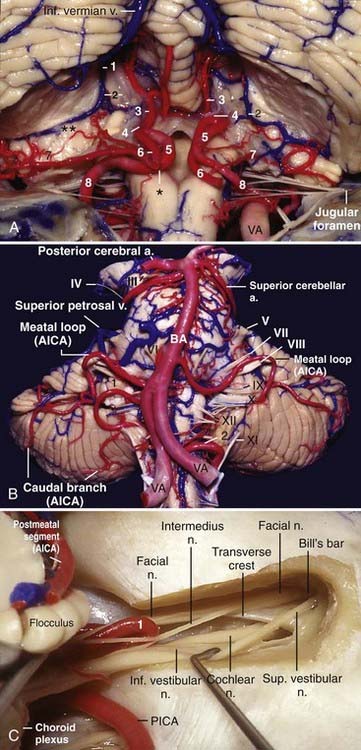
The supraolivary fossette is a triangular depression located behind and above the olive, anteromedial to the flocculus, and corresponds to the junction of the pons, the medulla, and the middle and inferior cerebellar peduncles. It is limited superiorly by the inferior aspect of the pons and the middle cerebellar peduncle and posteriorly by the inferior cerebellar peduncle. The fossette resembles a right-angled triangle with its right angle located between the superior pole of the olive and the inferior aspect of the pons, the superior leg corresponds to the inferior border of the pons and the middle cerebellar peduncle, the vertical leg corresponds to the posterior border of the olive, and the hypotenuse corresponds to the inferior cerebellar peduncle. Cranial nerves VI, VII, and VIII exit from the brainstem at the superior leg, and nerves IX, X, and XI exit from the brainstem at the hypotenuse (Fig. 2-8A).
Cerebellum
The fourth ventricle is often described as a tent-shaped midline structure surrounded mainly by the vermian components of the cerebellum. A regular tent has a roof that is divided into two halves, a floor, and two lateral walls; the fourth ventricle resembles a turned-over tent with its base facing forward and two open lateral walls. The floor is represented by the pons and medulla; the superior cerebellar peduncles, superior medullary velum, and the adjacent lingula constitute the superior part of the roof; the inferior part of the roof is composed of the inferior medullary velum, tela choroidea, choroid plexus, uvula, and the nodule; and the two open lateral walls are represented by lateral recesses through which the fourth ventricle communicates with the cerebellopontine angle (Fig. 2-8B and C).
Petrosal Surface of the Cerebellum and Fourth Ventricle
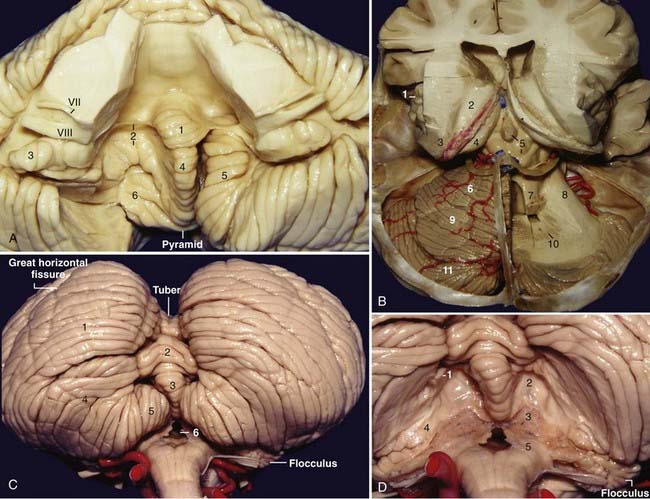
Each half of the petrosal surface is intersected by the great horizontal fissure, or petrosal fissure, that circumscribes the cerebellum.20 At the level of the flocculus, the petrosal fissure bifurcates into a larger suprafloccular portion and a smaller infrafloccular portion of the posterolateral fissure, which separates the flocculonodule lobule from the rest of the cerebellum and communicates with the cerebellomedullary fissure at the cerebellopontine angle. The folia that constitute the upper half of the petrosal surface are the folia of the tentorial surface that have folded over the middle cerebellar peduncle and over the core of the cerebellum. These folia are the wing of the central lobule and the quadrangular, simple, and superior semilunar lobules. The folia that constitute the lower half of the petrosal surface are the folia derived from the suboccipital surface that have folded over the inferior cerebellar peduncle and over the core of the cerebellum and correspond to the inferior semilunar and biventral lobules (Fig. 2-8A and D).
The lateral recess is the lateral extension of the fourth ventricle, and it connects the fourth ventricle to the cerebellopontine angle. It is directed in a medial-to-lateral, slightly superior-to-inferior, and posterior-to-anterior direction and forms an angle of about 45 degrees with the sagittal plane. The lateral recess has anterior, superior, and posterior walls and a floor. The anterior and superior walls are formed by the inferior cerebellar peduncle as it runs upward and then turns backward toward the cerebellum. The floor of the lateral recess consists of the tela choroidea anteriorly, the choroid plexus in the middle, and the inferior medullary velum posteriorly; at the foramen of Luschka the inferior medullary velum becomes thicker and is called the peduncle of the flocculus and forms the posterior wall of the foramen of Luschka. The superolateral recess is the space in the fourth ventricle limited medially by the nodule and inferiorly by the superior pole of the tonsil and covered by the inferior medullary velum. Above the superolateral recess, the superior cerebellar peduncle has a prominence, the dentate tubercle, where the dentate nucleus comes to the surface (Fig. 2-8D).
The morphology of the choroid plexus of the fourth ventricle resembles the letter “T” with two vertical bars. The horizontal part of the choroid plexus, which starts from the fourth ventricle and protrudes into the cerebellopontine angle, resembles the horns of a bull. The vertical part and the proximal half of the horizontal part of the choroid plexus of the fourth ventricle are usually supplied by the PICA; the lateral half of the horizontal part and the choroid plexus located at the cerebellopontine angle are generally supplied by the AICA.21
The tonsils are two reniform structures that are the hemispheric components of the uvula and are attached to the cerebellum through the peduncles of the tonsil, which are located at the superolateral aspect of each tonsil. The superior, medial, anterior, posterior, and most of the lateral surfaces of the tonsils are free. The spaces around the tonsils are the supratonsillar space between its superior pole and the inferior medullary velum, the vallecula between the medial surfaces of the two tonsils, the cerebellomedullary fissure between the anterior surface of the tonsil and the medulla, and the retrotonsillar space between the posterior surface of the tonsil and the adjacent vermis. The furrowed band of Reil connects the uvula to the tonsil, and the copula pyramidis connects the pyramid to the biventral lobule (Fig. 2-9A). The copular point is the angiographic landmark at which the retrotonsillar veins unite to form the inferior vermian vein; the copular point denotes the location of the copula pyramidis. The copular point can also be defined as the lowest part of the inferior vermian artery, the vermian branch of the PICA surrounding the copula pyramidis.
Tentorial Surface of the Cerebellum and Fourth Ventricle
The tentorial surface faces the tentorium and consists of two cerebellar incisurae, three margins, and two angles. The cerebellar incisurae are the anterior and posterior cerebellar incisurae; the brainstem fits into the anterior cerebellar incisura and the falx cerebelli fits into the posterior cerebellar incisura. The margins are the anterosuperior margin, or the posterior wall of the cerebellomesencephalic fissure that extends from the top of the culmen downward, forward, and laterally to reach a point above and behind the middle cerebellar peduncle; the anterolateral margin, which separates the tentorial from the petrosal surface; and the posterolateral margin, which separates the tentorial from the suboccipital surface. The junction between the anterosuperior and anterolateral margins forms the anterior angle, and the junction between the anterolateral and posterolateral margins forms the lateral angle (see Figs. 2-11C and 2-14D). Angiographically on the lateral projection, the lateral angle is located just below the knee between the transverse and sigmoid sinuses, and the outer portion of the anterolateral margin runs somewhat below the superior petrosal sinus (Fig. 2-15A). Anteriorly to posteriorly, the vermis and the hemispheric counterpart of the tentorial surface are the lingula (without the hemispheric counterpart), the central lobule (wing of the central lobule), the culmen (quadrangular lobule), the declive (simple lobule), and the folium (part of the superior semilunar lobule). The primary fissure is located between the quadrangular and simple lobules; the most prominent fissure, the postclival fissure, is located between the simple and superior semilunar lobules. The tentorial surface contains the cerebellomesencephalic or precentral cerebellar fissure, which is situated between the cerebellum and the midbrain. Posteriorly, it is limited by the culmen and quadrangular lobule above and the central lobule and its wing below. Anteriorly, it is limited from midline to laterally by the lingula and the superior and middle cerebellar peduncles. The interpeduncular or interbrachial sulcus, which separates the superior from the middle cerebellar peduncles, ascends from the bottom of the cerebellomesencephalic fissure toward the lateral aspect of the pons, where it is joined by the pontomesencephalic sulcus and proceeds superiorly as the lateral mesencephalic sulcus to the medial geniculate body; the lateral mesencephalic sulcus separates the crus cerebri from the tegmentum (see Fig. 2-9B).
Suboccipital Surface of the Cerebellum and Fourth Ventricle
The suboccipital surface contains the posterior cerebellar incisura and the vermohemispheric or paravermian fissure, which separates the inferior vermis from the cerebellar hemisphere. The components of the inferior vermis and its hemispheric counterparts are the folium (superior semilunar lobule), tuber (inferior semilunar lobule), pyramid (biventral lobule), uvula (tonsil), and nodule (flocculus). In the anatomic position, the most inferior part of the inferior vermis is the pyramid. The most prominent fissure on the suboccipital surface is the great horizontal fissure, which is a circumferential fissure that begins in the posterior cerebellar notch between the folium and the tuber and runs forward and slightly downward on the suboccipital surface, between the superior and inferior semilunar lobules, and then onto the petrosal surface as the petrosal fissure. The secondary fissure is located between the tonsils and the biventral lobule (Fig. 2-9C).
After removal of the tonsils, the inferior portion of the roof of the fourth ventricle comes into view (Fig. 2-9D). After removal of the inferior portion of the roof of the fourth ventricle, the floor of the fourth ventricle is exposed.
The floor of the fourth ventricle has a rhomboid shape and consists of a strip between the lower margin of the cerebellar peduncles and the site of attachment of the tela choroidea; called the junctional part, this strip is formed by the medullary striae, which extend into the lateral recesses. The junctional part divides the floor of the fourth ventricle into two unequal triangles: the superior and larger one, with its apex directed toward the aqueduct, is the pontine part, and the inferior and smaller one, with its apex directed toward the obex, is the medullary part of the floor. These three parts of the floor are also divided longitudinally into two symmetrical halves by the median sulcus. The sulcus limitans, another longitudinal sulcus, divides each half of the floor into a raised median strip called the median eminence and a lateral strip called the area vestibularis. The motor nuclei of the cranial nerves are located medial to the sulcus limitans, and the sensory nuclei are situated lateral to it. The pontine part is characterized by two rounded prominences, the facial colliculi, located on the median eminence, one on each side of the median sulcus. The facial colliculi are limited laterally by the superior fovea, a dimple formed by the sulcus limitans. The medullary part has the configuration of a feather, or pen nib, and is called the calamus scriptorius, with three triangular areas overlying the hypoglossal and vagus nuclei (hypoglossal and vagal trigones) and the area postrema; just lateral to the hypoglossal trigone, the sulcus limitans has another dimple called the inferior fovea. At the junctional part the sulcus limitans is discontinuous (Fig. 2-10A).
Veins of the Posterior Fossa
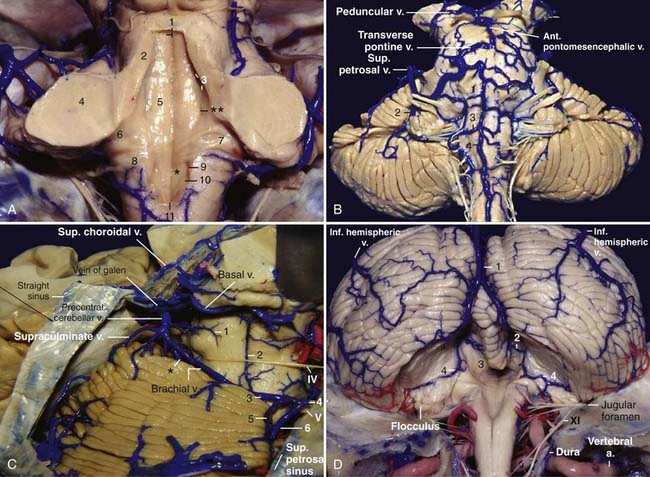
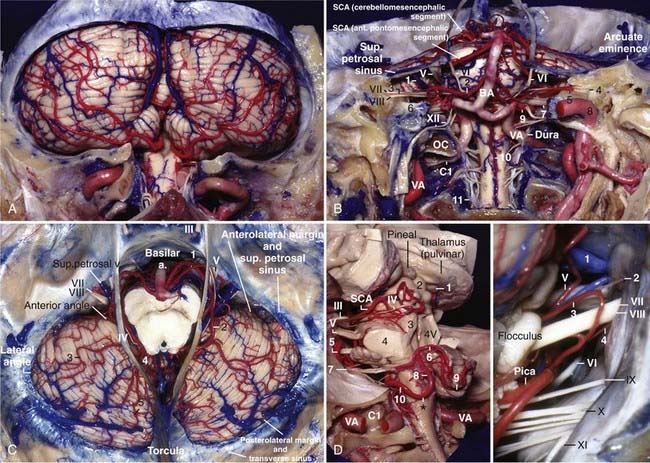
The posterior fossa venous system is divided into three groups: the anterior or petrosal group, which drains into the superior and inferior petrosal sinuses; the superior or galenic group, which drains into the vein of Galen; and the posterior or tentorial group, which drains into the sinuses near the torcula.22 There is a tendency for the veins to drain into the nearest draining system.
The veins running on the petrosal surface of the cerebellum and the anterior surface of the brainstem tend to drain into the petrosal sinuses via the superior petrosal vein, except for the veins running on the surface of the midbrain, which drain into the galenic system. The superior petrosal vein is usually formed by the junction of the transverse pontine and pontotrigeminal (brachial) veins and the vein of the cerebellopontine fissure (great horizontal fissure) (Fig. 2-10B).
The tentorial surface and the posterior aspect of the brainstem are served by three draining systems: the midline portion of the cerebellomesencephalic fissure, the veins near the central lobule and culmen (superior vermian veins), and the veins draining the intermediate portion of the wing of the central lobule and the quadrangular lobule (superior hemispheric veins, anterior group), which tend to drain into the vein of Galen. The veins draining the lateral portion of the wing of the central, quadrangular, and simple lobules and the tentorial part of the superior semilunar lobule (superior hemispheric veins, lateral group) tend to drain into the superior petrosal sinus. The veins draining the declive, folium (declival vein), and the intermediate portion of the simple and superior semilunar lobules (superior hemispheric veins, posterior group) tend to drain into the torcula or transverse or tentorial sinus in the tentorium cerebelli (Fig. 2-10C).
The posterior inferior hemispheric veins drain the suboccipital surface of the cerebellar hemispheres. Drainage of the inferior vermis is via the inferior vermian veins, which are formed by the junction of the superior and inferior retrotonsillar veins running in the retrotonsillar space (Fig. 2-10D).
The inferior portion of the roof of the fourth ventricle and the lateral recess are drained by the vein of the lateral recess of the fourth ventricle, also called the vein of the cerebellomedullary fissure. It courses laterally under the lateral recess toward the cerebellopontine angle, passes above or below the flocculus, joins the vein of the middle cerebellar peduncle or the vein of the cerebellopontine fissure, and finally empties into the superior petrosal sinus via the superior petrosal vein. The vein of the lateral recess of the fourth ventricle can also anastomose with the retrotonsillar veins at the retrotonsillar space to establish communication between the petrosal and the tentorial groups of venous drainage (Fig. 2-10D).
The brachial veins running in the cerebellomesencephalic fissure can also establish communication between the petrosal and galenic groups via the pontotrigeminal and precentral cerebellar veins (Fig. 2-10C).
The veins of the posterior fossa can be differentiated into the petrosal group, the superior or galenic group, and the posterior or tentorial group (see Figs. 2-14D and 2-15A to C):
Arteries of the Posterior Fossa
The vertebral artery (VA) arises from the subclavian artery, enters the transverse foramen of C6, and then ascends through the transverse foramina of the upper cervical vertebrae up to C2. After exiting from the transverse foramen of C2, the VA deviates laterally and enters the laterally placed transverse foramen of C1. The VA then turns behind the lateral mass and above the posterior arch of C1, courses medially and superiorly, and pierces the dura at the foramen magnum. At this level the VA usually gives off the posterior spinal and posterior meningeal arteries. The intradural segment of the VA is divided into lateral medullary and anterior medullary segments before joining its contralateral mate to form the basilar artery (Figs. 2-10D and 2-11B; also see Fig. 2-12B).
FIGURE 2-14 A, Frontal view of a vertebrobasilar angiogram. 1, Collicular or quadrigeminal point; 2, P3 segment; 3, beginning of the P2A segment; 4, collateral sulcus; 5, beginning of the cerebellomesencephalic segment of the superior cerebellar artery (SCA); 6, P2A segment; 7, lateral pontomesencephalic segment of the SCA; 8, anterior pontomesencephalic segment of the SCA; 9, supratonsillar segment of the posterior inferior cerebellar artery (PICA, cranial loop); 10, meatal loop of the anterior inferior cerebellar artery (AICA); 11, posterior medullary segment of the PICA; 12, caudal loop of the PICA; 13, lateral medullary segment of the PICA; 14, anterior medullary segment of the PICA; 15, extradural vertebral artery behind the lateral mass of C1; 16, vertebral artery in the foramen transversarium of C1; 17, vertebral artery in the foramen transversarium of C2. The blue arrowhead indicates the origin of the AICA from the basilar artery, the red arrowhead indicates the origin of the PICA from the vertebral artery, and the green arrows indicate the probable transition between the extradural and intradural segments of the vertebral artery. Note the constriction in the vertebral artery. B, Lateral view of the late arterial phase of a vertebrobasilar angiogram. 1, “Cranial loop” of the PICA; 2, vermian division of the PICA (pyramidal loop); 3, hemispheric branch of the PICA; 4, posterior medullary segment of the PICA; 5, “caudal loop” of the PICA. The red arrowheads indicate the lateral posterior choroidal arteries in the lateral ventricle (indicate the location of the posterior wall of the pulvinar of the thalamus), the green arrow indicates the posterior pericallosal artery, and the blue arrowheads indicate the medial posterior choroidal artery (MPChA). The most posterior point of the trajectory of the MPChA indicates the posterior limit of the quadrigeminal plate and consequently the posterior limit of the brainstem in the lateral view (yellow dashed line). For the anatomic location of the lateral posterior choroidal artery and MPChA, refer to Figures 2-4B and 2-6D. Note that the foramen of Monro is located above the tip of the basilar artery (in the same coronal plane). C, Lateral view of the arterial phase of a vertebrobasilar angiogram. 1. “Meatal loop” of the AICA. The blue arrowheads indicate small branches to the lateral recess of the fourth ventricle through the foramen of Luschka, and the red arrowheads indicate the main trunk of the AICA in the great horizontal fissure, which is supplying an arteriovenous malformation located in the superior semilunar lobule. D, Lateral view of the cerebellum. Compare this illustration with Figures 2-8B and C and 2-12A. 1, Flocculus; 2, petrosal fissure or great horizontal fissure; 3, superior semilunar lobule; LA, lateral angle. The arrows indicate the anterolateral margin.
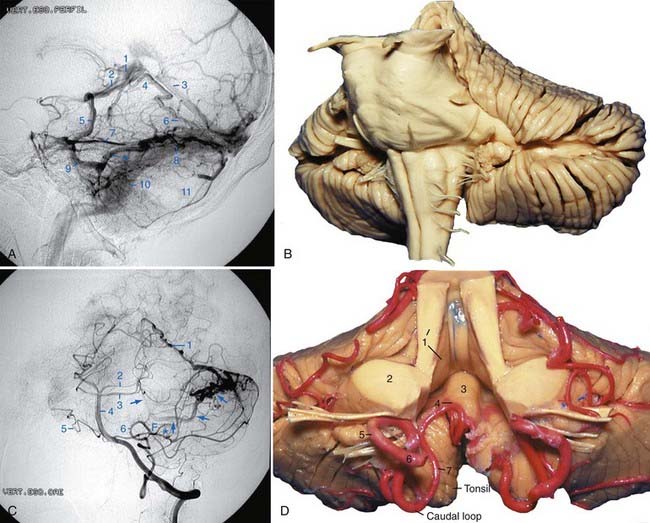
FIGURE 2-15 A, Lateral view of the venous phase of a vertebrobasilar angiogram. Compare with Figure 2-8D. 1, Vein of Galen; 2, cerebellomesencephalic fissure; 3, straight sinus; 4, culmen; 5, vein running on the middle cerebral peduncle that continues with a vein in the petrosal fissure (vein of the great horizontal fissure)9; 6, vein running on the tentorial surface of the cerebellum (superior hemispheric vein) toward the vein of Galen; 7, superior petrosal sinus; 8, transverse sinus; 9, vein running in the petrosal fissure or great horizontal fissure (vein of the great horizontal fissure); 10, sigmoid sinus; 11, suboccipital surface of the cerebellum; *, a superior hemispheric vein draining an arteriovenous malformation that runs on the tentorial surface, descends toward the petrosal surface, and joins the vein of the great horizontal fissure. B, Left anterior oblique view of the cerebellum and brainstem. Compare this illustration with Figure 2-12C. C, Left anterior oblique view of a vertebrobasilar angiogram. 1, Vein running on the tentorial surface toward the vein of Galen (superior hemispheric vein); 2, posterior cerebral artery; 3, superior cerebellar artery; 4, basilar artery; 5, meatal loop of the anterior inferior cerebellar artery (AICA); 6, posterior inferior cerebellar artery (PICA). The arrows indicate the vein shown in Figure 2-12A, which originates from the tentorial surface (superior hemispheric vein) and descends toward the petrosal surface to join the vein in the great horizontal fissure. F, approximate location of the flocculus; it was estimated by the location meatal loop of the AICA.* D, Anterior view of the PICA and the roof of the fourth ventricle. Compare this illustration with Figure 2-8A. 1, Superior cerebellar peduncle; 2, middle cerebellar peduncle; 3, nodule (covered by the inferior medullary velum); 4, supratonsillar segment of the PICA (cranial loop); 5, lateral medullary segment of the PICA; 6, anterior medullary segment of the PICA; 7, posterior medullary segment of the PICA.
The lateral medullary segment of the VA extends from its entrance into the posterior fossa to the preolivary sulcus. From its entrance the VA courses anteriorly, medially, and superiorly through the lower cranial nerve rootlets, lateral to the medulla, to reach the preolivary sulcus. The anterior medullary segment begins at the preolivary sulcus, courses in front of or between the hypoglossal rootlets, and crosses the pyramid to join with the other VA at or near the pontomedullary sulcus to form the basilar artery. The main branches of the VA are the posterior spinal artery, anterior spinal artery, PICA, and anterior and posterior meningeal arteries. The VA also sends off branches to supply the lateral and anterior parts of the medulla along its way around the medulla (see Figs. 2-11B, 2-12B, and 2-14A).
The posterior inferior cerebellar artery arises from the VA and supplies the medulla, the inferior vermis, the inferior portion of the fourth ventricle, the tonsils, and the inferior aspect of the cerebellum. The “regular” PICA has the most complex and variable course of the cerebellar arteries and is divided into five segments.23 The anterior medullary segment lays in front of the medulla and extends from the origin to the level of the inferior olive. The lateral medullary segment courses beside the medulla and extends from the inferior olive to the origin of the glossopharyngeal, vagus, and accessory nerves. The tonsillomedullary or posterior medullary segment begins at the level of the nerves and loops below the inferior pole of the cerebellar tonsil and upward along the medial surface of the tonsil toward the inferior medullary velum (caudal loop). The telovelotonsillar or supratonsillar segment courses in the cleft between the tela choroidea and the inferior medullary velum rostrally and the superior pole of the cerebellar tonsil caudally. It begins below the fastigium, where the PICA turns posteriorly over the medial side of the superior pole of the tonsil. This segment forms the “cranial loop.” It sometimes passes posteriorly before reaching the superior pole of the tonsil, thus giving the cranial loop a variable relationship to the fastigium. The junction of the posterior medullary and supratonsillar segments is called the choroidal point. The fifth segment is the cortical segment; after a short distance distal to the apex of the cranial loop, the PICA continues posteriorly downward in the retrotonsillar fissure, where it usually bifurcates into the tonsillohemispheric branch, which supplies the under aspect of the cerebellar hemisphere, and the inferior vermian branch, which lies on the lower aspect of the inferior vermis and forms a convex loop around the copula pyramidis (pyramidal loop). The most anterior point of the pyramidal loop is called the copular point. The terminal portion of the vermian branch curves around the tuber in the posterior cerebellar notch (see Figs. 2-11A and B; 2-11, left; 2-12A; 2-14A and B; 2-15C and D; 2-16A and B).
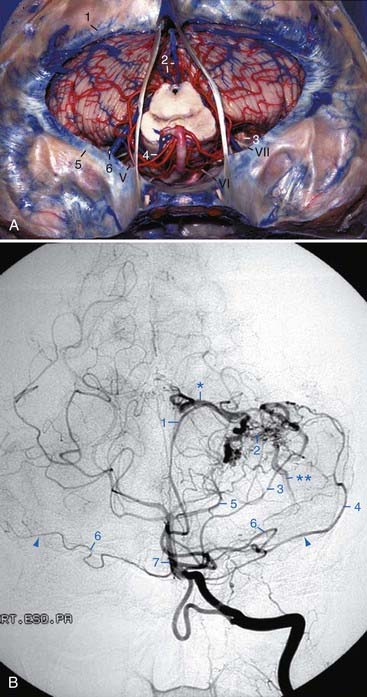
The anterior inferior cerebellar artery and the PICA are defined according to their origin rather than by the portions of cerebellum that they supply. The AICA arises more frequently from the lower third and less frequently from the middle third of the basilar artery. It courses posteriorly, laterally, and usually downward on the belly of the pons, in contact with either the superior or inferior aspect of the abducens nerve. In this course it supplies the lateral aspect of the lower two thirds of the pons and the upper medulla. Either immediately before or after crossing the roots of the facial, intermedius, and acoustic nerves within the cerebellopontine angle, the AICA bifurcates into its two major branches, the rostrolateral and the caudomedial arteries. The main or rostrolateral trunk has been divided into three segments according to their relationship to cranial nerves VII and VIII24: the premeatal, meatal, and postmeatal segments. The premeatal segment begins at the basilar artery and courses around the brainstem to reach cranial nerves VII and VIII and the region of the meatus, usually anteroinferior to the nerves. Seventy-seven percent of internal auditory arteries and 49% of recurrent perforating arteries to the brainstem arise from this segment. The meatal segment is located in the vicinity of the internal auditory meatus, where the nerve-related vessels turn toward the brainstem; this segment often forms a laterally convex loop, the meatal loop, directed toward or through the meatus. It usually stays medial to the meatus, but it sometimes protrudes into the canal. The postmeatal segment begins distal to the nerves and courses medially to supply the brainstem and the cerebellum. The subarcuate artery generally arises from this segment (see Figs. 2-11B and D, right; 2-12B; 2-14A and C; 2-15C; 2-16A and B).
The caudomedial artery originates on the lateral aspect of the pons in the vicinity of the sixth nerve and courses posterosuperiorly toward the pontomedullary sulcus; it has a caudal loop on the lateral aspect of the pons and medulla. This lateral loop can course on the anteroinferolateral aspect of the flocculus or on the petrosal aspect of the biventral lobule. Multiple small arteries to the choroid plexus of the lateral recess often arise from the inner aspect of this lateral loop. Distal to the loop, the biventral segment turns posteroinferiorly on the lateral edge of the inferior surface of the biventral lobule or within the cerebellomedullary fissure to reach the posterior surface of the cerebellum, where it anastomoses with branches of the PICA (Fig. 2-12B).
The superior cerebellar artery is the most rostral of the infratentorial vessels, and it arises near the apex of the basilar artery and encircles the pons and the lower midbrain. It supplies the tentorial surface of the cerebellum, the upper brainstem, the deep cerebellar nuclei, and the inferior colliculi. The SCA is divided into four segments. The anterior pontomesencephalic segment courses laterally under the oculomotor nerve on the anterior aspect of the upper pons, often in an arcuate convex curve inferiorly; the configuration of the anterior pontomesencephalic segment is related to the height of the basilar bifurcation. With a low basilar bifurcation (anterior to the pons), this segment tends to pass upward, whereas with a high basilar bifurcation (anterior to the midbrain), this segment pursues an anterior and inferior course. The lateral pontomesencephalic segment begins at the anterolateral margin of the brainstem and follows caudally onto the lateral side of the upper pons in the infratentorial portion of the ambient cistern, where it terminates at the anterior margin of the cerebellomesencephalic fissure; it is related medially to the brainstem, laterally to the wing of the central lobule, and inferiorly to the middle cerebellar peduncle. The anterior part of this segment is often visible above the free edge of the tentorium, whereas its caudal loop projects toward and often reaches the root entry zone of the trigeminal nerve. Bifurcation of the SCA into rostral and caudal trunks often occurs in this segment; the rostral trunk supplies the vermis and a variable portion of the adjacent tentorial surface, and the caudal trunk supplies the surface lateral to the area supplied by the rostral trunk. The cerebellomesencephalic segment courses in the cerebellomesencephalic fissure through a series of hairpin-like curves and then passes upward to reach the anterosuperior margin of the cerebellum. Inside the cerebellomesencephalic fissure, the rostral and caudal trunks send off small precentral branches. These precentral branches arising from the rostral trunk supply the inferior colliculi (the superior colliculi are supplied by the PCA) and the superior medullary velum, and those arising from the caudal trunk supply the deep cerebellar nuclei. Finally, the cortical segment is represented by the hemispheric and the vermian branches, which supply the tentorial surface of the cerebellum. Among these cortical branches, the marginal or lateral branch deserves special attention: it is present in 62% of individuals, it is the first large cortical branch of the SCA, and it arises from the lateral pontomesencephalic segment and courses anteriorly and laterally to reach the anterolateral margin of the cerebellum. It is an important arteriographic landmark for locating the anterolateral margin and the anterior angle of the cerebellum (see Figs. 2-11A and C, 2-14A, 2-15C, 2-16A and B).25
After their origin from the brainstem, the facial and intermedius nerves pass anterolaterally with the vestibulocochlear nerve; in this location, the facial nerve lies in an anterosuperior groove on the vestibulocochlear nerve, with the intermedius nerve between them. At the lateral end of the internal acoustic meatus, the vertical Bill bar and the transverse crest divide the fundus of the meatus into four quadrants: the facial nerve is located in the anterosuperior quadrant, the cochlear nerve in the anteroinferior quadrant, the superior vestibular nerve in the posterosuperior quadrant, and the inferior vestibular nerve in the posteroinferior quadrant (see Fig. 2-12C).
Fujii K, Lenkey C, Rhoton ALJr. Microsurgical anatomy of the choroidal arteries: fourth ventricle and cerebellopontine angles. J Neurosurg. 1980;52:504-524.
Gibo H, Caarver CC, Rhoton ALJr. Microsurgical anatomy of the middle cerebral artery. J Neurosurg. 1981;54:151-169.
Gibo H, Lenkey C, Rhoton ALJr. Microsurgical anatomy of the supraclinoid portion of the internal carotid artery. J Neurosurg. 1981;55:560-574.
Hardy DG, Peace D, Rhoton ALJr. Microsurgical anatomy of the superior cerebellar artery. Neurosurgery. 1980;6:10-28.
Huang YP, Wolf BS. Veins of the posterior fossa. In: Newton TH, Potts DG, editors. Radiology of the Skull and Brain, Vol 2. St Louis: CV Mosby; 1974:2155-2216. book 3
Huang YP, Wolf BS. The basal cerebral vein and its tributaries. In: Newton TH, Potts DG, editors. Radiology of the Skull and Brain, Vol 2. St. Louis: CV Mosby; 1974:2111-2154. book 3
Lister JR, Rhoton ALJr, Matsushima T, et al. Microsurgical anatomy of the posterior inferior cerebellar artery. Neurosurgery. 1982;10:170-199.
Martin RG, Grant JL, Peace D, et al. Microsurgical relationships of the anterior inferior cerebellar artery and the facial-vestibulocochlear nerve complex. Neurosurgery. 1980;6:483-507.
Matsushima T, Rhoton ALJr, Lenkey C. Microsurgery of the fourth ventricle: Part 1: microsurgical anatomy. Neurosurgery. 1982;11:631-667.
Nagata S, Rhoton ALJr, Barry M. Microsurgical anatomy of the choroidal fissure. Surg Neurol. 1988;30:3-59.
Oka K, Rhoton ALJr, Barry M, et al. Microsurgical anatomy of the superficial veins of the cerebrum. Neurosurgery. 1985;17:711-748.
Ono M, Kubik S, Abernathey CD. Atlas of the Cerebral Sulci. Stuttgart, Germany: Georg Thieme Verlag; 1990.
Ono M, Ono M, Rhoton ALJr. Microsurgical anatomy of the region of the tentorial incisura. J Neurosurg. 1984;60:365-399.
Ono M, Rhoton ALJr, Peace D, et al. Microsurgical anatomy of the deep venous system of the brain. Neurosurgery. 1984;15:621-657.
Rhoton ALJr. Microsurgical anatomy of the third ventricular region. In: Apuzzo MLJ, editor. Surgery of the Third Ventricle Region. Baltimore: Williams & Wilkins; 1987:570-590.
Rhoton ALJr, Fujii K, Fradd B. Microsurgical anatomy of the anterior choroidal artery. Surg Neurol. 1979;12:171-187.
Rosner SS, Rhoton ALJr, Ono M, et al. Microsurgical anatomy of the anterior perforating arteries. J Neurosurg. 1984;61:468-485.
Taveras JM, Wood EH. Diagnotic Neuroradiology. Baltimore: Williams & Wilkins; 1964.
Timurkaynak E, Rhoton ALJr, Barry M. Microsurgical anatomy and operative approaches to the lateral ventricles. Neurosurgery. 1986;19:685-723.
Wen HT, Rhoton ALJr, de Oliveira E, et al. Microsurgical anatomy of the temporal lobe: part 1. Mesial temporal lobe and its vascular relationship as applied to amygdalohippocampectomy. Neurosurgery. 1999;45:549-592.
Wen HT, Rhoton ALJr, de Oliveira E. Transchoroidal approach to the third ventricle: an anatomic study of the choroidal fissure and its clinical application. Neurosurgery. 1998;42:1205-1219.
Wolf BS, Huang YP. The insula and deep middle cerebral venous drainage system: normal anatomy and angiography. Am J Roentgenol Radium Ther Nucl Med. 1963;90:472-489.
Yamamoto I, Rhoton ALJr, Peace D. Microsurgery of the third ventricle: Part 1. Microsurgical anatomy. Neurosurgery. 1981;8:334-356.
Yasargil MG. Microneurosurgery: Microsurgical Anatomy of the Basal Cisterns and Vessels of the Brain, vol 1. Stuttgart, Germany: Georg Thieme Verlag; 1984.
Zeal AA, Rhoton ALJr. Microsurgical anatomy of the posterior cerebral artery. J Neurosurg. 1978;48:534-559.
1 Ono M, Kubik S, Abernathey CD. Atlas of the Cerebral Sulci. Stuttgart, Germany: Georg Thieme Verlag; 1990.
2 Wolf BS, Huang YP. The insula and deep middle cerebral venous drainage system: normal anatomy and angiography. Am J Roentgenol Radium Ther Nucl Med. 1963;90:472-489.
3 Gibo H, Caarver CC, Rhoton ALJr. Microsurgical anatomy of the middle cerebral artery. J Neurosurg. 1981;54:151-169.
4 Timurkaynak E, Rhoton ALJr, Barry M. Microsurgical anatomy and operative approaches to the lateral ventricles. Neurosurgery. 1986;19:685-723.
5 Wen HT, Rhoton ALJr, de Oliveira E, et al. Microsurgical anatomy of the temporal lobe: part 1. Mesial temporal lobe and its vascular relationship as applied to amygdalohippocampectomy. Neurosurgery. 1999;45:549-592.
6 Yamamoto I, Rhoton ALJr, Peace D. Microsurgery of the third ventricle: Part 1. microsurgical anatomy. Neurosurgery. 1981;8:334-356.
7 Nagata S, Rhoton ALJr, Barry M. Microsurgical anatomy of the choroidal fissure. Surg Neurol. 1988;30:3-59.
8 Wen HT, Rhoton ALJr, de Oliveira E. Transchoroidal approach to the third ventricle: an anatomic study of the choroidal fissure and its clinical application. Neurosurgery. 1998;42:1205-1219.
9 Rhoton ALJr. Microsurgical anatomy of the third ventricular region. In: Apuzzo MLJ, editor. Surgery of the Third Ventricle Region. Baltimore: Williams & Wilkins; 1987:570-590.
10 Ono M, Ono M, Rhoton ALJr. Microsurgical anatomy of the region of the tentorial incisura. J Neurosurg. 1984;60:365-399.
11 Gibo H, Lenkey C, Rhoton ALJr. Microsurgical anatomy of the supraclinoid portion of the internal carotid artery. J Neurosurg. 1981;55:560-574.
12 Rosner SS, Rhoton ALJr, Ono M, et al. Microsurgical anatomy of the anterior perforating arteries. J Neurosurg. 1984;61:468-485.
13 Taveras JM, Wood EH. Diagnotic Neuroradiology. Baltimore: Williams & Wilkins; 1964.
14 Oka K, Rhoton ALJr, Barry M, et al. Microsurgical anatomy of the superficial veins of the cerebrum. Neurosurgery. 1985;17:711-748.
15 Ono M, Rhoton ALJr, Peace D, et al. Microsurgical anatomy of the deep venous system of the brain. Neurosurgery. 1984;15:621-657.
16 Rhoton ALJr, Fujii K, Fradd B. Microsurgical anatomy of the anterior choroidal artery. Surg Neurol. 1979;12:171-187.
17 Zeal AA, Rhoton ALJr. Microsurgical anatomy of the posterior cerebral artery. J Neurosurg. 1978;48:534-559.
18 Huang YP, Wolf BS. The basal cerebral vein and its tributaries. In: Newton TH, Potts DG, editors. Radiology of the Skull and Brain, Vol 2. St. Louis: CV Mosby; 1974:2111-2154. book 3
19 Yasargil MG. Microneurosurgery: Microsurgical Anatomy of the Basal Cisterns and Vessels of the Brain, vol 1. Stuttgart, Germany: Georg Thieme Verlag; 1984.
20 Matsushima T, Rhoton ALJr, Lenkey C. Microsurgery of the fourth ventricle: part 1: microsurgical anatomy. Neurosurgery. 1982;11:631-667.
21 Fujii K, Lenkey C, Rhoton ALJr. Microsurgical anatomy of the choroidal arteries: fourth ventricle and cerebellopontine angles. J Neurosurg. 1980;52:504-524.
22 Huang YP, Wolf BS. Veins of the posterior fossa. In: Newton TH, Potts DG, editors. Radiology of the Skull and Brain, Vol 2. St Louis: CV Mosby; 1974:2155-2216. book 3
23 Lister JR, Rhoton ALJr, Matsushima T, et al. Microsurgical anatomy of the posterior inferior cerebellar artery. Neurosurgery. 1982;10:170-199.
24 Martin RG, Grant JL, Peace D, et al. Microsurgical relationships of the anterior inferior cerebellar artery and the facial-vestibulocochlear nerve complex. Neurosurgery. 1980;6:483-507.
25 Hardy DG, Peace D, Rhoton ALJr. Microsurgical anatomy of the superior cerebellar artery. Neurosurgery. 1980;6:10-28.