[level-membership-for-basic-science-category]Chapter 5
Formation of Germ Layers and Early Derivatives
As it is implanting into the uterine wall, the embryo undergoes profound changes in its organization. Up to the time of implantation, the blastocyst consists of the inner cell mass, from which the body of the embryo proper arises, and the outer trophoblast, which represents the future tissue interface between the embryo and mother. Both components of the blastocyst serve as the precursors of other tissues that appear in subsequent stages of development. Chapter 3 discusses the way in which the cytotrophoblast gives rise to an outer syncytial layer, the syncytiotrophoblast, shortly before attaching to uterine tissue (see Fig. 3.18). Not long thereafter, the inner cell mass begins to give rise to other tissue derivatives as well. The subdivision of the inner cell mass ultimately results in an embryonic body that contains the three primary embryonic germ layers: the ectoderm (outer layer), mesoderm (middle layer), and endoderm (inner layer). The process by which the germ layers are formed through cell movements is called gastrulation.
Two-Germ-Layer Stage
Just before the embryo implants into the endometrium early in the second week, significant changes begin to occur in the inner cell mass and in the trophoblast. As the cells of the inner cell mass become rearranged into an epithelial configuration, sometimes referred to as the embryonic shield, a thin layer of cells appears ventral to the main cellular mass (see Fig. 3.18). The main upper layer of cells is known as the epiblast, and the lower layer is called the hypoblast, or primitive endoderm (Fig. 5.1).
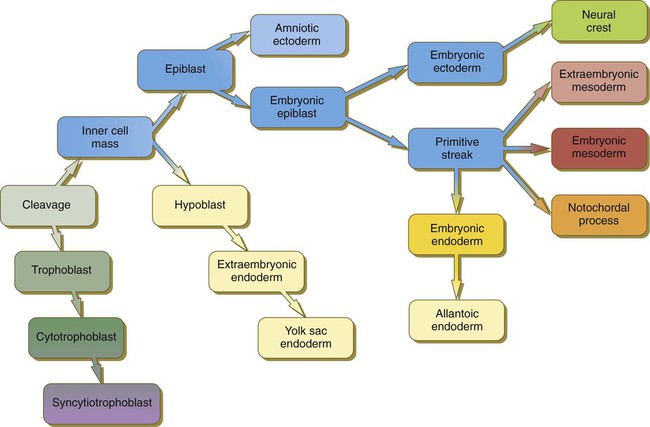
(Note: The colors in the boxes are found in all illustrations involving the embryonic and extraembryonic germ layers.)
How the hypoblast forms in human embryos is not understood, but studies on mouse embryos have shown that as early as the 64-cell stage, some cells of the inner cell mass express the transcription factor nanog, whereas others express Gata 6. These cells are arranged in a salt and pepper pattern within the inner cell mass (Fig. 5.2A). The nanog-expressing cells represent the precursors of the epiblast, and those expressing Gata 6 will become the hypoblast. The basis for the differentiation of these two distinct precursor cell types is not completely understood, but according to the “time inside–time outside” hypothesis, those cells that enter the inner cell mass earliest are biased to express nanog, which perpetuates their pluripotency. Possibly because of the influence of fibroblast growth factor-4 (FGF-4), secreted by these first arrivals to the inner cell mass, later immigrants are then biased to express Gata 6. The Gata 6–expressing cells produce molecules that increase their adhesive properties, as well as their mobility, and they make their way to the lower surface of the inner cell mass to form a thin epithelium, the hypoblast. Those Gata 6 cells that fail to reach the surface of the inner cell mass undergo apoptosis (cell death). The nanog-expressing cells of the inner cell mass also assume an epithelial configuration as they form the epiblast. Between the epiblast and hypoblast a basal lamina forms.
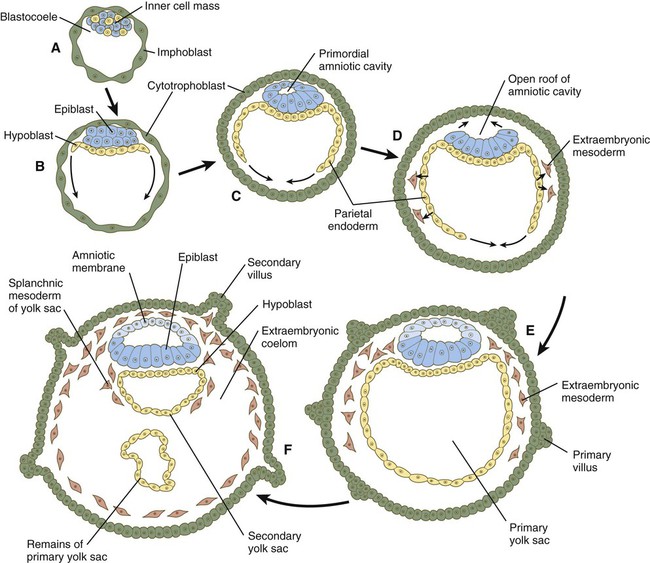
The syncytiotrophoblast is not shown. A, Late blastocyst. Within the inner cell mass, blue nanog-expressing pre-epiblastic cells and yellow Gata 6–expressing prehypoblastic cells are mixed in a salt and pepper pattern. B, Beginning of implantation at 6 days. The hypoblast has formed and is beginning to spread beneath the cytotrophoblast as the parietal endoderm. C, Implanted blastocyst at
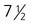 days. D, Implanted blastocyst at 8 days. E, Embryo at 9 days. F, Late second week.
days. D, Implanted blastocyst at 8 days. E, Embryo at 9 days. F, Late second week.A small group of hypoblast cells that becomes translocated to the future anterior end of the embryo (called anterior visceral endoderm by mouse embryologists) has been shown to possess remarkable signaling powers. The cells first secrete the signaling molecules, lefty-1 and Cerberus-1 (Cer-1), which inhibit the activity of the signaling molecules, nodal and Wnt, in the overlying epiblast but allow nodal and Wnt-3 expression in the posterior epiblast (see Fig. 5.8A). (Nodal signaling from the posterior epiblast stimulates the initial formation of the anterior visceral endoderm.) This represents the first clear expression of anteroposterior polarity in the embryo. It also forms two signaling domains within the early embryo. The anterior visceral endoderm soon begins to induce much of the head and forebrain and inhibits the formation of posterior structures. In the posterior part of the epiblast, nodal signaling activity stimulates the formation of the primitive streak (see next section), which is the focal point for gastrulation and germ layer formation. After the hypoblast has become a well-defined layer, and the epiblast has taken on an epithelial configuration, the former inner cell mass is transformed into a bilaminar disk, with the epiblast on the dorsal surface and the hypoblast on the ventral surface.
The epiblast contains the cells that make up the embryo itself, but extraembryonic tissues also arise from this layer. The next layer to appear after the hypoblast is the amnion, a layer of extraembryonic ectoderm that ultimately encloses the entire embryo in a fluid-filled chamber called the amniotic cavity (see Chapter 7). Because of the paucity of specimens, the earliest stages in the formation of the human amnion and amniotic cavity are not completely understood. Studies on primate embryos indicate that a primordial amniotic cavity first arises by cavitation (formation of an internal space) within the pre-epithelial epiblast; it is covered by cells derived from the inner cell mass (see Fig. 5.2). According to some investigators, the roof of the amnion then opens, thus exposing the primordial amniotic cavity to the overlying cytotrophoblast. Soon thereafter (by about 8 days after fertilization), the original amniotic epithelium reforms a solid roof over the amniotic cavity.
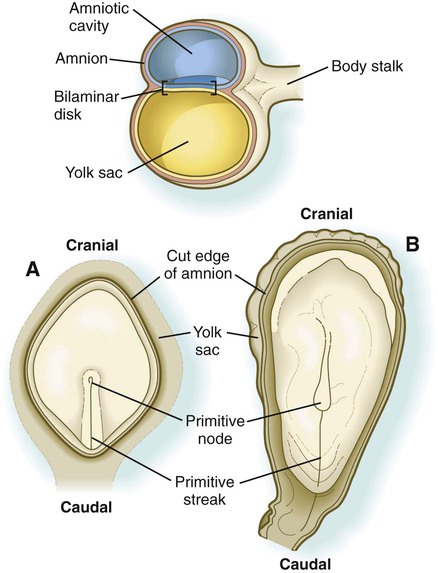
While the early embryo is still sinking into the endometrium (about 9 days after fertilization), cells of the hypoblast begin to spread and line the inner surface of the cytotrophoblast with a continuous layer of extraembryonic endoderm called parietal endoderm (Fig. 5.3; see Fig. 5.2). When the endodermal spreading is completed, a vesicle called the primary yolk sac has taken shape (see Fig. 3.18C). At this point (about 10 days after fertilization), the embryo complex constitutes the bilaminar germ disk, which is located between the primary yolk sac on its ventral surface and the amniotic cavity on its dorsal surface (Fig. 5.4). Shortly after it forms, the primary yolk sac becomes constricted, forming a secondary yolk sac and leaving behind a remnant of the primary yolk sac (see Figs. 3.18D and 5.2F).
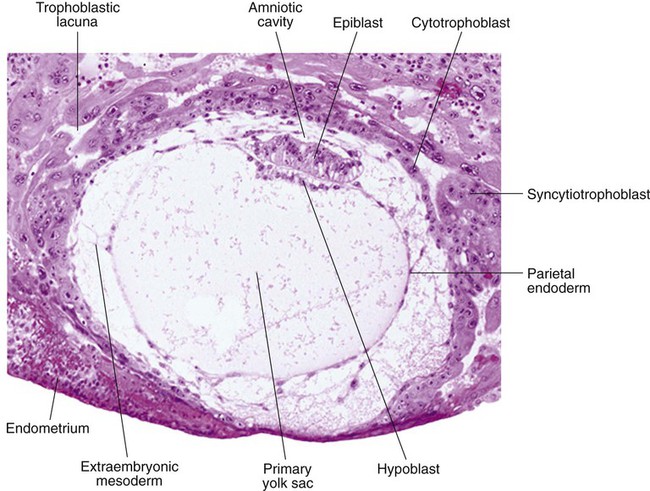
Starting at about 12 days after fertilization, another extraembryonic tissue, the extraembryonic mesoderm, begins to appear (see Fig. 5.2). The first extraembryonic mesodermal cells seem to arise from a transformation of parietal endodermal cells. These cells are later joined by extraembryonic mesodermal cells that have originated from the primitive streak. The extraembryonic mesoderm becomes the tissue that supports the epithelium of the amnion and yolk sac and the chorionic villi, which arise from the trophoblastic tissues (see Chapter 7). The support supplied by the extraembryonic mesoderm is not only mechanical, but also trophic because the mesoderm serves as the substrate through which the blood vessels supply oxygen and nutrients to the various epithelia.
Gastrulation and the Three Embryonic Germ Layers
At the end of the second week, the embryo consists of two flat layers of cells: the epiblast and the hypoblast. As the third week of pregnancy begins, the embryo enters the period of gastrulation, during which the three embryonic germ layers form from the epiblast (see Fig. 5.1). The morphology of human gastrulation follows the pattern seen in birds. Because of the large amount of yolk in birds’ eggs, the avian embryo forms the primary germ layers as three overlapping flat disks that rest on the yolk, similar to a stack of pancakes. Only later do the germ layers fold to form a cylindrical body. Although the mammalian egg is essentially devoid of yolk, the morphological conservatism of early development still constrains the human embryo to follow a pattern of gastrulation similar to that seen in reptiles and birds. Because of the scarcity of material, even the morphology of gastrulation in human embryos is not known in detail. Nevertheless, extrapolation from avian and mammalian gastrulation can provide a reasonable working model of human gastrulation.
Gastrulation begins with the formation of the primitive streak, a linear midline condensation of cells derived from the epiblast in the posterior region of the embryo through an induction by cells at the edge of the embryonic disk in that region (see Fig. 5.4). Members of the transforming growth factor-β (TGF-β) and Wnt families of signaling molecules have been identified as likely inducing agents. Initially triangular, the primitive streak soon becomes linear and elongates, largely through a combination of proliferation and migration, as well as internal cellular rearrangements, called convergent-extension movements. With the appearance of the primitive streak, the anteroposterior (craniocaudal) and right-left axes of the embryo can be readily identified (see Fig. 5.4).
The primitive streak is a region where cells of the epiblast converge in a well-defined spatial and temporal sequence. As cells of the epiblast reach the primitive streak, they change shape and pass through it on their way to forming new layers beneath (ventral to) the epiblast (Fig. 5.5C). Marking studies have shown that cells entering the primitive streak form distinct lineages as they leave. The most posterior cells both to enter and leave the streak as it is beginning to elongate form the extraembryonic mesoderm lining the trophoblast and yolk sac, as well as that forming the blood islands (see Fig. 6.19). Another wave of mesoderm, arising later and more anteriorly in the primitive streak, forms the paraxial, lateral plate, and cardiac mesoderm. A final wave, which enters and leaves the anteriormost end of the primitive streak, gives rise to midline axial structures (the notochord, the prechordal plate, and the primitive node itself) and also the embryonic endoderm. The composite results of such marking experiments are organized into fate maps, such as that illustrated in Figure 5.5A.
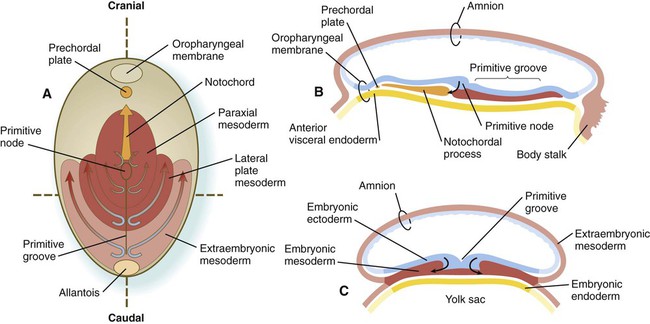
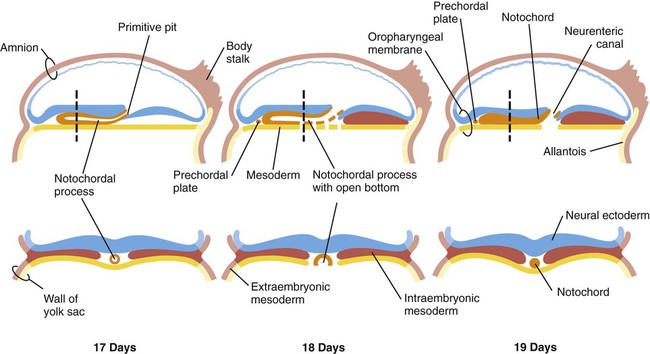
The endodermal precursor cells that pass through the anterior primitive streak largely displace the original hypoblast, but research has shown that some of the original hypoblastic cells become integrated into the newly forming embryonic endodermal layer. The displaced hypoblastic cells form extraembryonic endoderm. The movement of cells through the primitive streak results in the formation of a groove (primitive groove) along the midline of the primitive streak. At the anterior end of the primitive streak is a small but well-defined accumulation of cells, called primitive node, or Hensen’s node.* This structure is of great developmental significance because, in addition to being the major posterior signaling center of the embryo (Box 5.1), it is the area through which cells migrate in a stream toward the anterior end of the embryo. These cells, called mesendoderm, soon segregate into a rodlike mesodermal notochord and the endodermal dorsal wall of the forming gut. Anterior to the notochord is a group of mesodermal cells called the prechordal plate (see Fig. 5.5A and B). (The important functions of the notochord and prechordal plate are discussed on p. 80.)
Box 5.1 Molecular Aspects of Gastrulation
For this aspect of early development, we must rely almost entirely on studies of mouse embryos. The original symmetry of the embryo is broken by the displacement of the future anterior visceral endoderm to the anterior side of the embryonic disk. This is a function of proliferation and later migration of the cells that will constitute the anterior visceral endoderm. Migration of these cells (and the resulting establishment of the anteroposterior axis) depends on the activation of the Wnt antagonist Dkk 1 (Dickkopf 1) in the future anterior part of the embryo. This confines Wnt activity to the future posterior part of the embryo, where it induces the expression of the signaling molecule Nodal (Fig. 5.8A). When the anterior visceral endoderm has become stabilized in the anterior part of the embryonic disk, it produces the Nodal inhibitors lefty-1 and Cer-1, which confine Nodal activity to the posterior end of the embryo where, responding to extraembryonic Wnt signals, it establishes a posterior signaling center, which induces the formation of the primitive streak, the definitive endoderm, and the mesoderm. In the chick embryo, the ectopic application of two other signaling molecules, chordin and Vg1, induces the formation of an ectopic primitive streak.
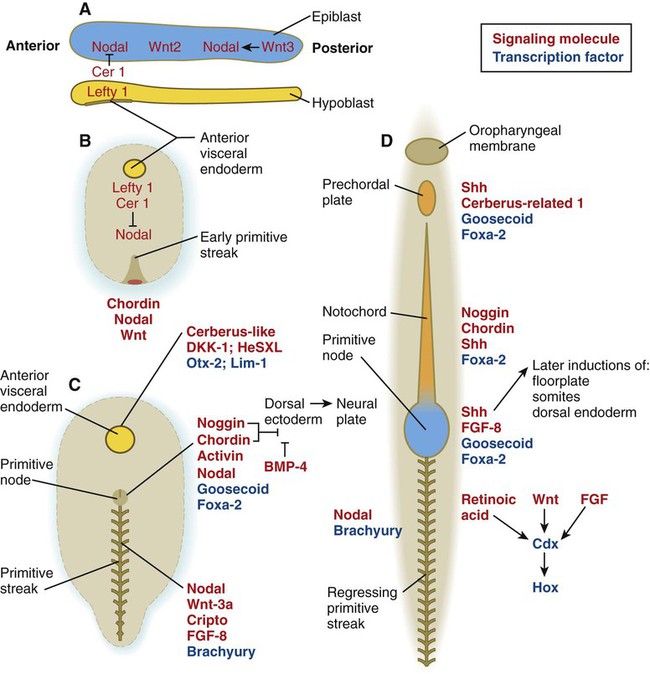
A, Preprimitive streak (sagittal section). B, Early formation of the primitive streak. C, Gastrulation (period of germ layer formation). D, Late gastrulation and neural induction. The molecules in red are signaling molecules, and the molecules in blue are transcription factors. Names of specific molecules (bold) are placed by the structures in which they are expressed.
As the primitive streak elongates, migrating cells of the epiblast join the tip of the streak, and a dynamic mass of cells, called the primitive node, becomes evident at the tip of the primitive streak. Cells of the node express many genes, including three classic molecular markers of the organizer region in many vertebrates—chordin, goosecoid, and hepatic nuclear factor-3β (now called Foxa-2). Not only is the winged helix transcription factor, Foxa-2, important for the formation of the node itself, but also it is vital for the establishment of midline structures cranial to the node. Foxa-2 is required for the initiation of notochord function. In its absence, the notochord and the floor plate of the neural tube (see Chapter 11) fail to form. In contrast, endoderm, the primitive streak, and intermediate mesoderm do develop. Goosecoid, a homeodomain transcription factor, is prominently expressed in the organizer region of all vertebrates studied. Goosecoid activates chordin, noggin, and other genes of the organizer region. If ectopically expressed, it stimulates the formation of a secondary body axis. Chordin and noggin, signaling molecules associated with the node, are involved with neural induction, and expression of nodal on the left side of the embryo is a key element in the setting of left-right asymmetry.
Two genes, T and nodal, play prominent roles in the function of the primitive streak and posterior mesoderm formation. Expression of the T gene seems to be activated by products of the Foxa-2 and goosecoid genes. In T mutants (brachyury), the notochord begins to form through the activity of Foxa-2, but it fails to complete development. Studies on T mutants have shown that activity of the brachyury gene is necessary for normal movements of future mesodermal cells through the primitive streak during gastrulation. In brachyury (short tail) mutant mice, mesodermal cells pile up at a poorly formed primitive streak, and the embryos show defective elongation of the body axis (including a short tail) posterior to the forelimbs. T gene mutants may be responsible for certain gross caudal body defects in humans. Nodal, a member of the transforming growth factor-β (TGF-β) family of growth factor genes (see Table 4.1), is expressed throughout the posterior epiblast before gastrulation, but its activity is concentrated at the primitive node during gastrulation. Similar to the brachyury gene, the effects of nodal are strongly seen in the caudal region of the embryo. In the null mutant of nodal, the primitive streak fails to form, and the embryo is deficient in mesoderm. Similarly, mutants of cripto, an early-acting member of the epidermal growth factor family and an essential cofactor in the nodal signaling pathway, produce a trunkless phenotype.
As cells pass through the primitive streak, a region of Hox gene expression begins to form around the streak. The pattern of Hox gene expression in the future trunk and posterior part of the embryo is based on signaling by three molecules—retinoic acid, Wnt, and FGF—that act on the transcription factor Cdx (the mammalian equivalent of caudal in Drosophila) in the area of the regressing primitive streak just behind the last-forming somites. Cdx acts on the Hox genes, which impose unique characteristics to the segmental structures that form along the anteroposterior axis of the embryo (Fig. 5.8D).
Anterior Visceral Endoderm (Hypoblast)
In mammals, even before mesodermal cells begin migrating through the primitive node, the anterior hypoblast (called the anterior visceral endoderm by mouse embryologists) expresses genes characteristic of the prechordal plate and initiates head formation. The anterior visceral endoderm itself is subdivided into an anterior part, which serves as a signaling center for early heart formation (see p. 104), and a more posterior part, which becomes part of the prechordal plate complex and induces formation of the head. According to one model, induction of the head and forebrain in mammals is a two-step process, in which an early induction by the anterior visceral endoderm confers a labile anterior character to the head and brain, and a later induction by the prechordal plate mesoderm reinforces and maintains this induction.
A major function of the anterior visceral endoderm is to emit molecular signals that inhibit the development of posterior embryonic structures. To produce a head, it is necessary to block the bone morphogenetic protein-4 (BMP-4) signal (by noggin) and a Wnt signal (by Dkk-1). Signaling molecules and transcription factors are produced in the head signaling centers. In mice bearing mutants of Lim-1 (Lhx-1), a homeobox-containing transcription factor, and cereberus-like 1, a signaling molecule, headless mice are born (Fig. 5.9). The headless mice are born without neural structures anterior to rhombomere 3 (see Fig. 6.3). Otx-2, another transcription factor present in the head signaling center, is also a general marker of the induced anterior region of the central nervous system. Many other molecules are also expressed in the head signaling center. How these orchestrate the formation of the head remains to be determined.

The specific craniocaudal characteristics of the structures arising from the newly formed paraxial mesoderm are specified by patterns of Hox gene expression, first in the epiblast and then in the mesodermal cells themselves. The transformations of morphology and the behavior of the cells passing through the primitive streak are associated with profound changes not only in their adhesive properties and internal organization, but also in the way that they relate to their external environment. Much of the extraembryonic mesoderm forms the body stalk, which connects the caudal part of the embryo to the extraembryonic tissues that surround it (see Figs. 5.4 and 7.1). The body stalk later becomes the umbilical cord.
The movements of the cells passing through the primitive streak are accompanied by major changes in their structure and organization (Fig. 5.6). While in the epiblast, the cells have the properties of typical epithelial cells, with well-defined apical and basal surfaces, and they are associated with a basal lamina that underlies the epiblast. As they enter the primitive streak, these cells elongate, lose their basal lamina, and take on a characteristic morphology that has led to their being called bottle cells. When they become free of the epiblastic layer in the primitive groove, the bottle cells assume the morphology and characteristics of mesenchymal cells, which are able to migrate as individual cells if they are provided with the proper extracellular environment (see Fig. 5.6). Included in this transformation is the loss of specific cell adhesion molecules (CAMs), in particular E-cadherin (see p. 254) as the cells convert from an epithelial to a mesenchymal configuration. This transformation is correlated with the expression of the transcription factor snail, which is also active in the separation of mesenchymal neural crest cells from the epithelial neural tube (see p. 254). As cells in the epiblast are undergoing epithelial-mesenchymal transition, they begin to express the CAM N-cadherin, which is necessary for their spreading out from the primitive streak in the newly forming mesodermal layer.
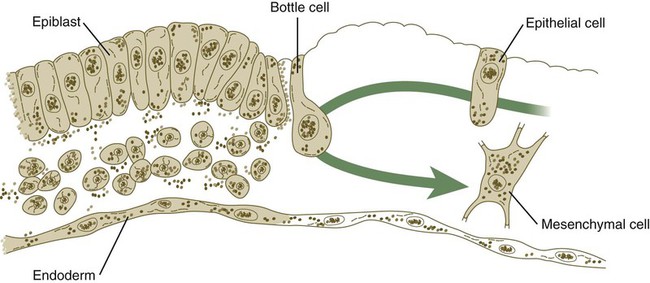
Changes in the shape of a cell as it migrates along the epiblast (epithelium), through the primitive streak (bottle cell), and away from the groove as a mesenchymal cell that will become part of the mesodermal germ layer. The same cell can later assume an epithelial configuration as part of a somite.
Starting in early gastrulation, cells of the epiblast produce hyaluronic acid, which enters the space between the epiblast and hypoblast. Hyaluronic acid, a polymer consisting of repeating subunits of D-glucuronic acid and N-acetylglucosamine, is frequently associated with cell migration in developing systems. The molecule has a tremendous capacity to bind water (up to 1000 times its own volume), and it functions to keep mesenchymal cells from aggregating during cell migrations. Although after leaving the primitive streak the mesenchymal cells of the embryonic mesoderm find themselves in a hyaluronic acid–rich environment, hyaluronic acid alone is not enough to support their migration from the primitive streak. In all vertebrate embryos that have been investigated to date, the spread of mesodermal cells away from the primitive streak or the equivalent structure is found to depend on the presence of fibronectin associated with the basal lamina beneath the epiblast. The embryonic mesoderm ultimately spreads laterally as a thin sheet of mesenchymal cells between the epiblast and hypoblast layers (see Fig. 5.5C).
Regression of the Primitive Streak
After its initial appearance at the extreme caudal end of the embryo, the primitive streak expands cranially until about 18 days after fertilization (see Fig. 5.4). Thereafter, it regresses caudally (see Fig. 5.11) and strings out the notochord in its wake. Vestiges remain into the fourth week. During that time, the formation of mesoderm continues by means of cells migrating from the epiblast through the primitive groove. Regression of the primitive streak is accompanied by the establishment and patterning of the paraxial mesoderm (see p. 97), which gives rise to the somites and ultimately the segmental axial structures of the trunk and caudal regions of the body. As regression of the primitive streak comes to a close, its most caudal extent is marked by a mass of mesenchymal cells, which form the tail bud. This structure plays an important role in forming the most posterior portion of the neural tube (see p. 93).
The primitive streak normally disappears without a trace, but in rare instances, large tumors called teratomas appear in the sacrococcygeal region (see Fig. 1.2A). Teratomas often contain bizarre mixtures of many different types of tissue, such as cartilage, muscle, fat, hair, and glandular tissue. Because of this, sacrococcygeal teratomas are thought to arise from remains of the primitive streak (which can form all germ layers). Teratomas also are found in the gonads and the mediastinum. These tumors are thought to originate from germ cells.
Notochord and Prechordal Plate
Cranial to the notochord is a small region where embryonic ectoderm and endoderm abut without any intervening mesoderm. Called the oropharyngeal membrane (see Fig. 5.5), this structure marks the site of the future oral cavity. Between the cranial tip of the notochordal process and the oropharyngeal membrane is a small aggregation of mesodermal cells closely apposed to endoderm, called the prechordal plate (see Fig. 5.5). In birds, the prechordal plate emits molecular signals that are instrumental in stimulating the formation of the forebrain, similar to the anterior visceral endoderm in mammals.
Both the prechordal plate and the notochord arise from the ingression of a population of epiblastic cells, which join other cells of primitive streak origin, within the primitive node. As the primitive streak regresses, the cellular precursors of first the prechordal plate and then the notochord migrate rostrally from the node, but they are left behind as a rodlike aggregation of cells (notochordal process; see Fig. 5.5A and B) in the wake of the regressing primitive streak. In mammals, shortly after ingression, the cells of the notochordal process temporarily spread out and fuse with the embryonic endoderm (Fig. 5.7). The result is the formation of a transitory neurenteric canal that connects the emerging amniotic cavity with the yolk sac. Later, the cells of the notochord separate from the endodermal roof of the yolk sac and form the definitive notochord, a solid rod of cells in the midline between the embryonic ectoderm and endoderm (see Fig. 5.7).
Induction of the Nervous System
Neural Induction
Deletion and transplantation experiments in amphibians set the stage for the present understanding of neural induction. (See Chapters 6 and 11 for further details on the formation of the nervous system.) In the absence of chordamesoderm moving from the dorsal lip of the blastopore (the amphibian equivalent of the primitive node), the nervous system does not form from the dorsal ectoderm. In contrast, if the dorsal lip of the blastopore is grafted beneath the belly ectoderm of another host, a secondary nervous system and body axis form in the area of the graft (Fig. 5.10).
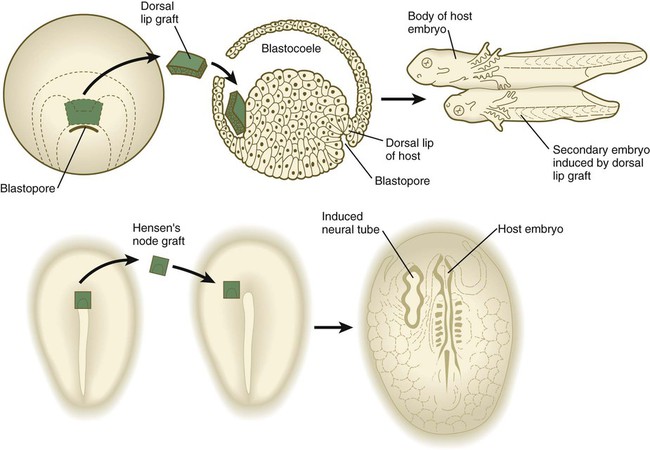
Top, Graft of the dorsal lip of the blastopore in a salamander embryo induces a secondary embryo to form. Bottom, Graft of Hensen’s node from one avian embryo to another induces the formation of a secondary neural tube. (Top based on studies by Spemann H: Embryonic development and induction, New York, 1938, Hafner; Bottom based on studies by Waddington C: J Exp Biol 10:38-46, 1933.)
The dorsal lip has been called the organizer because of its ability to stimulate the formation of a secondary body axis. Subsequent research has shown that the interactions occurring in the region of the dorsal lip in amphibians are far more complex than a single induction between chordamesoderm and ectoderm. Deletion and transplantation experiments have also been conducted on embryos of birds and mammals (see Fig. 5.10); clearly, the primitive node and the notochordal process in birds and mammals are homologous in function to the dorsal lip and chordamesoderm in amphibians. This means that, in higher vertebrates, the primitive node and the notochordal process act as the neural inductor, and the overlying ectoderm is the responding tissue. Over the years, embryologists have devoted an enormous amount of research to identifying the nature of the inductive signal that passes from the chordamesoderm to the ectoderm.
Early Formation of the Neural Plate
The first obvious morphological response of the embryo to neural induction is the transformation of the dorsal ectoderm overlying the notochordal process into an elongated patch of thickened epithelial cells called the neural plate (Fig. 5.11). The border of the neural plate is specified by exposure of those cells to a certain concentration of BMP. This is the region from which the neural crest (see p. 254) arises.
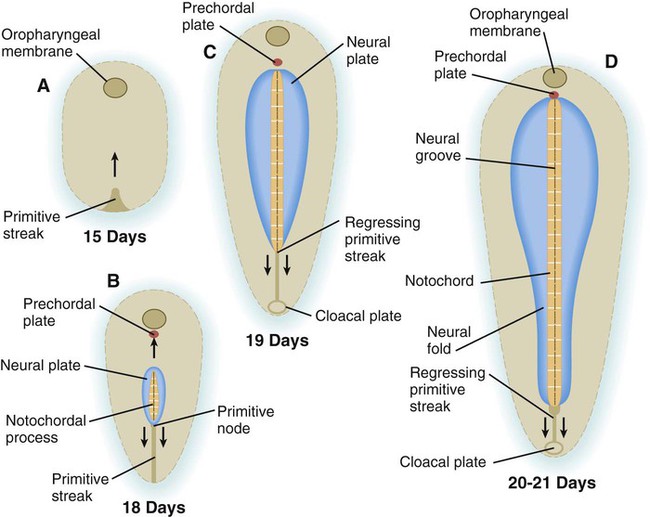
A, Day 15. B, Day 18. C, Day 19. D, Days 20 to 21.
As development progresses, certain decisions are made that narrow the developmental options of cells (Fig. 5.12). For example, at an early stage in cleavage, some cells become committed to the extraembryonic trophoblastic line and are no longer capable of participating in the formation of the embryo itself. At the point at which cells are committed to becoming trophoblast, a restriction event has occurred. When a group of cells has gone through its last restriction event (e.g., the transition from cytotrophoblast to syncytiotrophoblast), their fate is fixed, and they are said to be determined.* These terms, which were coined in the early days of experimental embryology, are now understood to reflect limitations in gene expression as cell lineages follow their normal developmental course. The rare instances in which cells or tissues strongly deviate from their normal developmental course, a phenomenon called metaplasia, are of considerable interest to pathologists and individuals who study the control of gene expression.
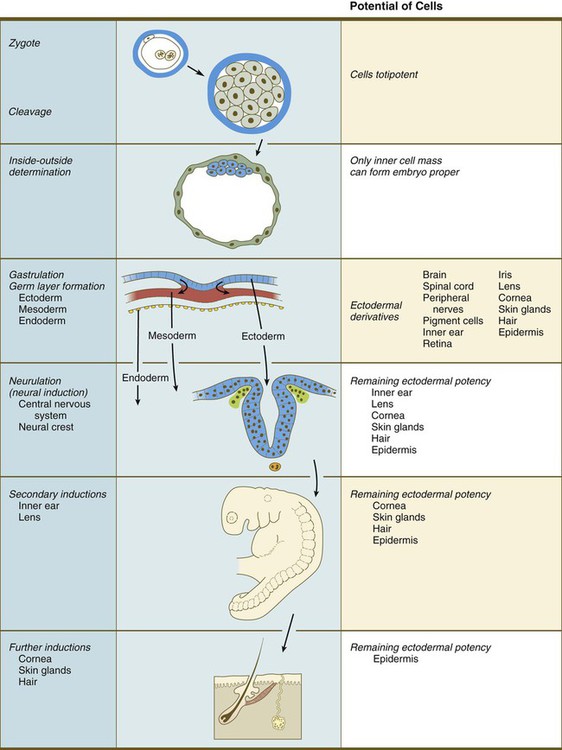
The labels on the right illustrate the progressive restriction of the developmental potential of cells that are in the line leading to the formation of the epidermis. On the left are developmental events that remove groups of cells from the epidermal track.
Box 5.2 Molecular Basis for Left-Right Asymmetry
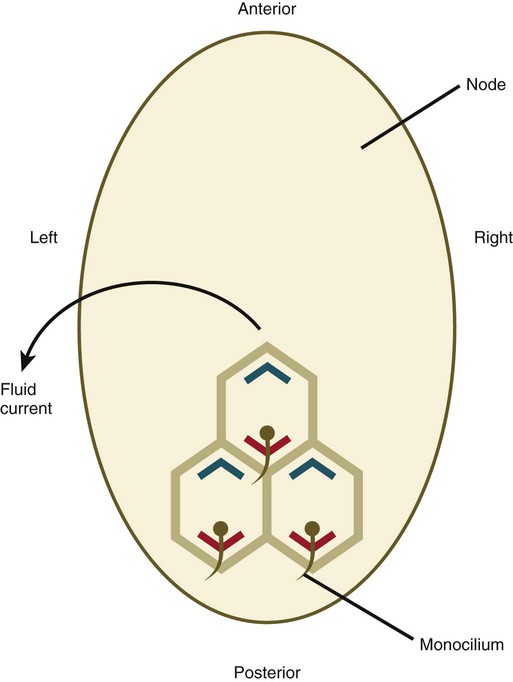
Up to the time of gastrulation, the embryo is bilaterally symmetrical, but at that time mechanisms are set in place that ultimately result in the right-sided looping of the heart, followed by asymmetric looping of the gut and the asymmetric positioning of the liver, spleen, and lobation of the lungs. In mammalian embryos, the earliest known manifestation of asymmetry involves the beating of cilia around the primitive node (Fig. 5.13). This beating results in a directional current leading to the expression of two signaling molecules in the transforming growth factor-β (TGF-β) family—nodal, a symmetry-breaking molecule in the left side of the embryo, and lefty-1 along the left side of the primitive streak—over a very restricted developmental time (from the two-somite to the six-somite stage in the mouse). Lefty-1 may function to prevent the diffusion of left-determining molecules to the right side of the embryo. A sequence of molecular interactions downstream of nodal results in the activation of the Pitx2 gene, a transcription factor, also on the left side. The Pitx2 protein leads to later asymmetric development, such as rotation of the gut and stomach, position of the spleen, and the asymmetric lobation of the lungs. Although the left-sided expression of nodal in the lateral mesoderm seems to be a point of commonality in the determination of left-right asymmetry in all vertebrates, earlier (upstream) molecular events differ among the classes of vertebrates. In the chick, important signaling molecules, such as sonic hedgehog (shh) and fibroblast growth factor-8 (FGF-8), are asymmetrically distributed around the node, whereas in the mouse, the distribution is uniform.
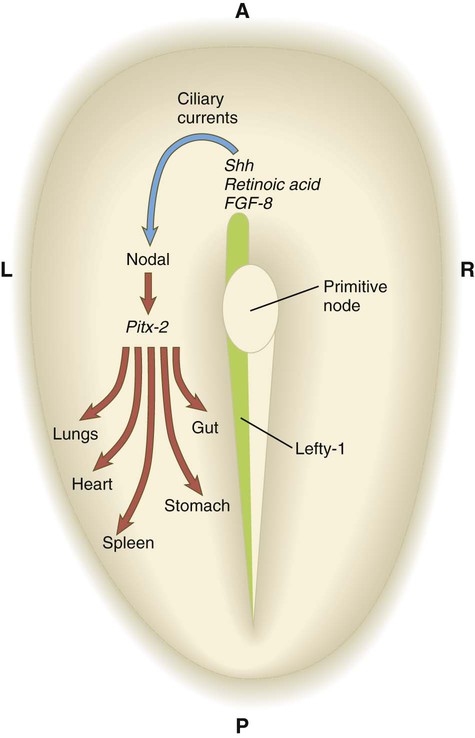
Ciliary currents at the primitive node sweep the symmetry-breaking molecule nodal toward the left side of the embryo, where it stimulates an asymmetric cascade of gene expression via Pitx-2. Lefty-1, expressed along the left side of the embryo, may prevent diffusion of molecules to the right side. Only the most important molecules in a complex cascade are shown. FGF-8, fibroblast growth factor-8; Shh, sonic hedgehog.
How the anteroposterior polarity, exemplified by the primitive streak, is translated via ciliary currents into left-right asymmetry is the subject of considerable research. A likely candidate is planar cell polarity, which is a mechanism directing cells to orient themselves along an axis in the plane of a flat epithelial tissue. This is accomplished by the asymmetric distribution of several planar cell polarity proteins along this axis. In the node, Dishevelled is concentrated in the posterior region of the cells, and a counterpart, Prickle, is arranged along the anterior border (Fig. 5.14). The basal body in each of the 200 to 300 monociliated cells of the node is associated with Dishevelled, and the cilium that protrudes from the cell does so at an angle that produces the leftward fluid current when the cilium beats. There is speculation that a Wnt gradient lies behind the asymmetric distribution of Dishevelled and Prickle, but this remains to be confirmed.
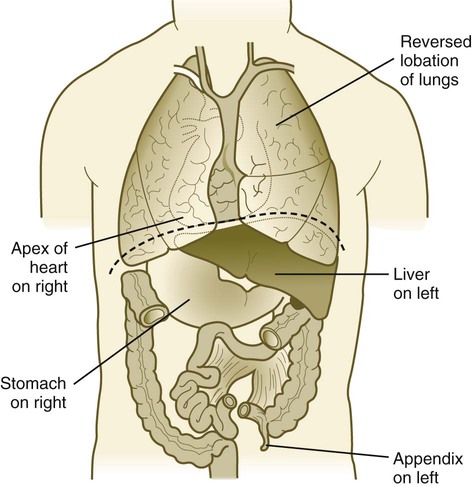
In roughly 1 in 10,000 individuals, the left-right asymmetry of the body is totally reversed, a condition called situs inversus (Fig. 5.15). This condition is often not recognized until the individual is examined relatively late in life by an astute diagnostician. Several mutations and syndromes are associated with this condition, but one of the most instructive is Kartagener’s syndrome, in which situs inversus is associated with respiratory symptoms (sinusitis and bronchiectasis) resulting from abnormalities of the dynein arms in cilia (immotile cilia). In a similar mouse mutant, the cilia around the primitive node do not function properly, and the lack of directionality of the resulting fluid currents around the node is suspected to result in the random localization of nodal and other asymmetry-producing molecules to the right side of the embryo. Partial situs inversus, such as an isolated right-sided heart (dextrocardia), can also occur. With more than 24 genes currently known to be involved in left-right asymmetry, such isolated occurrences of organ asymmetry are probably the result of mutations of genes farther downstream in the asymmetry cascade.
Cell Adhesion Molecules
Contemporary research has provided a molecular basis for many of the cell aggregation and sorting phenomena described by earlier embryologists. Of several families of CAMs that have been described, three are of greatest importance to embryonic development. The first are the cadherins, which are single transmembrane glycoproteins typically arranged as homodimers that protrude from the cell surface. In the presence of calcium (Ca++), cadherin dimers from adjacent cells adhere to one another and cause the cells to become firmly attached to one another (Fig. 5.16). One of the most ubiquitous is E-cadherin, which binds epithelial cells to one another (see Fig. 16.6). During epithelial-mesenchymal transformations, such as that shown in Figure 5.8, the epithelial cells lose their E-cadherins as they transform into mesenchymal cells, but if these cells reform an epithelium later in development, they re-express E-cadherins.
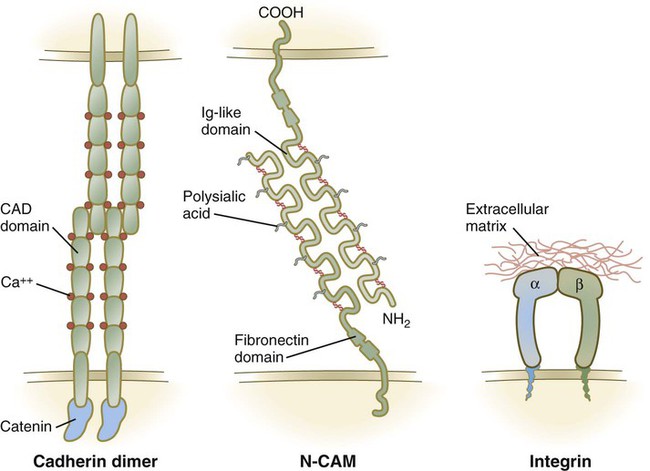
The immunoglobulin Ig (immunoglobulin)-CAMs are characterized by having varying numbers of immunoglobulinlike extracellular domains. These molecules adhere to similar (homophilic binding) or different (heterophilic binding) CAMs on neighboring cells, and they do so without the mediation of calcium ions (see Fig. 5.16). One of the most prominent members of this family is N-CAM, which is strongly expressed within the developing nervous system. Ig-CAMs do not bind cells as tightly as cadherins, and they provide for fine-tuning of intercellular connections. N-CAM is unusual in having a high concentration of negatively charged sialic acid groups in the carbohydrate component of the molecule, and embryonic forms of N-CAM have three times as much sialic acid as the adult form of the molecule.
In the early embryo, before primary induction of the central nervous system, the ectoderm expresses N-CAM and E-cadherin (formerly known as L-CAM). After primary induction, cells within the newly formed neural tube continue to express N-CAM, but they no longer express E-cadherin. They also strongly express N-cadherin. In contrast, the ectoderm ceases to express N-CAM, but it continues to express E-cadherin (Fig. 5.17).
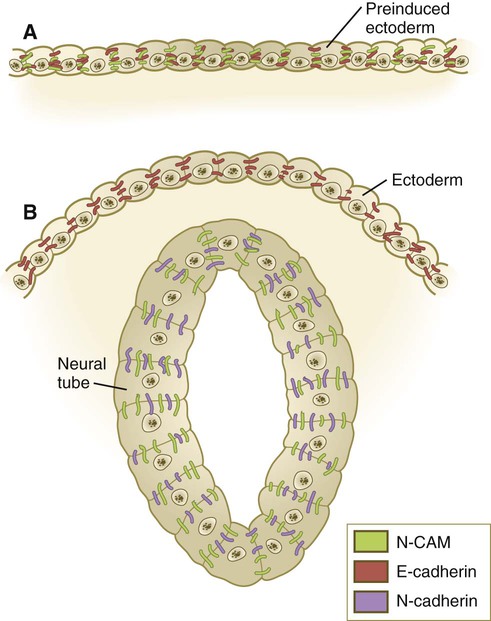
Preinduced ectoderm (A) after induction of the neural tube (B). CAM, cell adhesion molecule.
The third major family of CAMs, the integrins, attaches cells to components of basal laminae and the extracellular matrix (see Fig. 5.16). Integrins form heterodimers consisting of 1 of 16 α chains and 1 of 8 β chains. The matrix molecules to which they bind cells include fibronectin, laminin, and tenascin (see Fig. 12.3).
Summary
 Just before implantation, the inner cell mass becomes reorganized as an epithelium (epiblast), and a second layer (hypoblast) begins to form beneath it. Within the epiblast, the amniotic cavity forms by cavitation; outgrowing cells of the hypoblast give rise to the endodermal lining of the yolk sac. Extraembryonic mesoderm seems to form by an early transformation of parietal endodermal cells and cells migrating through the primitive streak.
Just before implantation, the inner cell mass becomes reorganized as an epithelium (epiblast), and a second layer (hypoblast) begins to form beneath it. Within the epiblast, the amniotic cavity forms by cavitation; outgrowing cells of the hypoblast give rise to the endodermal lining of the yolk sac. Extraembryonic mesoderm seems to form by an early transformation of parietal endodermal cells and cells migrating through the primitive streak.










*Hensen’s node is the commonly used designation for the primitive node in avian embryos, but this term is sometimes used in the mammalian embryological literature as well. This structure is the structural and functional equivalent of the dorsal lip of the blastopore in amphibians.
*The term specified (specification) is becoming increasingly used as a near synonym to determination in referring to the fixation of the future fate of a cell.
[/level-membership-for-basic-science-category][not-level-membership-for-basic-science-category]Chapter 5
Formation of Germ Layers and Early Derivatives
As it is implanting into the uterine wall, the embryo undergoes profound changes in its organization. Up to the time of implantation, the blastocyst consists of the inner cell mass, from which the body of the embryo proper arises, and the outer trophoblast, which represents the future tissue interface between the embryo and mother. Both components of the blastocyst serve as the precursors of other tissues that appear in subsequent stages of development. Chapter 3 discusses the way in which the cytotrophoblast gives rise to an outer syncytial layer, the syncytiotrophoblast, shortly before attaching to uterine tissue (see Fig. 3.18). Not long thereafter, the inner cell mass begins to give rise to other tissue derivatives as well. The subdivision of the inner cell mass ultimately results in an embryonic body that contains the three primary embryonic germ layers: the ectoderm (outer layer), mesoderm (middle layer), and endoderm (inner layer). The process by which the germ layers are formed through cell movements is called gastrulation.
Two-Germ-Layer Stage
Just before the embryo implants into the endometrium early in the second week, significant changes begin to occur in the inner cell mass and in the trophoblast. As the cells of the inner cell mass become rearranged into an epithelial configuration, sometimes referred to as the embryonic shield, a thin layer of cells appears ventral to the main cellular mass (see Fig. 3.18). The main upper layer of cells is known as the epiblast, and the lower layer is called the hypoblast, or primitive endoderm (Fig. 5.1).
(Note: The colors in the boxes are found in all illustrations involving the embryonic and extraembryonic germ layers.)
How the hypoblast forms in human embryos is not understood, but studies on mouse embryos have shown that as early as the 64-cell stage, some cells of the inner cell mass express the transcription factor nanog, whereas others express Gata 6. These cells are arranged in a salt and pepper pattern within the inner cell mass (Fig. 5.2A). The nanog-expressing cells represent the precursors of the epiblast, and those expressing Gata 6 will become the hypoblast. The basis for the differentiation of these two distinct precursor cell types is not completely understood, but according to the “time inside–time outside” hypothesis, those cells that enter the inner cell mass earliest are biased to express nanog, which perpetuates their pluripotency. Possibly because of the influence of fibroblast growth factor-4 (FGF-4), secreted by these first arrivals to the inner cell mass, later immigrants are then biased to express Gata 6. The Gata 6–expressing cells produce molecules that increase their adhesive properties, as well as their mobility, and they make their way to the lower surface of the inner cell mass to form a thin epithelium, the hypoblast. Those Gata 6 cells that fail to reach the surface of the inner cell mass undergo apoptosis (cell death). The nanog-expressing cells of the inner cell mass also assume an epithelial configuration as they form the epiblast. Between the epiblast and hypoblast a basal lamina forms.
The syncytiotrophoblast is not shown. A, Late blastocyst. Within the inner cell mass, blue nanog-expressing pre-epiblastic cells and yellow Gata 6–expressing prehypoblastic cells are mixed in a salt and pepper pattern. B, Beginning of implantation at 6 days. The hypoblast has formed and is beginning to spread beneath the cytotrophoblast as the parietal endoderm. C, Implanted blastocyst at
days. D, Implanted blastocyst at 8 days. E, Embryo at 9 days. F, Late second week.A small group of hypoblast cells that becomes translocated to the future anterior end of the embryo (called anterior visceral endoderm by mouse embryologists) has been shown to possess remarkable signaling powers. The cells first secrete the signaling molecules, lefty-1 and Cerberus-1 (Cer-1), which inhibit the activity of the signaling molecules, nodal and Wnt, in the overlying epiblast but allow nodal and Wnt-3 expression in the posterior epiblast (see Fig. 5.8A). (Nodal signaling from the posterior epiblast stimulates the initial formation of the anterior visceral endoderm.) This represents the first clear expression of anteroposterior polarity in the embryo. It also forms two signaling domains within the early embryo. The anterior visceral endoderm soon begins to induce much of the head and forebrain and inhibits the formation of posterior structures. In the posterior part of the epiblast, nodal signaling activity stimulates the formation of the primitive streak (see next section), which is the focal point for gastrulation and germ layer formation. After the hypoblast has become a well-defined layer, and the epiblast has taken on an epithelial configuration, the former inner cell mass is transformed into a bilaminar disk, with the epiblast on the dorsal surface and the hypoblast on the ventral surface.
The epiblast contains the cells that make up the embryo itself, but extraembryonic tissues also arise from this layer. The next layer to appear after the hypoblast is the amnion, a layer of extraembryonic ectoderm that ultimately encloses the entire embryo in a fluid-filled chamber called the amniotic cavity (see Chapter 7). Because of the paucity of specimens, the earliest stages in the formation of the human amnion and amniotic cavity are not completely understood. Studies on primate embryos indicate that a primordial amniotic cavity first arises by cavitation (formation of an internal space) within the pre-epithelial epiblast; it is covered by cells derived from the inner cell mass (see Fig. 5.2). According to some investigators, the roof of the amnion then opens, thus exposing the primordial amniotic cavity to the overlying cytotrophoblast. Soon thereafter (by about 8 days after fertilization), the original amniotic epithelium reforms a solid roof over the amniotic cavity.
While the early embryo is still sinking into the endometrium (about 9 days after fertilization), cells of the hypoblast begin to spread and line the inner surface of the cytotrophoblast with a continuous layer of extraembryonic endoderm called parietal endoderm (Fig. 5.3; see Fig. 5.2). When the endodermal spreading is completed, a vesicle called the primary yolk sac has taken shape (see Fig. 3.18C). At this point (about 10 days after fertilization), the embryo complex constitutes the bilaminar germ disk, which is located between the primary yolk sac on its ventral surface and the amniotic cavity on its dorsal surface (Fig. 5.4). Shortly after it forms, the primary yolk sac becomes constricted, forming a secondary yolk sac and leaving behind a remnant of the primary yolk sac (see Figs. 3.18D and 5.2F).
Starting at about 12 days after fertilization, another extraembryonic tissue, the extraembryonic mesoderm, begins to appear (see Fig. 5.2). The first extraembryonic mesodermal cells seem to arise from a transformation of parietal endodermal cells. These cells are later joined by extraembryonic mesodermal cells that have originated from the primitive streak. The extraembryonic mesoderm becomes the tissue that supports the epithelium of the amnion and yolk sac and the chorionic villi, which arise from the trophoblastic tissues (see Chapter 7). The support supplied by the extraembryonic mesoderm is not only mechanical, but also trophic because the mesoderm serves as the substrate through which the blood vessels supply oxygen and nutrients to the various epithelia.
Gastrulation and the Three Embryonic Germ Layers
At the end of the second week, the embryo consists of two flat layers of cells: the epiblast and the hypoblast. As the third week of pregnancy begins, the embryo enters the period of gastrulation, during which the three embryonic germ layers form from the epiblast (see Fig. 5.1). The morphology of human gastrulation follows the pattern seen in birds. Because of the large amount of yolk in birds’ eggs, the avian embryo forms the primary germ layers as three overlapping flat disks that rest on the yolk, similar to a stack of pancakes. Only later do the germ layers fold to form a cylindrical body. Although the mammalian egg is essentially devoid of yolk, the morphological conservatism of early development still constrains the human embryo to follow a pattern of gastrulation similar to that seen in reptiles and birds. Because of the scarcity of material, even the morphology of gastrulation in human embryos is not known in detail. Nevertheless, extrapolation from avian and mammalian gastrulation can provide a reasonable working model of human gastrulation.
Gastrulation begins with the formation of the primitive streak, a linear midline condensation of cells derived from the epiblast in the posterior region of the embryo through an induction by cells at the edge of the embryonic disk in that region (see Fig. 5.4). Members of the transforming growth factor-β (TGF-β) and Wnt families of signaling molecules have been identified as likely inducing agents. Initially triangular, the primitive streak soon becomes linear and elongates, largely through a combination of proliferation and migration, as well as internal cellular rearrangements, called convergent-extension movements. With the appearance of the primitive streak, the anteroposterior (craniocaudal) and right-left axes of the embryo can be readily identified (see Fig. 5.4).
The primitive streak is a region where cells of the epiblast converge in a well-defined spatial and temporal sequence. As cells of the epiblast reach the primitive streak, they change shape and pass through it on their way to forming new layers beneath (ventral to) the epiblast (Fig. 5.5C). Marking studies have shown that cells entering the primitive streak form distinct lineages as they leave. The most posterior cells both to enter and leave the streak as it is beginning to elongate form the extraembryonic mesoderm lining the trophoblast and yolk sac, as well as that forming the blood islands (see Fig. 6.19). Another wave of mesoderm, arising later and more anteriorly in the primitive streak, forms the paraxial, lateral plate, and cardiac mesoderm. A final wave, which enters and leaves the anteriormost end of the primitive streak, gives rise to midline axial structures (the notochord, the prechordal plate, and the primitive node itself) and also the embryonic endoderm. The composite results of such marking experiments are organized into fate maps, such as that illustrated in Figure 5.5A.
The endodermal precursor cells that pass through the anterior primitive streak largely displace the original hypoblast, but research has shown that some of the original hypoblastic cells become integrated into the newly forming embryonic endodermal layer. The displaced hypoblastic cells form extraembryonic endoderm. The movement of cells through the primitive streak results in the formation of a groove (primitive groove) along the midline of the primitive streak. At the anterior end of the primitive streak is a small but well-defined accumulation of cells, called primitive node, or Hensen’s node.* This structure is of great developmental significance because, in addition to being the major posterior signaling center of the embryo (Box 5.1), it is the area through which cells migrate in a stream toward the anterior end of the embryo. These cells, called mesendoderm, soon segregate into a rodlike mesodermal notochord and the endodermal dorsal wall of the forming gut. Anterior to the notochord is a group of mesodermal cells called the prechordal plate (see Fig. 5.5A and B). (The important functions of the notochord and prechordal plate are discussed on p. 80.)
Box 5.1 Molecular Aspects of Gastrulation
For this aspect of early development, we must rely almost entirely on studies of mouse embryos. The original symmetry of the embryo is broken by the displacement of the future anterior visceral endoderm to the anterior side of the embryonic disk. This is a function of proliferation and later migration of the cells that will constitute the anterior visceral endoderm. Migration of these cells (and the resulting establishment of the anteroposterior axis) depends on the activation of the Wnt antagonist Dkk 1 (Dickkopf 1) in the future anterior part of the embryo. This confines Wnt activity to the future posterior part of the embryo, where it induces the expression of the signaling molecule Nodal (Fig. 5.8A). When the anterior visceral endoderm has become stabilized in the anterior part of the embryonic disk, it produces the Nodal inhibitors lefty-1 and Cer-1, which confine Nodal activity to the posterior end of the embryo where, responding to extraembryonic Wnt signals, it establishes a posterior signaling center, which induces the formation of the primitive streak, the definitive endoderm, and the mesoderm. In the chick embryo, the ectopic application of two other signaling molecules, chordin and Vg1, induces the formation of an ectopic primitive streak.
A, Preprimitive streak (sagittal section). B, Early formation of the primitive streak. C, Gastrulation (period of germ layer formation). D, Late gastrulation and neural induction. The molecules in red are signaling molecules, and the molecules in blue are transcription factors. Names of specific molecules (bold) are placed by the structures in which they are expressed.
As the primitive streak elongates, migrating cells of the epiblast join the tip of the streak, and a dynamic mass of cells, called the primitive node, becomes evident at the tip of the primitive streak. Cells of the node express many genes, including three classic molecular markers of the organizer region in many vertebrates—chordin, goosecoid, and hepatic nuclear factor-3β (now called Foxa-2). Not only is the winged helix transcription factor, Foxa-2, important for the formation of the node itself, but also it is vital for the establishment of midline structures cranial to the node. Foxa-2 is required for the initiation of notochord function. In its absence, the notochord and the floor plate of the neural tube (see Chapter 11) fail to form. In contrast, endoderm, the primitive streak, and intermediate mesoderm do develop. Goosecoid, a homeodomain transcription factor, is prominently expressed in the organizer region of all vertebrates studied. Goosecoid activates chordin, noggin, and other genes of the organizer region. If ectopically expressed, it stimulates the formation of a secondary body axis. Chordin and noggin, signaling molecules associated with the node, are involved with neural induction, and expression of nodal on the left side of the embryo is a key element in the setting of left-right asymmetry.
Two genes, T and nodal, play prominent roles in the function of the primitive streak and posterior mesoderm formation. Expression of the T gene seems to be activated by products of the Foxa-2 and goosecoid genes. In T mutants (brachyury), the notochord begins to form through the activity of Foxa-2, but it fails to complete development. Studies on T mutants have shown that activity of the brachyury gene is necessary for normal movements of future mesodermal cells through the primitive streak during gastrulation. In brachyury (short tail) mutant mice, mesodermal cells pile up at a poorly formed primitive streak, and the embryos show defective elongation of the body axis (including a short tail) posterior to the forelimbs. T gene mutants may be responsible for certain gross caudal body defects in humans. Nodal, a member of the transforming growth factor-β (TGF-β) family of growth factor genes (see Table 4.1), is expressed throughout the posterior epiblast before gastrulation, but its activity is concentrated at the primitive node during gastrulation. Similar to the brachyury gene, the effects of nodal are strongly seen in the caudal region of the embryo. In the null mutant of nodal, the primitive streak fails to form, and the embryo is deficient in mesoderm. Similarly, mutants of cripto, an early-acting member of the epidermal growth factor family and an essential cofactor in the nodal signaling pathway, produce a trunkless phenotype.
As cells pass through the primitive streak, a region of Hox gene expression begins to form around the streak. The pattern of Hox gene expression in the future trunk and posterior part of the embryo is based on signaling by three molecules—retinoic acid, Wnt, and FGF—that act on the transcription factor Cdx (the mammalian equivalent of caudal in Drosophila) in the area of the regressing primitive streak just behind the last-forming somites. Cdx acts on the Hox genes, which impose unique characteristics to the segmental structures that form along the anteroposterior axis of the embryo (Fig. 5.8D).
Anterior Visceral Endoderm (Hypoblast)
In mammals, even before mesodermal cells begin migrating through the primitive node, the anterior hypoblast (called the anterior visceral endoderm by mouse embryologists) expresses genes characteristic of the prechordal plate and initiates head formation. The anterior visceral endoderm itself is subdivided into an anterior part, which serves as a signaling center for early heart formation (see p. 104
[/not-level-membership-for-basic-science-category]
