Chapter 46 Dyslexia
Developmental dyslexia (or specific reading disability) is defined as an unexpected difficulty in accuracy or fluency of reading for an individual’s chronological age, intelligence, level of education, or professional status. Dyslexia is, at its core, a problem with phonological processing: that is, getting to the elemental sounds of spoken language, affecting both spoken and written language. As a consequence, individuals who are dyslexic require accommodations for their lack of reading and/or oral fluency. As dyslexic children progress in school, given good instruction, reading accuracy often improves; however, lack of fluency persists and remains a lifelong problem. Dyslexia is the most common and most comprehensively studied of the learning disabilities, affecting 80 percent of all individuals identified as learning-disabled. Historically, dyslexia in adults was first observed in the latter half of the 19th century, and developmental dyslexia in children was first reported in 1896 [Morgan, 1896]. Although the diagnosis and implications of dyslexia were often uncertain in the past, advances in our knowledge of the epidemiologic, neurobiologic, and cognitive influences on the disorder allow it to be approached within the framework of a traditional medical model. This chapter reviews these advances and their implications for the approach to children and adults with dyslexia.
Definition
Perhaps the most consistent and enduring core of the definition is the concept of dyslexia as an unexpected difficulty in reading. “Unexpected” refers to the presence of a reading difficulty in a child (or adult) who appears to have all of the factors (intelligence, motivation, exposure to reasonable reading instruction) present to be a good reader but who continues to struggle [Shaywitz, 1998]. Recent evidence provides empiric support for defining dyslexia as an unexpected difficulty in reading. Using data from the Connecticut Longitudinal Study, we [Ferrer et al., 2010] demonstrated that, in typical readers, reading and IQ development are dynamically linked over time. Not only do reading and IQ track together over time, they also influence one another. Such mutual interrelationships are not perceptible in dyslexic readers, suggesting that reading and cognition develop more independently in these individuals (Figure 46-1). These findings provide the first empirical demonstration of a coupling between cognition and reading in typical readers, confirming the general public perception that if you are a good reader, you are likely to be very intelligent and, conversely, if you struggle to read, you may be less intelligent. These new data demonstrating a developmental uncoupling between cognition and reading in dyslexic readers indicate that dyslexia is a special case that violates the assumption that reading and IQ are always linked, and confirm that, in dyslexia, one can be highly intelligent and still struggle to read.
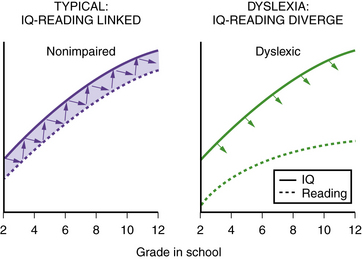
Fig. 46-1 Uncoupling of reading and IQ over time: empirical evidence for a definition of dyslexia.
(Data adapted from Ferrer E et al. Uncoupling of reading and IQ over time: empirical evidence for a definition of dyslexia. Psychological Science; 2010.)
More challenging has been the question of how to operationalize the unexpected nature of dyslexia. Thus, using differing methods and criteria, definitions have attempted to capture the “unexpected” nature of dyslexia by requiring a discrepancy of a certain degree between a child’s measured IQ and his reading achievement. For example, schools have typically relied on criteria based on an absolute discrepancy, most commonly one or one-and-one-half standard deviations between standard scores on IQ and reading tests. We want to emphasize that the difficulty has been not with the concept of a disparity, but rather with the real-life practical effect of implementing this model in a primary school setting. For example, children who were clearly struggling as early as kindergarten or first grade had to wait, often until third grade or later, until their failure in reading was of such a magnitude that they met discrepancy requirements. Attempts to clarify the criteria by meta-analyses comparing discrepant to simply low-achieving poor readers (defined on the basis of a reading score below a certain cut point, e.g., below a standard score of 90) find overlap between the two groups on reading-related constructs but differences on IQ-related measures [Stuebing et al., 2002]. In addition, studies examining growth curve models for low-achieving and discrepant readers indicate comparable reading plateaus (level of reading achievement) reached by the two groups but with the IQ-discrepant readers showing the lowest achievement level at any IQ level during the school years [Francis et al., 1996]. Not only do poor readers identified by either discrepancy or low-achievement criteria resemble one another on measures of reading and growth rates of reading, but each group also differs along multiple dimensions from groups of typically achieving boys and girls [Lyon et al., 2001].
These findings have strong educational implications. It is not valid to deny the education services available for disabled or at-risk readers either to low-achieving, nondiscrepant children, or to those children who are not low-achieving but who, at the same time, are reading below a level expected for their ability. The observed similarity of the discrepant and low-achieving groups in reading-related constructs argues for identification approaches that include both low-achieving children and those struggling readers who are discrepant but who do not satisfy an arbitrary cut point for designation as low-achieving. Seventy-five percent of children identified by discrepancy criteria also meet low-achievement criteria in reading; the remaining 25 percent who meet only discrepancy criteria may fail to be identified and yet still be struggling to read [Shaywitz et al., 1992].
Epidemiology
Epidemiologic data indicate that, like hypertension and obesity, dyslexia fits a dimensional model. Within the population, reading ability and reading disability occur along a continuum, with reading disability representing the lower tail of a normal distribution of reading ability [Shaywitz et al., 1992]. Dyslexia is perhaps the most common neurobehavioral disorder affecting children, with prevalence rates ranging from 5 to 17.5 percent [Interagency Committee on Learning Disabilities, 1987; Shaywitz, 2003]. Longitudinal studies, prospective [Francis et al., 1996; Shaywitz et al., 1999] and retrospective [Bruck, 1992; Felton et al., 1990], indicate that dyslexia is a persistent, chronic condition; it does not represent a transient developmental lag (Figure 46-2). Over time, poor readers and good readers tend to maintain their relative positions along the spectrum of reading ability [Francis et al., 1996; Shaywitz et al., 1995].
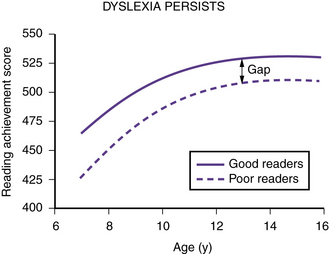
Fig. 46-2 Trajectory of reading skills over time in nonimpaired and dyslexic readers.
Numbers on the ordinate are Rasch scores (W scores) from the Woodcock–Johnson reading test [Woodcock and Johnson, 1989], and numbers on the abscissa are ages in years. Dyslexic and nonimpaired readers improve their reading scores as they get older, but the gap between the dyslexic and nonimpaired readers remains. Dyslexia is a deficit, not a developmental lag.
(Adapted from Shaywitz S. Overcoming dyslexia: A new and complete science-based program for reading problems at any level. New York: Alfred A. Knopf, 2003. Copyright 2003 by S. Shaywitz. Adapted with permission.)
Etiology
Dyslexia is both familial and heritable [Pennington and Gilger, 1996]. Family history is one of the most important risk factors, with 23–65 percent of children who have a parent with dyslexia reported to have the disorder [Scarborough, 1990]. A rate among siblings of affected persons of approximately 40 percent and among parents of 27–49 percent [Pennington and Gilger, 1996] provides opportunities for early identification of affected siblings and often for delayed but helpful identification of affected adults. Given that dyslexia is familial and heritable, initial hopes that dyslexia would be explained by one or just a few genes have been disappointing. Thus, along with a great many common diseases, genome-wide association studies (GWAS) in dyslexia have so far identified genetic variants that account for only a very small percentage of the risk – less than 1 percent [Meaburn et al., 2008]. Current evidence suggests “that common diseases involve thousands of genes and proteins interacting on complex pathways” [Duncan, 2009], and that, similar to experience with other complex disorders (heart disease, diabetes), it is unlikely that a single gene or even a few genes will identify people with dyslexia. Rather, dyslexia is best explained by multiple genes, each contributing a small amount of the variance. Thus, current evidence suggests that the etiology of dyslexia is best conceptualized within a multifactorial model, with multiple genetic and environmental risk and protective factors leading to dyslexia.
Cognitive Influences
Among investigators in the field, there is a strong consensus supporting the phonologic theory. This theory recognizes that speech is natural and inherent, but that reading is acquired and must be taught. To read, the beginning reader must connect the letters and letter strings (i.e., the orthography) to something that already has inherent meaning – the sounds of spoken language. In the process, a child has to develop the insight that spoken words can be pulled apart into the elemental particles of speech (i.e., phonemes) and that the letters in a written word represent these sounds [Shaywitz, 2003]; such awareness is largely deficient in dyslexic children and adults [Liberman and Shankweiler, 1991; Shankweiler et al., 1979; Shaywitz, 2003]. Results from large and well-studied populations with reading disability confirm that, in young school-age children [Stanovich and Siegel, 1994] and in adolescents [Shaywitz et al., 1999], a deficit in phonology represents the most robust and specific correlate of reading disability [Morris et al., 1998; Ramus et al., 2003]. Such findings form the basis for the most successful and evidence-based interventions designed to improve reading [Report, 2000].
Implications of the Phonologic Model of Dyslexia
Reading comprises two main processes: decoding and comprehension [Gough and Tunmer, 1986]. In dyslexia, a deficit at the level of the phonologic module impairs the ability to segment the spoken word into its underlying phonologic elements. As a result, the reader experiences difficulty, initially in spoken language and then in written language. The phonologic deficit is domain-specific; that is, it is independent of other, nonphonologic abilities. In particular, the higher-order cognitive and linguistic functions involved in comprehension, such as general intelligence and reasoning, vocabulary [Share and Stanovich, 1995], and syntax [Shankweiler et al., 1995], are generally intact. This pattern – a deficit in phonologic analysis contrasted with intact higher-order cognitive abilities – offers an explanation for the paradox of otherwise intelligent, often gifted people who experience great difficulty in reading [Ferrer et al., 2010; Shaywitz, 1996, 2003].
Neurobiologic Studies
Neural Systems for Reading
Our understanding of the neural systems for reading emerged more than a century ago, with descriptions of adults who (usually due to a stroke) suddenly lost their ability to read, a condition termed acquired alexia. These postmortem studies, pioneered by Dejerine as early as 1891, suggested that a portion of the left posterior brain region (which includes the angular gyrus and supramarginal gyrus in the inferior parietal lobule and the posterior aspect of the superior temporal gyrus) is critical for reading [Dejerine, 1891]. Another left posterior brain region, one more ventral in the occipito-temporal area, was also described by Dejerine [Dejerine, 1892] as critical in reading. Within the last two decades, the development of functional brain imaging, particularly functional magnetic resonance imaging (fMRI), has provided the most consistent and replicable data on the location of the neural systems for reading and how they differ in dyslexic readers. fMRI is noninvasive and safe, and can be used repeatedly, properties which make it ideal for studying people, especially children. The signal used to construct MRI images derives from the determination of the blood oxygen level-dependent (BOLD) response; the increase in BOLD signal in regions that are activated by a stimulus or task results from the combined effects of increases in the tissue blood flow, volume and oxygenation, and in cognitive tasks the changes are typically in the order of 1–5 percent. Details of fMRI are reviewed in Anderson and Gore [1997], Frackowiak et al. [2004], and Jezzard et al. [2001]. To date, fMRI in dyslexic individuals can be carried out reliably only at a group level. The technology for determining brain activation at an individual subject level remains a work in progress.
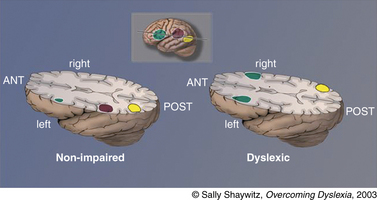
Reflecting the language basis for reading and dyslexia, three neural systems critical in reading and dyslexia are localized in the left hemisphere (Figure 46-3): two left hemisphere posterior systems, one around the occipito-temporal region and another in the left occipito-temporal region, and an anterior system around the inferior frontal gyrus (Broca’s area) [Brambati et al., 2006; Helenius et al., 1999; Kronbichler et al., 2006; Nakamura et al., 2006; Paulesu et al., 2001; Shaywitz et al., 2002; Shaywitz et al., 2003; Shaywitz et al., 1998].
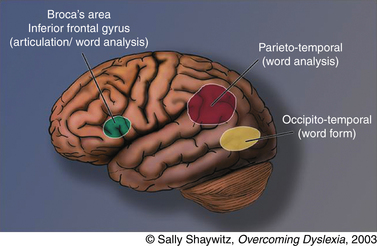
Fig. 46-3 Neural systems for reading.
(Adapted from Shaywitz S. Overcoming dyslexia: A new and complete science-based program for reading problems at any level. New York: Alfred A. Knopf, 2003. Copyright 2003 by S. Shaywitz. Adapted with permission.)
Many brain imaging studies in children and adults with developmental dyslexia (see below) have documented the importance of the left occipito-temporal system in reading, and its properties involving word analysis, operating on individual units of words (e.g., phonemes). In our figure we refer to the occipito-temporal system, which encompasses portions of the supramarginal gyrus in the inferior parietal lobule, portions of the posterior aspect of the superior temporal gyrus, and in some studies, may even encompass portions of the angular gyrus in the parietal lobe. The second posterior reading system is localized in the left occipito-temporal area, which Cohen and Dehaene have termed the visual word-form area (VWFA) [Cohen et al., 2000; Dehaene et al., 2005; Vinckier et al., 2007]. Just how the VWFA functions to integrate phonology (sounds) and orthography (print) is as yet unknown, though some have suggested that visual familiarity, phonological processing, and semantic processing all make significant but different contributions to activation of the word-form region [Cohen et al., 2004; Henry et al., 2005; Johnson and Rayner, 2007; Xue et al., 2006]. Still another reading-related neural circuit involves an anterior system in the left inferior frontal gyrus (Broca’s area), a system that has long been associated with articulation and also serves an important function in word analysis [Fiez and Peterson, 1998; Frackowiak et al., 2004].
The Reading Systems in Dyslexia in Children and Adults
Converging evidence from many laboratories around the world has demonstrated what has been termed “a neural signature for dyslexia”: that is, inefficient functioning of left posterior reading systems during reading real words and pseudowords, and often what has been considered as compensatory overactivation in other parts of the reading system. This evidence from functional brain imaging has, for the first time, made visible what previously was a hidden disability (Figure 46-4).
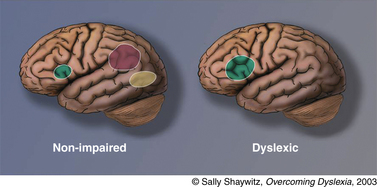
Fig. 46-4 Neural signature for dyslexia.
A neural signature for dyslexia is illustrated in this schematic view of left hemisphere brain systems in (left) nonimpaired and (right) dyslexic readers. In typical readers, the three systems provided in Figure 46-3 are shown. In dyslexic readers, the anterior system is slightly overactivated compared with systems of typical readers; in contrast, the two posterior systems are underactivated. This pattern of underactivation in left posterior reading systems is referred to as the neural signature for dyslexia.
(Adapted from Shaywitz S. Overcoming dyslexia: A new and complete science-based program for reading problems at any level. New York: Alfred A. Knopf, 2003. Copyright 2003 by S. Shaywitz. Adapted with permission.)
For example, in a study from our own research group, we [Shaywitz et al., 2002] used fMRI to study 144 dyslexic and nonimpaired boys and girls as they read pseudowords and real words. Our results indicated significantly greater activation in typical readers than in dyslexic readers during phonologic analysis in the posterior reading systems. Our data converge with reports from many investigators using functional brain imaging in dyslexia that show a failure of left hemisphere posterior brain systems to function properly during reading (reviewed in Richlan et al. [2009]; Shaywitz and Shaywitz [2005]). Recent studies report similar findings in German [Kronbichler et al., 2006] and Italian [Brambati et al., 2006] dyslexic readers.
Development of Reading Systems in Dyslexia
While converging evidence points to three important neural systems for reading, few studies have examined age-related changes in these systems in typical readers or in dyslexic children. We [Shaywitz et al., 2007] used fMRI to study age-related changes in reading in a cross-sectional study of 232 dyslexic and nonimpaired boys and girls as they read pseudowords. Findings indicated that the neural systems for reading that develop with age in typical readers differ from those that develop in dyslexic readers. Specifically, a system for reading that develops with age in dyslexic readers involves a more posterior and medial system, in contrast to a more anterior and lateral system within the left occipito-temporal area in typical readers. Interestingly, this difference in activation patterns between the two groups of readers has parallels to reported brain activation differences observed during reading of two Japanese writing systems: Kana and Kanji. Left anterior lateral occipito-temporal activation, similar to that seen in typical readers, occurred during reading Kana [Nakamura et al., 2005]. Kana script employs symbols that are linked to the sound or phonologic element (comparable to English and other alphabetic scripts). In Kana and in alphabetic scripts, children initially learn to read words by learning how letters and sounds are linked and then, over time, these linkages are integrated and permanently instantiated as a word form.
In contrast, posterior medial occipito-temporal activation, comparable to that observed in dyslexic readers, was noted during reading of Kanji script [Nakamura et al., 2005]. Consideration of the mechanisms used for reading Kanji compared to Kana provides insights into potentially different mechanisms that develop with age in dyslexic contrasted to typical readers. Kanji script uses ideographs where each character must be memorized, suggesting that the left posterior medial occipito-temporal system functions as a memory-based system. It is reasonable to suppose that, as dyslexic children mature, this posterior medial system supports memorization rather than the progressive sound–symbol linkages observed in typical readers. There is also evidence that, as they mature, dyslexic readers are not able to make good use of sound–symbol linkages and, instead, come to rely on memorized words. For example, phonologic deficits continue to characterize struggling readers even as they enter adolescence and adult life [Bruck, 1992; Shaywitz et al., 1999], and persistently poor adult readers read words by memorization so that they are able to read familiar words but have difficulty reading unfamiliar words [Shaywitz et al., 2003].
Furthermore, functional brain imaging has provided, for the first time, evidence demonstrating that dyslexic readers require the accommodation of extra time on high-stakes standardized tests. Thus, although dyslexic readers exhibit an inefficiency of functioning in the left occipito-temporal word-form area, they appear to develop ancillary systems involving areas around the inferior frontal gyrus in both hemispheres, as well as the right hemisphere homolog of the left occipito-temporal word-form area [Shaywitz et al., 2002]. While these ancillary systems allow the dyslexic reader to read accurately, dyslexic readers continue to read nonfluently. Inefficient functioning in this system for skilled reading has very important practical implications for the dyslexic reader – it provides the neurobiological evidence for the biologic necessity for the accommodation of additional time on high-stakes tests (Figure 46-5).
Diagnosis
In the preschool child, a history of language delay or of not attending to the sounds of words (e.g., trouble learning nursery rhymes or playing rhyming games with words, confusing words that sound alike, mispronouncing words) and trouble learning to recognize the letters of the alphabet, along with a positive family history, represent important risk factors for dyslexia. In the school-aged child, presenting complaints most commonly center on school performance – “she’s not doing well in school” – and often parents (and teachers) do not appreciate that the reason is a reading difficulty. A typical picture is that of a child who may have had a delay in speaking, does not learn letters by kindergarten, has not begun to learn to read by first grade, and has difficulty consistently sounding out words. The child progressively falls behind, with teachers and parents puzzled about why such an intelligent child may have difficulty learning to read. The reading difficulty is unexpected with respect to the child’s ability, age, or grade. Even after acquiring decoding skills, the child typically remains a slow reader. Bright dyslexic children may laboriously learn how to read words accurately, but they do not become fluent readers because they do not recognize words rapidly and automatically. Dysgraphia and spelling difficulties are often present and accompanied by laborious note-taking. Self-esteem is frequently affected, particularly if the disorder has gone undetected for a long period (Box 46-1) [Shaywitz, 2003].

Dyslexia is conceptualized as fitting what is referred to as a “sea of strengths” model, with strengths in higher cognitive functions surrounding the weakness in phonological awareness and fluent reading [Shaywitz, 2003]. Thus, the same individual who struggles to retrieve words and reads slowly with effort may have significant strengths in conceptual abilities, problem-solving, verbal reasoning, the ability to grasp the big picture, and what has been referred to as creative or out-of-the-box thinking. Indeed, it is just these higher-level strengths that make it imperative for individuals who have dyslexia to receive the accommodations (e.g., extra time) they require in order to demonstrate their abilities.
Assessment of Pre-reading and Reading
Even before the time at which a child is expected to read, a child’s readiness to read or at-risk status for specific reading disability may be assessed by measurement of the skills, especially phonologic, related to reading success. After a predictable developmental pathway, children’s phonologic abilities can be evaluated beginning at about age 4 years. Such tests mainly are centered on a child’s ability to focus on phonemes, the basic particles of spoken language. Initial tests typically ask what word rhymes with another or what spoken word begins (or ends) with the same sound as another. At more advanced levels, tests ask children to pronounce a spoken word after a sound is removed, such as “can you say steak without the t sound?” (sake) or “can you count the number of sounds you hear in man?” (three sounds). In general, as a child develops, he or she gains the ability to notice and to manipulate smaller and smaller parts of spoken words. Tests of phonologic capabilities and reading readiness are becoming increasingly available; one such test is the Comprehensive Test of Phonological Processing in Reading (CTOPP, PRO-ED), which is nationally standardized for age 5 through adult years. In addition to phonology, knowledge of letter names and sounds is the strongest predictor of a child’s readiness to read. An appropriate battery of tests for the early recognition of reading problems includes tests of phonology, letter names and sounds, vocabulary, print conventions, and listening comprehension. Tests of reading are also useful because they allow comparison of a child’s reading skills with his peers at a time when he should be beginning to read [Shaywitz, 2003]. The importance of such early assessments is that they can identify at-risk children early on, so that these boys and girls can be provided with the effective, evidence-based reading interventions now available.
For informal screening by primary care physicians in an office setting, it is recommended that the physician listen to the child read aloud from his own grade-level reader. Keeping a set of graded readers available in the office serves the purpose and does not require children to bring their own schoolbooks. Oral reading is a very sensitive measure of reading accuracy and reading fluency. A set of read-aloud paragraphs appropriate for children in the primary grades can be found in Appendix 46-1.
Three reading samples are provided [Hiebert, 2003]. They can be photocopied and given to young patients as reading tests for informal screening of dyslexia during an office visit. The superscript numbers indicate word counts.
(From Hiebert E. QuickReads. Parsippany, NJ: Modern Curriculum Press, 2003. © 2003 by Pearson Education, Inc. Used by permission.)
Outcome: Phonologic Deficit in Adolescence and Adult Life
Lack of fluency and deficits in phonologic coding continue to characterize dyslexic readers, even in adolescence; performance on phonologic processing measures contributes most to discriminating between dyslexic and typical adolescent readers and typical and superior adolescent readers as well [Shaywitz et al., 1999]. Dyslexia does not spontaneously remit, nor do these children demonstrate a lag mechanism for “catching up” in the development of reading skills. However, many dyslexic readers can become quite proficient in reading a finite domain of words that recur in their area of special interest, usually words that are important for their careers. For example, an individual who is dyslexic in childhood may in adult life become interested in molecular biology and learn to decode words that form a minivocabulary important in molecular biology. Such an individual, although able to decode words in this domain, still exhibits evidence of his early reading problems when reading unfamiliar words, which then are read accurately but not fluently and automatically [Ben-Dror et al., 1991; Bruck, 1994; Lefly and Pennington, 1991; Shaywitz et al., 1999]. Because they are able to read words accurately (albeit very slowly), dyslexic adolescents and young adults may mistakenly be assumed to have “outgrown” their dyslexia. Data from studies of children with dyslexia who have been followed prospectively support the notion that, in adolescents, the rate of reading and the facility with spelling may be most useful clinically in differentiating average from poor readers in students in secondary school, college, and even graduate school. These older dyslexic students may be similar to their unimpaired peers on untimed measures of word recognition but continue to suffer from the phonologic deficit that makes reading less automatic, more effortful, and slow.
Treatment
As in other areas of medicine, knowledge is being accrued to inform the practice of evidence-based education. Based on a prior consensus report from the National Research Council [Snow et al., 1998] and on the results of its own analysis, the National Reading Panel [Report, 2000] reported that five critical elements were necessary to teach reading effectively: phonemic awareness (i.e., ability to focus on and manipulate phonemes – speech sounds – in spoken syllables and words); phonics (i.e., understanding how letters are linked to sounds to form letter–sound correspondences and spelling patterns); fluency (i.e., ability to read accurately, rapidly, and with good intonation); vocabulary; and comprehension strategies. The National Reading Panel emphasized that these elements must be taught systematically and explicitly, rather than in a more casual, fragmented, or implicit manner. Such systematic phonics instruction is more effective than “whole-word” instruction, which teaches little or no phonics or teaches phonics haphazardly or using a “by-the-way” approach.
Large-scale studies have focused on younger children, and there are few or no data available on the effect of these training programs on older children. The data on younger children are extremely encouraging, indicating that using evidence-based methods can remediate and even prevent reading difficulties in primary school-aged children [Foorman et al., 2003; Shaywitz, 2003; Torgesen et al., 1999].
Attentional Mechanisms in Reading and Dyslexia
For almost two decades, the central dogma in reading research has been that the generation of the phonological code from print is modular: that is, automatic, not attention-demanding, and not requiring any other cognitive process. Recent findings now present a competing view, suggesting that attentional mechanisms play a critical role in reading and that disruption of these attentional mechanisms plays a causal role in reading difficulties. These data, reviewed by us previously [Shaywitz and Shaywitz, 2008], provide one of the most exciting new developments on the horizon: the potential use of pharmacologic agents that influence not only attention, but reading as well. Randomized clinical trials of such agents are currently in progress and may lead to a role for pharmacotherapy in reading as an adjunct to the reading interventions discussed above.
Accommodations
An essential component of the management of dyslexia in students in secondary school, college, and graduate school incorporates the provision of accommodations. High-school and college students with a history of childhood dyslexia often present a paradoxical picture; they are similar to their unimpaired peers on measures of word recognition and comprehension, but they continue to suffer from the phonologic deficit that makes reading less automatic, more effortful, and slow. Neurobiologic data provide strong evidence for the necessity of extra time for readers with dyslexia (see Figure 46-5). Functional MRI data demonstrate that, in the word-form area, the region supporting rapid reading functions inefficiently. Readers compensate by developing anterior systems bilaterally and the right homolog of the left word-form area. Such compensation allows for accurate reading, but it does not support fluent or rapid reading [Shaywitz et al., 2002]. For these readers with dyslexia, the provision of extra time is an essential accommodation; it allows them the time to decode each word and to apply their unimpaired higher-order cognitive and linguistic skills to the surrounding context to get at the meaning of words that they cannot entirely or rapidly decode. While readers who are dyslexic improve greatly with additional time, providing additional time to nondyslexic readers results in very minimal to lack of improvement in scores. Although providing extra time for reading is by far the most common accommodation for people with dyslexia, other helpful accommodations include allowing the use of laptop computers with spelling checkers, and access to recorded books. (Recordings read by actual people are available from Learning Ally formerly Recording for the Blind and Dyslexic, 800-221-4792 or audible.com, while digitized materials are read by artificial voice on Kindle or Kurzweil readers.) Other helpful accommodations include providing access to syllabi and lecture notes, tutors to “talk through” and review the content of reading material, alternatives to multiple-choice tests (e.g., reports or projects), waivers of high-stakes oral exams and a separate, quiet room for taking tests [Shaywitz, 2003]. With such accommodations, many students with dyslexia are successfully completing studies in a range of disciplines, including medicine.
Dyslexia and the Sea of Strengths
Dyslexia is conceptualized as a “sea of strengths” in higher cognitive functions surrounding the weakness in phonological awareness and fluent reading [Shaywitz, 2003]. Thus, the same individual who struggles to retrieve words and reads slowly with effort may have great strengths in conceptual abilities, problem-solving, verbal reasoning, the ability to grasp the big picture, and what has been referred to as creative or out-of-the-box thinking. Indeed, it is just these higher-level strengths that make it imperative for individuals who have dyslexia to receive the accommodations (e.g., extra time) they require in order to demonstrate their abilities. It is important to appreciate that phonologic difficulties in dyslexia are independent of intelligence. Many highly intelligent boys and girls have reading problems that are often overlooked and even ascribed to “lack of motivation.” In counseling their patients who are dyslexic, physicians should bear in mind that many outstanding writers, lawyers, physicians, and scientists, including Nobel laureates, such as Niels Bohr in physics (1925) and Baruj Benacerraf (1984) and Carol Greider (2009) in physiology or medicine, are dyslexic. More information regarding the relationship between dyslexia and creativity may be found on the website of the Yale Center for Dyslexia and Creativity, www.dyslexia.Yale.edu.
References
 The complete list of references for this chapter is available online at www.expertconsult.com.
The complete list of references for this chapter is available online at www.expertconsult.com.
Anderson A., Gore J.. The physical basis of neuroimaging techniques. Lewis M., Peterson B., editors. Child and adolescent psychiatric clinics of North America. vol. 6. Philadelphia, PA: W. B. Saunders Co; 1997:213-264.
Ben-Dror I., Pollatsek A., Scarpati A. Word identification in isolation and in context by college dyslexic students. Brain Lang. 1991;40:471-490.
Brambati S., Termine C., Ruffino M., et al. Neuropsychological deficits and neural dysfunction in familial dyslexia. Brain Res. 2006;1113(1):174-185. [doi: 10.1016/j.brainres.2006.06.099]
Bruck M. Persistence of dyslexics’ phonological awareness deficits. Dev Psychol. 1992;28(5):874-886.
Bruck M. Outcomes of adults with childhood histories of dyslexia. In: Hulme C., Joshi R., editors. Cognitive and linguistic bases of reading, writing, and spelling. Mahwah, NJ: Lawrence Erlbaum Associates; 1994:179-200.
Cohen L., Dehaene S., Naccache L., et al. The visual word form area: spatial and temporal characterization of an initial stage of reading in normal subjects and posterior split-brain patients. Brain. 2000;123(Pt 2):291-307.
Cohen L., Jobert A., Le Bihan D., et al. Distinct unimodal and multimodal regions for word processing in the left temporal cortex. Neuroimage. 2004;23(4):1256-1270.
Dehaene S., Cohen L., Sigman M., et al. The neural code for written words: a proposal. Trends Cogn Sci. 2005;9(7):335-341.
Dejerine J. Sur un cas de cécité verbale avec agraphie, suivi d’autopsie. C R Société du Biologie. 1891;43:197-201.
Dejerine J. Contribution a l’etude anatomo-pathologique et clinique des differentes varietes de cecite verbale. Memoires de la Societe de Biologie. 1892;4:61-90.
Duncan D. Scientist at Work: Eric Schadt, Enlisting Computers to Unravel the True Complexity of Disease. New York Times. 2009. August 24, 2009
Felton R.H., Naylor C.E., Wood F.B. Neuropsychological profile of adult dyslexics. Brain Lang. 1990;39:485-497.
Ferrer E., Shaywitz B., Holahan J., et al. Uncoupling of Reading and IQ Over Time: Empirical Evidence for a Definition of Dyslexia. Psychol Sci. 2010;21(1):93-101.
Fiez J.A., Peterson S.E. Neuroimaging studies of word reading. Proc Natl Acad Sci USA. 1998;95(3):914-921.
Foorman B.R., Brier J.I., Fletcher J.M. Interventions aimed at improving reading success: an evidence-based approach. Dev Neuropsychol. 2003;24:613-639.
Frackowiak R., Friston K., Frith C., et al. Human Brain Function, ed 2. San Diego: Academic Press, Elsevier Science; 2004.
Francis D., Shaywitz S., Stuebing K., et al. Developmental lag versus deficit models of reading disability: A longitudinal, individual growth curves analysis. J Educ Psychol. 1996;88(1):3-17.
Gough P.B., Tunmer W.E. Decoding, reading, and reading disability. Remedial and Special Education. 1986;7:6-10.
Helenius P., Tarkiainen A., Cornelissen P., et al. Dissociation of normal feature analysis and deficient processing of letter-strings in dyslexic adults. Cerebral Cortex. 1999;4:476-483.
Henry C., Gaillard R., Volle E., et al. Brain activations during letter-by-letter reading: a follow-up study. Neuropsychologia. 2005;43(14):1983-1989.
Hiebert E. QuickReads. Parsippany, NJ: Modern Curriculum Press; 2003.
Interagency Committee on Learning Disabilities. Learning disabilities: A report to the U.S. Congress. Washington, DC: U.S. Government Printing Office; 1987.
Jezzard P., Matthews P., Smith S. Functional MRI, An Introduction to Methods. Oxford: Oxford University Press; 2001.
Johnson R., Rayner K. Top-down and bottom-up effects in pure alexia: evidence from eye movements. Neuropsychologia. 2007;45(10):2246-2257.
Kronbichler M., Hutzler F., Staffen W., et al. Evidence for a dysfunction of left posterior reading areas in German dyslexic readers. Neuropsychologia. 2006;44(10):1822-1832.
Lefly D.L., Pennington B.F. Spelling errors and reading fluency in compensated adult dyslexics. Ann Dyslexia. 1991;41:143-162.
Liberman I.Y., Shankweiler D. Phonology and beginning to read: A tutorial. In: Rieben L., Perfetti C.A., editors. Learning to read: Basic research and its implications. Hillsdale, NJ: Lawrence Erlbaum; 1991:3-17.
Lyon G., Fletcher J., Shaywitz S., et al. Rethinking learning disabilities. Washington, DC: Thomas B. Fordham Foundation and Progressive Policy Institute; 2001.
Meaburn E., Harlaar N., Craig I., et al. Quantitative trait locus association scan of early reading disability and ability using pooled DNA and 100K SNP microarrays in a sample of 5760 children. Mol Psychiatry. 2008;13:729-740.
Morgan W.P. A case of congenital word blindness. Br Med J. 1896:1378.
Morris R.D., Stuebing K.K., Fletcher J.M., et al. Subtypes of reading disability: Variability around a phonological core. J Educ Psychol. 1998;90:347-373.
Nakamura K., Dehaene S., Jobert A., et al. Subliminal convergence of Kanji and Kana words: further evidence for functional parcellation of the posterior temporal cortex in visual word perception. J Cogn Neurosci. 2005;17(6):954-968.
Nakamura K., Hara N., Kouider S., et al. Task-guided selection of the dual neural pathways for reading. Neuron. 2006;52:557-564.
Paulesu E., Demonet J.-F., Fazio F., et al. Dyslexia-cultural diversity and biological unity. Science. 2001;291:2165-2167.
Pennington B., Gilger J. How is dyslexia transmitted? In: Chase C.H., Rosen G.D., Sherman G.F., editors. Developmental dyslexia. Neural, cognitive, and genetic mechanisms. Baltimore, MD: York Press; 1996:41-61.
Ramus F., Rosen S., Dakin S., et al. Theories of developmental dyslexia: insights from a multiple case study of dyslexic adults. Brain. 2003;126:841-865.
Report of the National Reading Panel. Teaching Children to Read: An Evidence Based Assessment of the Scientific Research Literature on Reading and its Implications for Reading Instruction. (Vol. NIH Pub. No. 00-4754). U.S. Department of Health and Human Services, Public Health Service, National Institutes of Health, National Institute of Child Health and Human Development; 2000.
Richlan F., Kronbichler M., Wimmer H. Functional Abnormalities in the Dyslexic Brain: A Quantitative Meta-Analysis of Neuroimaging Studies. Hum Brain Mapp. 2009;30:3299-3308.
Scarborough H.S. Very early language deficits in dyslexic children. Child Dev. 1990;61:1728-1743.
Shankweiler D., Crain S., Katz L., et al. Cognitive profiles of reading-disabled children: Comparison of language skills in phonology, morphology, and syntax. Psychol Sci. 1995;6(3):149-156.
Shankweiler D., Liberman I.Y., Mark L.S., et al. The speech code and learning to read. J Exp Psychol [Hum Learn]. 1979;5(6):531-545.
Share D.L., Stanovich K.E. Cognitive processes in early reading development: Accommodating individual differences into a model of acquisition. Issues in Education: Contributions from Educational Psychology. 1995;1(1):1-57.
Shaywitz B., Shaywitz S., Pugh K., et al. Disruption of posterior brain systems for reading in children with developmental dyslexia. Biol Psychiatry. 2002;52(2):101-110.
Shaywitz B., Skudlarski P., Holahan J., et al. Age-related changes in reading systems of dyslexic children. Ann Neurol. 2007;61:363-370.
Shaywitz B.A., Fletcher J.M., Holahan J.M., et al. Discrepancy compared to low achievement definitions of reading disability: Results from the Connecticut Longitudinal Study. J Learn Disabil. 1992;25(10):639-648.
Shaywitz B.A., Holford T.R., Holahan J.M., et al. A Matthew effect for IQ but not for reading: Results from a longitudinal study. Read Res Q. 1995;30(4):894-906.
Shaywitz B.A., Shaywitz S.E., Pugh K.R., et al. Disruption of posterior brain systems for reading in children with developmental dyslexia. Biol Psychiatry. 2002;52(2):101-110.
Shaywitz S. Dyslexia. Sci Am. 1996;275(5):98-104.
Shaywitz S. Current Concepts: Dyslexia. N Engl J Med. 1998;338(5):307-312.
Shaywitz S. Overcoming Dyslexia: A new and complete science-based program for reading problems at any level. New York, NY: Alfred A. Knopf; 2003.
Shaywitz S., Fletcher J., Holahan J., et al. Persistence of dyslexia: the Connecticut Longitudinal Study at adolescence. Pediatrics. 1999;104(6):1351-1359.
Shaywitz S., Shaywitz B. Dyslexia (Specific Reading Disability). Biol Psychiatry. 2005;57:1301-1309.
Shaywitz S., Shaywitz B., Fulbright R., et al. Neural systems for compensation and persistence: young adult outcome of childhood reading disability. Biol Psychiatry. 2003;54(1):25-33.
Shaywitz S., Shaywitz B., Pugh K., et al. Functional disruption in the organization of the brain for reading in dyslexia. Proc Natl Acad Sci USA. 1998;95:2636-2641.
Shaywitz S.E., Escobar M.D., Shaywitz B.A., et al. Evidence that dyslexia may represent the lower tail of a normal distribution of reading ability. N Engl J Med. 1992;326(3):145-150.
Shaywitz S.E., Shaywitz B.A. Paying attention to reading: The neurobiology of reading and dyslexia. Dev Psychopathol. 2008;20:1329-1349.
Snow C.E., Burns M.S., Griffin P., editors. Preventing reading difficulties in young children. Washington, DC: National Academy Press, 1998.
Stanovich K., Siegel L. Phenotypic performance profile of children with reading disabilities: A regression-based test of the phonological-core variable-difference model. J Educ Psychol. 1994;86(1):24-53.
Stuebing K., Fletcher J., LeDoux J., et al. Validity of IQ-Discrepancy Classifications of Reading Disabilities:A Meta-Analysis. Am Educ Res J. 2002;39(2):469-518.
Torgesen J., Wagner R., Rashotte C., et al. Preventing Reading Failure in Young Children with Phonological Processing Disabilities. J Educ Psychol. 1999;91:579-593.
Vinckier F., Dehaene S., Jobert A., et al. Hierarchical coding of letter strings in the ventral stream: dissecting the inner organization of the visual word-form system. Neuron. 2007;55(1):143-156.
Woodcock R.W., Johnson M.B.. Woodcock – Johnson Psycho – Educational Battery – Revised (WJ–R)-Allen TX: Developmental Learning Materials. 1989.
Xue G., Chen C., Jin Z., et al. Language experience shapes fusiform activation when processing a logographic artificial language: an fMRI training study. Neuroimage. 2006;31(3):1315-1326.