Chapter 8 Spinal Cord and Nerve Roots
The spinal cord provides innervation for the trunk and limbs through the paired spinal nerves and their peripheral ramifications. Through them it receives primary afferent fibres from peripheral receptors located in widespread somatic and visceral structures. It also sends motor axons to skeletal muscle and provides autonomic innervation of cardiac and smooth muscle and secretory glands. Many functions are regulated by intraspinal reflex connections. Profuse ascending and descending pathways link the spinal cord with the brain. They allow higher centres to monitor and perceive external and internal stimuli and modulate and control spinal efferent activity.
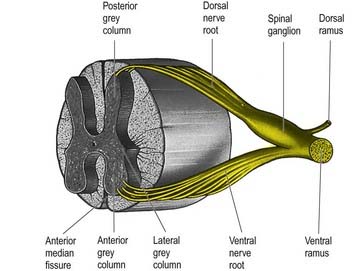
The spinal cord is essentially a segmental structure. It gives rise to 31 pairs of segmentally arranged spinal nerves, which are attached to the cord by a linear series of dorsal and ventral rootlets. Dorsal rootlets contain afferent nerve fibres, and ventral rootlets contain efferent fibres (see Fig. 1.5). Groups of adjacent rootlets coalesce to form dorsal or ventral nerve roots. These cross the subarachnoid space and unite to form functionally mixed spinal nerves as they pass through the intervertebral foramina. The dorsal roots bear dorsal root ganglia, which contain the cell bodies of primary afferent neurones.
Topographical Anatomy of the Spinal Cord
Macroscopic Anatomy of the Spinal Cord and Spinal Nerves
The gross anatomy of the structures that lie within the vertebral canal and its extensions through the intervertebral foramina, the spinal nerve or radicular (‘root’) canals is the subject of this chapter. The spinal cord, its blood vessels and nerve roots lie within a meningeal sheath, the theca, which occupies the central zone of the vertebral canal and extends from the foramen magnum, where it is in continuity with the meningeal coverings of the brain, to the level of the second sacral vertebra in the adult. Distal to this level the dura extends as a fine cord, the filum terminale externum, which fuses with the posterior periosteum of the first coccygeal segment. Tubular prolongations of the dural sheath extend around the spinal roots and nerves into the lateral zones of the vertebral canal and out into the ‘root’ canals, eventually fusing with the epineurium of the spinal nerves. Between the theca and the walls of the vertebral canal is the epidural (spinal extradural) space (Ch. 4), which is loosely filled with fat, connective tissue containing small arteries and lymphatics and an important venous plexus. Three-dimensional appreciation of the anatomy of the spinal theca and its surroundings is essential for the efficient management of spinal pain, spinal injuries, tumours and infections. Equally significant clinically is the anatomy of the often precarious blood supply of the spinal cord and its associated structures. The increasing application and refinement of diagnostic imaging and endoscopic procedures lend new importance to topographical detail here.
Spinal Cord (Medulla)
The spinal cord is an elongated, approximately cylindrical part of the central nervous system, occupying the superior two-thirds of the vertebral canal (Figs 8.1–8.6). Its average length in European males is 45 cm; its weight is approximately 30 g (for dimensional data, consult Barson and Sands 1977). It extends from the upper border of the atlas to the junction between the first and second lumbar vertebrae; this lower level varies, and there is some correlation with the length of the trunk, especially in females. The termination may be as high as the caudal third of the twelfth thoracic vertebra or as low as the disc between the second and third lumbar vertebrae, and its position rises slightly in vertebral flexion. The spinal cord is enclosed in the dura, arachnoid and pia mater, separated from each other by the subdural and subarachnoid spaces, respectively. The former is a potential space, whereas the latter contains cerebrospinal fluid (CSF). The cord is continuous cranially with the medulla oblongata and narrows caudally to the conus medullaris, from whose apex a connective tissue filament, the filum terminale, descends to the dorsum of the first coccygeal vertebral segment. The spinal cord varies in transverse width, gradually tapering craniocaudally, except at the levels of the cervical and lumbosacral enlargements. It is not cylindrical, being wider transversely at all levels, especially in the cervical segments.
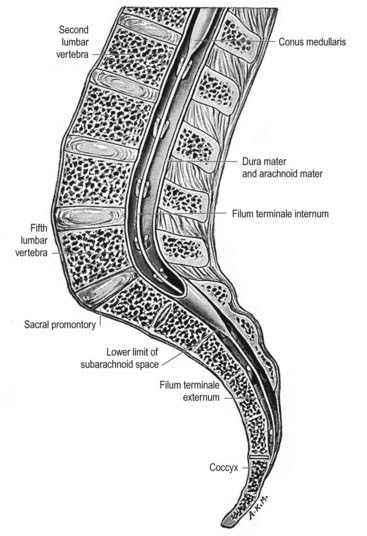
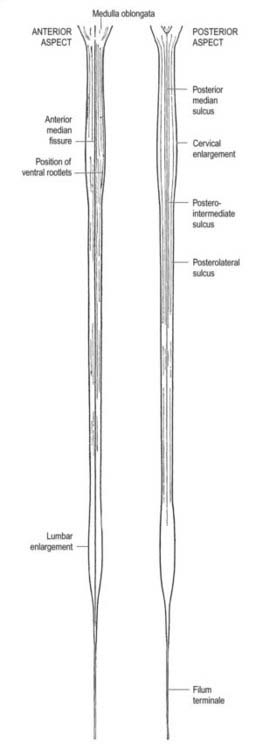

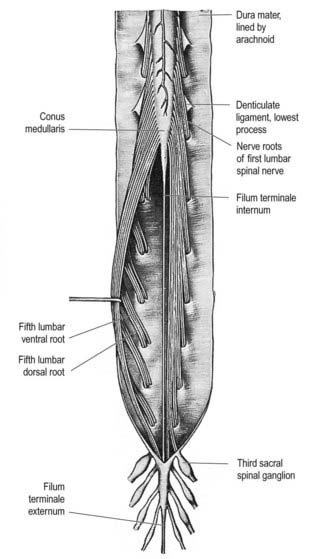
Fig. 8.3 Brain and spinal cord with attached spinal nerve roots and dorsal root ganglia, photographed from the dorsal aspect. Note the fusiform cervical and lumbar enlargements of the cord and the changing obliquity of the spinal nerve roots as the cord is descended. The cauda equina is undisturbed on the right but has been spread out on the left to show its individual components.
(Dissection by M. C. E. Hutchinson, GKT School of Medicine; photograph by Kevin Fitzpatrick on behalf of GKT School of Medicine, London.)
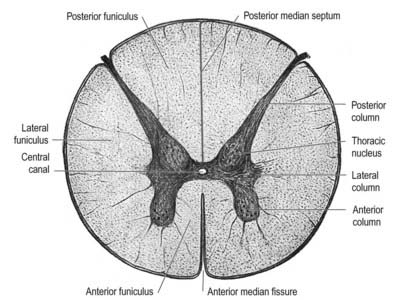
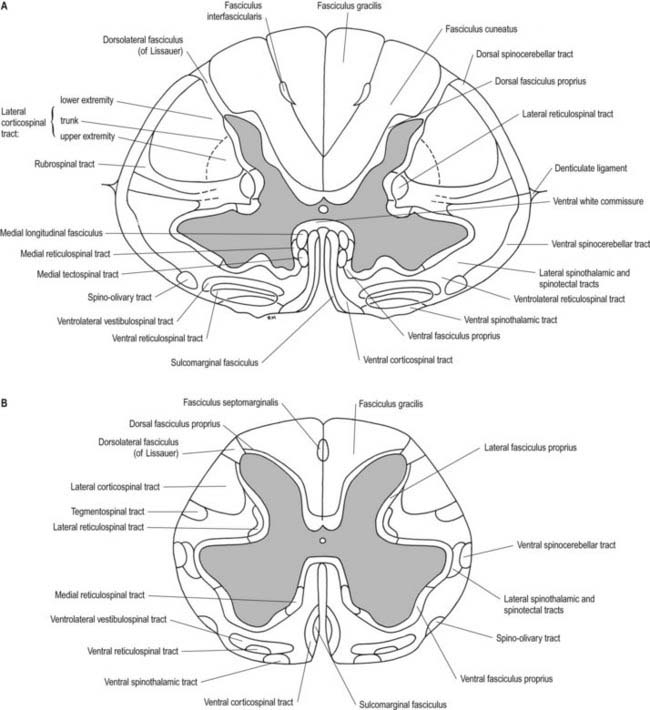
A posterolateral sulcus exists from 1.5 to 2.5 mm lateral to each side of the posterior median sulcus. Dorsal roots (strictly rootlets) of spinal nerves enter the cord along this sulcus. The white substance between the posterior median and posterolateral sulcus on each side is the posterior funiculus. In cervical and upper thoracic segments, a longitudinal posterointermediate sulcus marks a septum dividing each posterior funiculus into two large tracts: the fasciculus gracilis (medial) and the fasciculus cuneatus (lateral). Between the posterolateral sulcus and anterior median fissure is the anterolateral funiculus. This is subdivided into anterior and lateral funiculi by ventral spinal roots that pass through its substance to issue from the surface of the cord. The anterior funiculus is medial to, and includes, the emerging ventral roots; the lateral funiculus lies between the roots and the posterolateral sulcus. In upper cervical segments, nerve rootlets emerge through each lateral funiculus to form the spinal accessory nerve (cranial nerve XI), which ascends in the vertebral canal lateral to the spinal cord and enters the posterior cranial fossa via the foramen magnum (Fig. 8.7).
Dorsal and Ventral Roots
The paired dorsal and ventral roots of the spinal nerves are continuous with the spinal cord (see Figs 8.6 and 8.10). They cross the subarachnoid space and traverse the dura mater separately, uniting in or close to their intervertebral foramina to form the (mixed) spinal nerves. Because the spinal cord is shorter than the vertebral column, the more caudal spinal roots descend for varying distances around and beyond the cord to reach their corresponding foramina. In so doing they form, mostly distal to the apex of the cord, a divergent sheaf of spinal nerve roots, the cauda equina, which is gathered around the filum terminale in the spinal theca.
Meninges
Dura Mater
The single layer of dura that lines the cranial cavity divides into two layers as it passes downward through the foramen magnum, although it is still a single layer as it forms the anterior and posterior atlanto-occipital membranes. Within the vertebral column, it has been suggested that the outer endosteal layer becomes the periosteum of the vertebral canal, which is separated from the spinal dura mater by an extradural (epidural) space (see later). This interpretation, which would make the epidural space ‘intradural,’ is not generally agreed on (Newell 1999). The spinal dura mater forms a tube whose upper end is attached to the edge of the foramen magnum and to the posterior surfaces of the second and third cervical vertebral bodies, as well as to the posterior longitudinal ligament by fibrous bands, especially toward the caudal end of the vertebral canal. The dural tube narrows at the lower border of the second sacral vertebra. It invests the thin spinal filum terminale, descends to the back of the coccyx, and blends with the periosteum. The meningeal coverings of the spinal roots and nerves are described later in this chapter.
Epidural Space
The epidural space lies between the spinal dura mater and the tissues that line the vertebral canal (Fig. 8.8). It is closed above by fusion of the spinal dura with the edge of the foramen magnum, and below by the posterior sacrococcygeal ligament, which closes the sacral hiatus. It contains loosely packed connective tissue, fat, a venous plexus, small arterial branches, lymphatics and fine fibrous bands that connect the theca with the lining tissue of the vertebral canal. These bands, the meningovertebral ligaments, are best developed anteriorly and laterally. Similar bands tether the nerve root sheaths or ‘sleeves’ within their canals. There is also a midline attachment from the posterior spinal dura to the ligamentum nuchae at the atlanto-occipital and atlanto-axial levels (Dean and Mitchell 2002). The venous plexus consists of longitudinally arranged chains of vessels connected by circumdural venous ‘rings.’ The vertebral venous plexus is commonly known as Batson’s plexus. The anteriorly placed vessels receive the basivertebral veins.
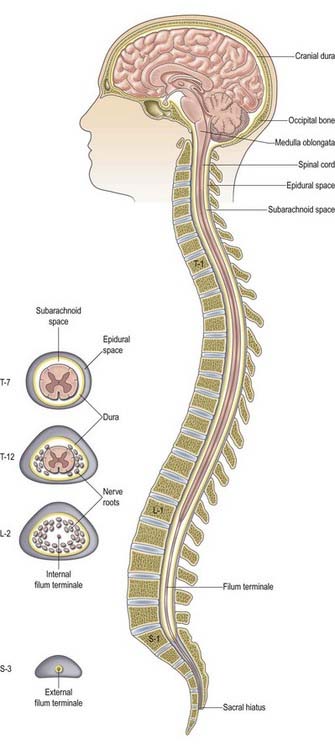
Fig. 8.8 Epidural space.
(Adapted with permission from Rosse, C., Gaddum-Rosse, P., 1997. Hollinshead’s Textbook of Anatomy, 5th ed. Lippincott-Raven, Philadelphia, Fig. 13-3.)
The shape of the space within each spinal segment is not uniform, although the segmental pattern is metamerically repeated. It is difficult to define the true shape of the ‘space’ because it changes with the introduction of fluid or as a result of preservation techniques. In the lumbar region, the dura mater is apposed to the walls of the vertebral canal anteriorly and attached by connective tissue in a manner that permits displacement of the dural sac during movement and venous engorgement. Adipose tissue is present posteriorly in recesses between the ligamentum flavum and the dura. The connective tissue extends for a short distance through the intervertebral foramina along the sheaths of the spinal nerves. Like the main thecal sac, the root sheaths are partially tethered to the walls of the foramina by fine meningovertebral ligaments. Contrast media and other fluids injected into the epidural space at the sacral level can spread up to the cranial base. Local anaesthetics injected near the spinal nerves, just outside the intervertebral foramina, may spread up or down the epidural space to affect the adjacent spinal nerves or may pass to the opposite side. The paravertebral spaces of each side communicate via the epidural space, particularly at the lumbar levels.
Subdural Space
The subdural space is a potential space in the normal spine because the arachnoid and dura are closely apposed (Haines, Harkey and Al-Mefty 1993). It does not connect with the subarachnoid space but continues for a short distance along the cranial and spinal nerves. Accidental subdural catheterization may occur during extradural injections. Injection of fluid into the subdural space may damage the cord either by direct toxic effects or by compression of the vasculature.
Arachnoid Mater
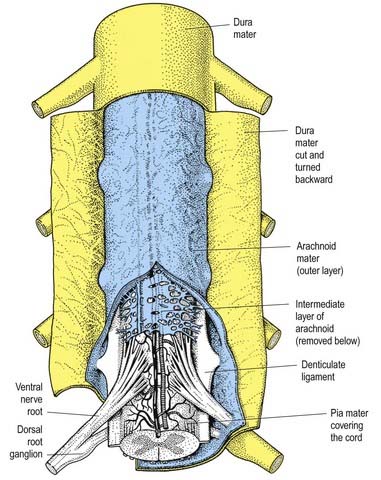
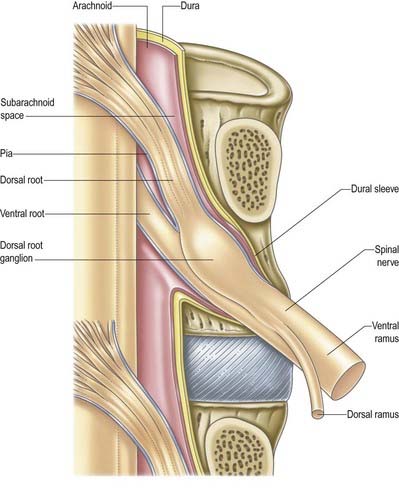
The arachnoid mater that surrounds the spinal cord is continuous with the cranial arachnoid mater (Figs 8.9, 8.10). It is closely applied to the deep aspect of the dura mater. At sites where vessels and nerves enter or leave the subarachnoid space, the arachnoid mater is reflected on to the surface of these structures and forms a thin coating of leptomeningeal cells over the surface of both vessels and nerves. Thus, a subarachnoid angle is formed as nerves pass through the dura into the intervertebral foramina. At this point, the layers of leptomeninges fuse and become continuous with the perineurium. The epineurium is in continuity with the dura. Such an arrangement seals the subarachnoid space so that particulate matter does not pass directly from the subarachnoid space into nerves. The existence of a pathway of lymphatic drainage from the CSF is controversial.
Pia Mater
The spinal pia mater closely invests the surface of the spinal cord and passes into the anterior median fissure (see Figs 8.9, 8.10). As in the cranial region, there is a subpial ‘space’; however, over the surface of the spinal cord the subpial collagenous layer is thicker than in the cerebral region, and it is continuous with the collagenous core of the ligamentum denticulatum.
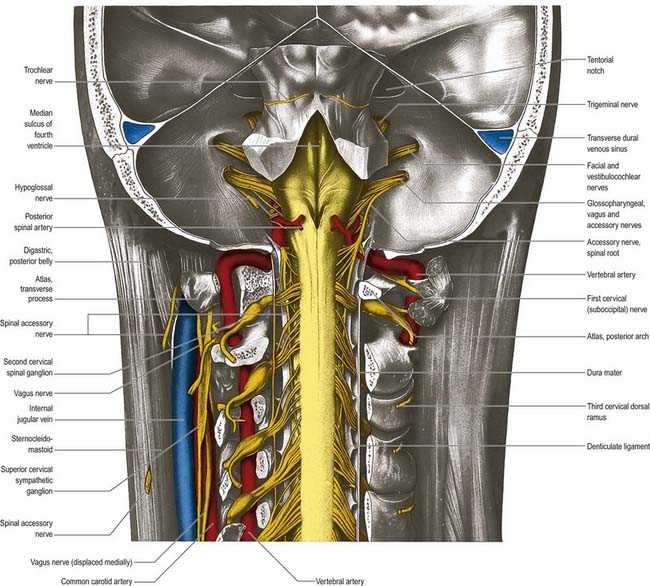
The ligamentum denticulatum is a flat, fibrous sheet that lies on each side of the spinal cord between the ventral and dorsal spinal roots. Its medial border is continuous with the subpial connective tissue of the cord, and its lateral border forms a series of triangular processes, the apices of which are fixed at intervals to the dura mater. There are usually 21 processes on each side. The first crosses behind the vertebral artery, where it is attached to the dura mater, and is separated by the artery from the first cervical ventral root. Its site of attachment to the dura mater is above the rim of the foramen magnum, just behind the hypoglossal nerve; the spinal accessory nerve ascends on its posterior aspect (see Fig. 8.7). The last of the dentate ligaments lies between the exiting twelfth thoracic and first lumbar spinal nerves and is a narrow, oblique band that descends laterally from the conus medullaris. Changes in the form and position of the dentate ligaments during spinal movements have been demonstrated by cineradiography.
Beyond the conus medullaris, the pia mater continues as a coating of the filum terminale.
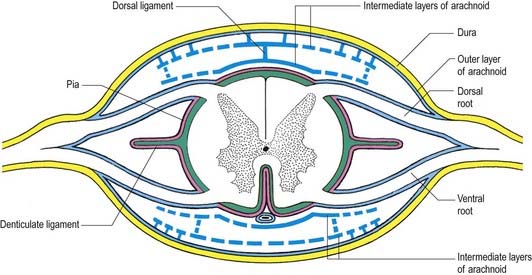
Intermediate Layer
In addition to the well-defined coats of arachnoid and pia mater, the cord is surrounded by an extensive intermediate layer of leptomeninges (see Fig. 8.10). This layer is concentrated in the dorsal and ventral regions and forms a highly perforated, almost lace-like structure that is focally compacted to form the dorsal, dorsolateral and ventral ligaments of the spinal cord. Dorsally, the intermediate layer is adherent to the deep aspect of the arachnoid mater and forms a discontinuous series of dorsal ligaments that attach the spinal cord to the arachnoid. The dorsolateral ligaments are more delicate and fenestrated, and they extend from the dorsal roots to the parietal arachnoid. As the intermediate layer spreads laterally over the dorsal surface of the dorsal roots, it becomes increasingly perforated and eventually disappears. A similar arrangement is seen over the ventral aspect of the spinal cord, but the intermediate layer is less substantial.
Spinal Nerves
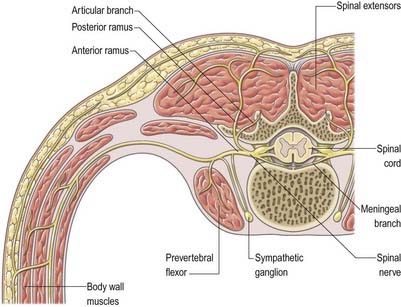
In those body segments that largely retain a metameric (segmental) structure (e.g. the thoracic region), spinal nerves exhibit a common plan (Fig. 8.11). The dorsal, epaxial ramus passes back, lateral to the articular processes, and divides into medial and lateral branches that penetrate the deeper muscles of the back; both branches innervate the adjacent muscles and supply a band of skin from the posterior median line to the scapular line. The ventral, hypaxial ramus is connected to a corresponding sympathetic ganglion by white and grey rami communicantes. It innervates the prevertebral muscles and curves around in the body wall to supply the lateral muscles of the trunk. Near the mid-axillary line it gives off a lateral branch that pierces the muscles and divides into anterior and posterior cutaneous branches. The main nerve advances in the body wall, where it supplies the ventral muscles, and terminates in branches to the skin.
Spinal Roots and Ganglia
Dorsal (Posterior) Roots
Little is known about the regions of entry and emergence of afferent and efferent rootlets in humans, but these zones of transition between the central and peripheral nervous systems have been extensively described in rodents (Fraher 2000).
Appearance and Orientation of Roots at Each Spinal Level
The size and direction of spinal nerve roots vary. The upper four cervical roots are small; the lower four are large. The thickness ratio of cervical dorsal roots to ventral roots is 3 : 1, which is greater than in the other regions. The first dorsal root is an exception, being smaller than the ventral root, and it is occasionally absent. The conventional view is that the first and second cervical spinal roots are short, running almost horizontally to their exits from the vertebral canal, and from the third to eighth cervical levels, the roots slope obliquely down. Obliquity and length increase successively, although the distance between spinal attachment and vertebral exit never exceeds the height of one vertebra. An alternative view is that upper cervical roots descend, the fifth is horizontal, the sixth to eighth ascend, the first two thoracic roots are horizontal, the next three ascend, the sixth is horizontal and the rest descend (Kubik and Müntener 1969). This view is based on the observation that the cervicothoracic part of the spinal cord grows more in length than the other parts do.
Lower lumbar and upper sacral roots are the largest, and their rootlets are most numerous. Coccygeal roots are the smallest. Kubik and Müntener (1969) confirm that lumbar, sacral and coccygeal roots descend with increasing obliquity to their exits. The spinal cord ends near the lower border of the first lumbar vertebra, and the lengths of successive roots rapidly increase; the consequent collection of roots is the cauda equina (see Fig. 8.3). The largest roots, and hence the largest spinal nerves, are continuous with the spinal cervical and lumbar swellings and innervate the upper and lower limbs.
Spinal Ganglia (Dorsal Root Ganglia)
Spinal ganglia are large groups of neurones on the dorsal spinal roots. Each is oval and reddish; its size is related to that of its root. A ganglion is bifid medially where the two fascicles of the dorsal root emerge to enter the cord. Ganglia are usually located in the intervertebral foramina, immediately lateral to the perforation of the dura mater by the roots (see Fig. 8.4). However, the first and second cervical ganglia lie on the vertebral arches of the atlas and axis, the sacral lies inside the vertebral canal, and the coccygeal ganglion usually lies within the dura mater. The first cervical ganglion may be absent. Small aberrant ganglia sometimes occur on the upper cervical dorsal roots between the spinal ganglia and the cord.
Spinal Nerves Proper
Immediately distal to the spinal ganglia, ventral and dorsal roots unite to form spinal nerves (Figs 8.11, 8.12). These soon divide into dorsal and ventral rami, both of which receive fibres from both roots. At all levels above the sacral, this division occurs within the intervertebral foramen. Division of the sacral spinal nerves occurs within the sacral vertebral canal, and the dorsal and ventral rami exit separately through posterior and anterior sacral foramina at each level. Spinal nerves trifurcate at some cervical and thoracic levels, and the third branch is called a ramus intermedius. At or distal to its origin, each ventral ramus gives off recurrent meningeal (sinuvertebral) branches and receives a grey ramus communicans from the corresponding sympathetic ganglion. The thoracic and first and second lumbar ventral rami each contributes a white ramus communicans to the corresponding sympathetic ganglia. The second, third and fourth sacral nerves also supply visceral branches, unconnected with sympathetic ganglia, that carry a parasympathetic outflow directly to the pelvic plexuses.
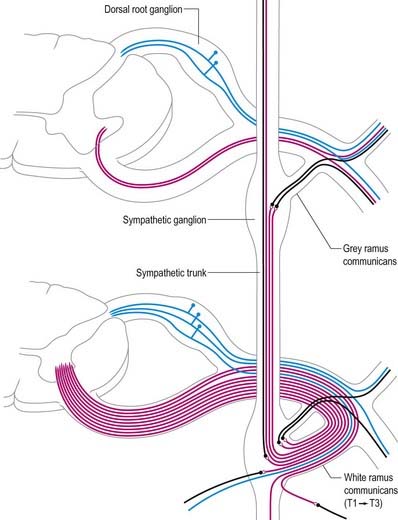
Coverings and Relations of the Spinal Roots and Nerves in the Radicular Canal
Tubular prolongations of the spinal dura mater, closely lined by the arachnoid, extend around the spinal roots and nerves as they pass through the lateral zone of the vertebral canal and through the intervertebral foramina (Figs 8.9, 8.10, 8.13). These prolongations, the spinal nerve sheaths (root sheaths), gradually lengthen as the spinal roots become increasingly oblique. Each individual dorsal and ventral root runs in the subarachnoid space with its own covering of pia mater. Each root pierces the dura separately, taking a sleeve of arachnoid with it, before joining within the dural prolongation just distal to the spinal ganglion. The dural sheaths of the spinal nerves fuse with the epineurium, within or slightly beyond the intervertebral foramina. The arachnoid prolongations within the sheaths do not extend as far distally as their dural coverings, but the subarachnoid space and its contained CSF extend sufficiently distally to form a radiologically demonstrable ‘root sleeve’ for each nerve. Shortening or obstruction of this sleeve seen on myelography indicates compression of the spinal nerve. At the cervical level, where the nerves are short and the vertebral movement is greatest, the dural sheaths are tethered to the periosteum of the adjacent transverse processes. In the lumbosacral region there is less tethering of the dura to the periosteum, although there may be an attachment posteriorly to the facet joint capsule.
Functional Components of Spinal Nerves
Each typical spinal nerve contains somatic and visceral (autonomic) fibres.
Variations of Spinal Roots and Nerves
The courses of spinal roots and nerves in relation to the thecal sac and vertebral and radicular canals may be aberrant. An individual intervertebral foramen may contain a duplicated sheath, nerve and roots, which are then absent at an adjacent level. Abnormal communications between roots may occur within the vertebral canal. These anomalies have been described and classified for the lumbosacral spine by Neidre and Macnab (1983).
Rami of the Spinal Nerves
Dorsal (posterior primary) rami of spinal nerves are usually smaller than the ventral rami and are directed posteriorly. Retaining a segmental distribution, they all (except for the first cervical, fourth and fifth sacral and coccygeal) divide into medial and lateral branches that supply the muscles and skin of the posterior regions of the neck and trunk (Fig. 8.14).
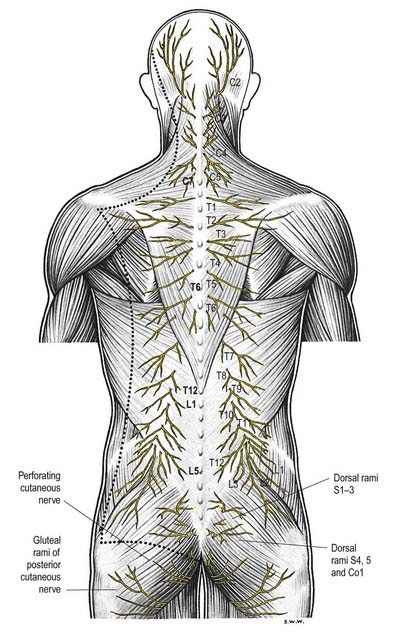
Cervical dorsal spinal rami
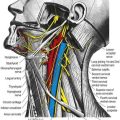
The first cervical dorsal ramus, the suboccipital nerve (see Fig. 8.7), is larger than the ventral. It emerges superior to the posterior arch of the atlas and inferior to the vertebral artery and enters the suboccipital triangle to supply the rectus capitis posterior major and minor, obliquus capitis superior and inferior and semispinalis capitis. A filament from the branch to the inferior oblique joins the second dorsal ramus. The suboccipital nerve occasionally has a cutaneous branch that accompanies the occipital artery to the scalp and connects with the greater and lesser occipital nerves. It may also communicate with the accessory nerve.
The second cervical dorsal ramus is slightly larger than the ventral and all the other cervical dorsal rami (see Figs 8.7, 11.10B). It emerges between the posterior arch of the atlas and the lamina of the axis, below the inferior oblique, which it supplies. It receives a connection from the first cervical dorsal ramus and divides into a large medial and smaller lateral branch. The medial branch, termed the ‘greater occipital nerve’, ascends between the inferior oblique and semispinalis capitis, pierces the latter and trapezius near their occipital attachments and is joined by a filament from the medial branch of the third dorsal ramus. It ascends with the occipital artery, divides into branches that connect with the lesser occipital nerve and supplies the skin of the scalp as far forward as the vertex. It supplies the semispinalis capitis and, occasionally, the back of the auricle. The lateral branch supplies the splenius capitis, longissimus capitis and semispinalis capitis and is often joined by the corresponding third cervical branch.
Dermatomes
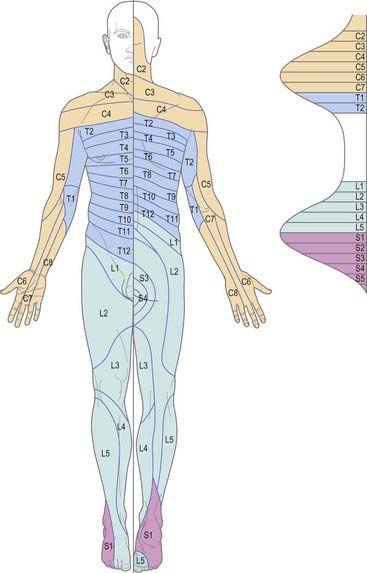
The cutaneous area supplied by one spinal nerve, through both rami, is a dermatome (Fig. 8.15). Typically, dermatomes extend around the body from the posterior to the anterior median line. The upper half of each zone is supplemented by the nerve above, and the lower half by the nerve below. The area supplied by dorsal rami is limited laterally by the dorsolateral line, which descends laterally from the occiput to the medial end of the acromion, continues to the posterior aspect of the greater trochanter and curves medially to the coccyx. Cutaneous strips supplied by dorsal rami do not correspond exactly to those served by ventral rami and differ in both breadth and position. Dermatomes of adjacent spinal nerves overlap markedly, particularly in the segments least affected by development of the limbs (i.e. second thoracic to first lumbar). In some regions (e.g. upper anterior thoracic wall), cutaneous nerves supplying adjoining areas are not from consecutive spinal nerves, and the overlap is minimal. When the second thoracic spinal ramus is severed, anaesthesia is sharply demarcated, but some overlap for awareness of painful and thermal stimuli may exist. Likewise, after section of a peripheral nerve (e.g. ulnar nerve at the wrist), the area of tactile loss is always greater than that of lost pain and temperature sensation. Hence, the area of total anaesthesia and analgesia following section of peripheral nerves is always less than might be anticipated from their anatomical distribution.
Internal Organization
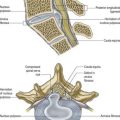
In transverse section, the spinal cord is incompletely divided into symmetric halves by a dorsal (posterior) median septum and a ventral (anterior) median sulcus (see Fig. 8.5). It consists of an outer layer of white matter and an inner core of grey matter. The dimensions and relative volumes of white matter and centrally aggregated neurone cell bodies vary according to the level (Fig. 8.16). The amount of grey matter at any level is a function of the amount of muscle, skin and other tissues innervated by neurones at that level. It is therefore largest by proportion in the cervical and lumbar enlargements, because neurones in these segments of the cord innervate the limbs; it is attenuated at thoracic levels. The absolute amount of white matter is greatest at cervical levels and decreases progressively at lower levels; this is the case because descending tracts shed fibres as they descend, and ascending tracts accumulate fibres as they ascend.
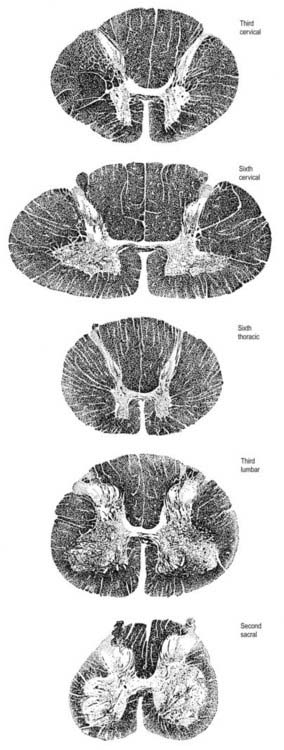
Spinal Grey Matter
In three dimensions, the spinal grey matter is shaped like a fluted column (see Fig. 8.6). In transverse section (see Fig. 8.16) the column is often described as being ‘butterfly shaped’ or resembling the letter ‘H.’ It consists of four cellular masses, referred to as the dorsal and ventral horns (or columns), which project dorsolaterally and ventrolaterally toward the surface. The grey matter that immediately surrounds the central canal and unites the two sides is termed the dorsal and ventral grey commissure. The dorsal horns are the site of termination of primary afferent fibres, which enter via the dorsal roots of spinal nerves. The tip of the dorsal horn is separated from the surface of the cord by a thin dorsolateral tract (tract of Lissauer) in which primary afferents ascend and descend for a short distance before terminating in the subjacent grey matter (Figs 8.17, 8.18). The dorsal horn may be described in terms of a head, neck and base, the individual constituents of which are described in more detail later. The ventral horns contain efferent neurones whose axons leave the spinal cord in ventral nerve roots. A small intermediate lateral horn is present at the thoracic and upper lumbar levels and contains the cell bodies of preganglionic sympathetic neurones.
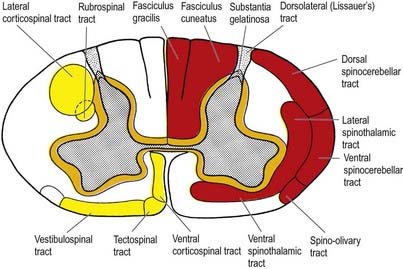
Fig. 8.17 Simplified diagram of the main tracts of the spinal cord. The ascending tracts are shown in red on the right side of the figure; the descending tracts are shown in yellow on the left side; the ‘intersegmental’ tracts are shown in orange on both sides. Many tracts are omitted (see Fig. 8.18).
Spinal grey matter is a complex mixture of neuronal cell bodies (somata), their processes (neurites) and synaptic connections, neuroglia and blood vessels (Fig. 8.19). Neurones in the grey matter are multipolar and vary in size and other features, particularly the length and arrangement of their axons and dendrites. Many are Golgi types I and II neurones. Axons of Golgi type I neurones pass out of the grey matter into ventral spinal roots or spinal tracts. Axons and dendrites of Golgi type II neurones are largely confined to the nearby grey matter. The distribution of neurones may be intrasegmental—deployed within a single segment—or intersegmental—spread through several segments.
CASE 1 Postpolio Syndrome
Discussion: A diagnosis of postpolio syndrome is established. This is a poorly understood disorder that may be caused by a gradual dropout of motor units or muscle fibres as a result of aging superimposed on residual anterior horn cells that were previously depleted as a result of the original poliomyelitis (Fig. 8.20). Increased metabolic demand on the maximally reinnervated motor units may also play a significant role.
CASE 2 Amyotrophic Lateral Sclerosis
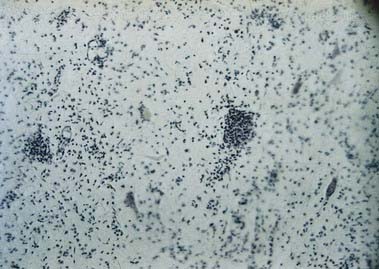
Discussion: Although a combination of upper and lower motor neurone symptoms and signs can be found in a variety of clinical conditions, this patient’s progressive history of motor symptoms over 6 months, with upper and lower motor neurone signs in three extremities in the absence of sensory and autonomic disturbances, points to a progressive motor neurone disorder—in this case, amyotrophic lateral sclerosis (Lou Gehrig’s disease, motor system disease). Some afflicted patients develop signs of frontotemporal degeneration as well. Pathologically, degeneration is observed primarily in the anterior horns of the spinal cord, in motor cranial nerve nuclei, and in the lateral corticospinal tracts. Although it begins at spinal levels, the disease generally spreads to involve motor cranial nerve nuclei, with resultant dysarthria, dysphagia and impaired respiratory function. See Figure 8.21.
Neuronal Cell Groups of the Spinal Cord
Viewed from the perspective of its longitudinal columnar organization, the grey matter of the spinal cord consists of a series of discontinuous cell groupings associated with their corresponding segmentally arranged spinal nerves. At any particular spinal level (as seen in transverse section), the spinal grey matter is considered to consist of 10 layers, called Rexed’s laminae, which are defined on the basis of neuronal size, shape, cytological features and density. The laminae are numbered sequentially in a dorsoventral sequence (Fig. 8.22).
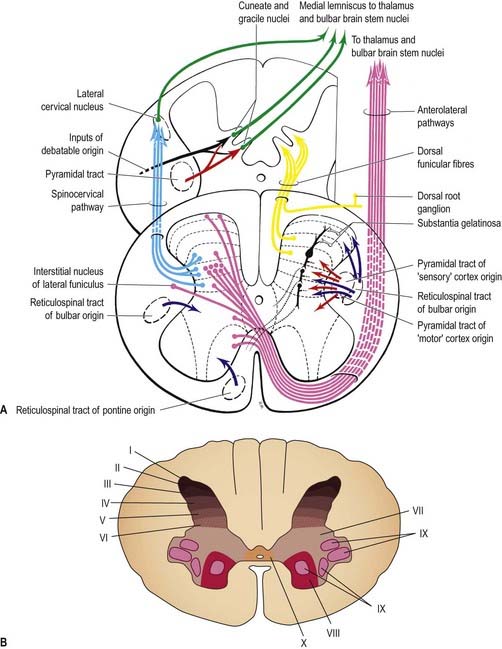
Laminae I to IV correspond to the head of the dorsal horn and are the main receiving areas for cutaneous primary afferent terminals and collateral branches. Many complex polysynaptic reflex paths (ipsilateral, contralateral, intrasegmental and intersegmental) start from this region, and many long ascending tract fibres that pass to higher levels arise from it. Lamina I (lamina marginalis) is a very thin layer with an ill-defined boundary at the dorsolateral tip of the dorsal horn. It has a reticular appearance, reflecting its content of intermingling bundles of coarse and fine nerve fibres. It contains small, intermediate and large neuronal somata, many of which are fusiform in shape. Lamina II occupies most of the head of the dorsal horn and consists of densely packed small neurones, accounting for its dark appearance in Nissl-stained sections. With myelin stains, lamina II is characteristically distinguished from adjacent laminae by the almost total lack of myelinated fibres. Lamina II corresponds to the substantia gelatinosa. Lamina III consists of somata that are mostly larger, more variable and less closely packed than those in lamina II. It also contains many myelinated fibres. Some workers believe that the substantia gelatinosa contains part or all of lamina III as well as lamina II. The ill-defined nucleus proprius of the dorsal horn corresponds to some of the cell constituents of laminae III and IV. Lamina IV is a thick, loosely packed, heterogeneous zone permeated by fibres. Its neuronal somata vary considerably in size and shape: small and round, intermediate and triangular, very large and stellate.
Laminae VII to IX show a variety of complex forms in the limb enlargements (see Fig. 8.22). Lamina VII includes much of the intermediate (lateral) horn. It contains prominent neurones of Clarke’s column (nucleus dorsalis, nucleus thoracicus, thoracic nucleus) and intermediomedial and intermediolateral cell groupings (Fig. 8.23). The lateral part of lamina VII has extensive ascending and descending connections with the midbrain and cerebellum (via the spinocerebellar, spinotectal, spinoreticular, tectospinal, reticulospinal and rubrospinal tracts) and is thus involved in the regulation of posture and movement. Its medial part has numerous propriospinal reflex connections with the adjacent grey matter and segments concerned with both movement and autonomic functions. Lamina VIII spans the base of the thoracic ventral horn but is restricted to its medial aspect in limb enlargements. Its neurones display a heterogeneous mixture of sizes and shapes from small to moderately large. Lamina VIII is a mass of propriospinal interneurones. It receives terminals from the adjacent laminae, many commissural terminals from the contralateral lamina VIII and descending connections from the interstitiospinal, reticulospinal and vestibulospinal tracts and the medial longitudinal fasciculus. The axons from these interneurones influence motor neurones bilaterally, perhaps directly but more probably by excitation of small neurones supplying γ-efferent fibres to muscle spindles. Lamina IX is a complex array of cells (Fig. 8.24) consisting of α and γ motor neurones and many interneurones. The large α motor neurones supply motor end-plates of extrafusal muscle fibres in striated muscle. Recording techniques have demonstrated tonic and phasic α motor neurones. The former have a lower rate of firing and lower conduction velocity and tend to innervate type S muscle units. The latter have higher conduction velocity and tend to supply fast twitch (type FR, FF) muscle units. The smaller γ motor neurones give rise to small-diameter efferent axons (fusimotor fibres), which innervate the intrafusal muscle fibres in muscle spindles. There are several functionally distinct types of γ motor neurone. The ‘static’ and ‘dynamic’ responses of muscle spindles have separate controls mediated by static and dynamic fusimotor fibres, which are distributed variously to nuclear chain and nuclear bag fibres.
Lamina X surrounds the central canal and consists of the dorsal and ventral grey commissures.
Dorsal Horn
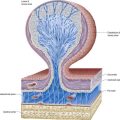
The dorsal horn is a major receptive zone (zone of termination) of primary afferent fibres, which enter the spinal cord through the dorsal roots of spinal nerves. Dorsal root fibres contain numerous molecules that are either known or suspected to fulfill a neurotransmitter or neuromodulator role. These include glutamate, substance P, calcitonin gene–related peptide, bombesin, vasoactive intestinal polypeptide, cholecystokinin (CCK), somatostatin, dynorphin and angiotensin II. Dorsal root afferents carry exteroceptive, proprioceptive and interoceptive information. Laminae I to IV are the main cutaneous receptive areas; lamina V receives fine afferents from the skin, muscle and viscera; and lamina VI receives proprioceptive and some cutaneous afferents. Most if not all primary afferent fibres divide into ascending and descending branches on entering the cord. These then travel for variable distances in the tract of Lissauer, near the surface of the cord, and send collaterals into the subjacent grey matter. The formation, topography and division of dorsal spinal roots have all been confirmed in humans.
At the dorsolateral tip of the dorsal horn, deep to the tract of Lissauer, lies a thin lamina of neurones, the lamina marginalis. Beneath this lies the substantia gelatinosa (laminae II and III), which is present at all levels and consists mostly of small Golgi type II neurones, together with some larger neurones. It receives afferents via the dorsal roots and is the site of origin of the spinothalamic tract complex. The large cells of the nucleus proprius lie ventral to the substantia gelatinosa (see Fig. 8.23). These propriospinal neurones link segments for the mediation of intraspinal coordination.
Ventral Horn
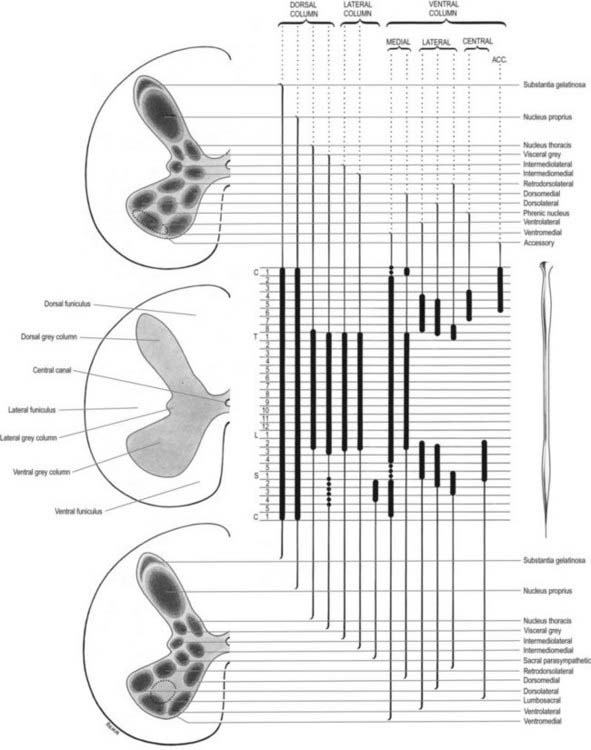
Considered longitudinally, ventral horn neurones are arranged in elongated groups and form a number of separate columns that extend through several segments. These are seen most easily in transverse sections (see Fig. 8.19). The ventral horn is essentially divided into medial, central and lateral cell columns, all of which are subdivided at certain levels, usually into dorsal and ventral parts. As can be seen in Figure 8.23, the medial group extends throughout the cord but may be absent in the fifth lumbar and first sacral segments. In the thoracic and the upper four lumbar segments, it is subdivided into ventromedial and dorsomedial groups. In segments cranial and caudal to this region, the medial group has only a ventromedial moiety, except in the first cervical segment, where only the dorsomedial group exists.
The central group of cells is the least extensive and is found only in some cervical and lumbosacral segments. The third to seventh cervical segments contain the centrally situated phrenic nucleus; abundant experimental and clinical evidence shows that its neurones innervate the diaphragm. Neurones whose axons are thought to enter the spinal accessory nerve form an irregular accessory group in the upper five or six cervical segments at the ventral border of the ventral horn (see Fig. 8.23).
The lateral group of cells in the ventral horn is subdivided into ventral, dorsal and retrodorsal groups, largely confined to the spinal segments that innervate the limbs. Their extents are indicated in Figure 8.23. The nucleus of Onuf, which is thought to innervate the perineal striated muscles, is a ventrolateral group of cells in the first and second sacral segments.
The motor neurones of the ventral horn are somatotopically organized. The basic arrangement is that medial cell groups innervate the axial musculature, and lateral cell groups innervate the limbs. The basic building block of the somatic motor neuronal populations is represented by a longitudinally disposed group of neurones that innervate a given muscle and in which the α and γ motor neurones are intermixed. The various groups innervating different muscles are aggregated into two major longitudinal columns: medial and lateral. In transverse section these form the medial and lateral cell groups in the ventral horn (Figs 8.24, 8.25).
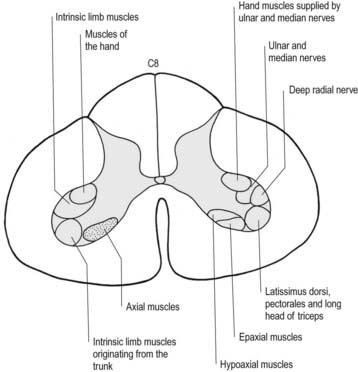
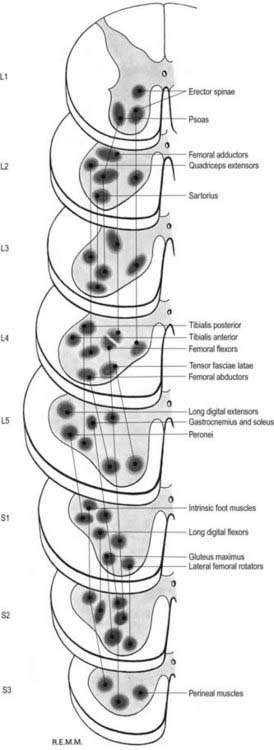
The lateral longitudinal motor column is found only in the enlargements of the spinal cord. The motor neurones in this column in the cervical and lumbar enlargements innervate muscles of the upper and lower limbs, respectively. In the cervical enlargement, motor neurones that supply muscles intrinsic to the upper limb are situated dorsally in the ventral grey column, and those innervating the most distal (hand) muscles are located farther dorsally. Motor neurones of the girdle muscles lie in the ventrolateral part of the ventral horn (see Fig. 8.25). There is a further somatotopic organization, in that the proximal muscles of the limb are supplied from motor cell groups located more rostrally in the enlargement than those supplying the distal muscles. For example, motor neurones innervating intrinsic muscles of the hand are sited in segments C8 and T1, whereas motor neurones of shoulder muscles are in segments C5 and C6. A similar overall arrangement of motor neurones innervating lower limb muscles applies in the lumbosacral cord (Fig. 8.26).
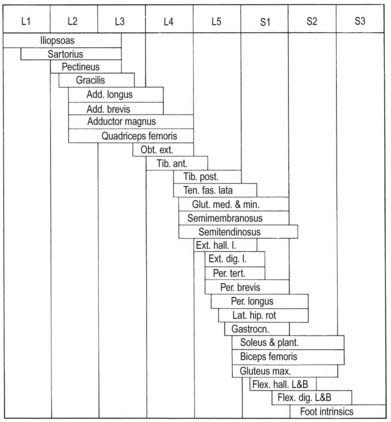
Fig. 8.26 Segmental arrangement of innervation of the lower limb muscles.
(Reproduced with permission and copyright © of the British Editorial Society of Bone and Joint Surgery [From Sharrard 1964. Sharrard WJ. Posterior iliopsoas transplantation in the treatment of paralytic dislocation of the hip. J Bone Joint Surg BR. 1964 Aug; 46: 426–444, Fig. 2.].)
Spinal Reflexes
The intrinsic connections of the spinal cord and brain stem subserve a number of reflexes by which the functions of peripheral structures are modulated in response to afferent information in a relatively automatic or autonomous fashion. The fundamental components of such reflex ‘arcs’ are thus an afferent and an efferent neurone. However, in all but the simplest of reflexes, interneurones intervene between the afferent and efferent components and confer the capacity to increase the versatility and complexity of reflex responses. Reflexes, by their very nature, are relatively fixed and stereotyped in form. Nevertheless, they are strongly influenced and modulated by descending connections. In the case of spinal reflexes, these descending controls come from both the brain stem and the cerebral cortex. Pathology of descending supraspinal pathways commonly causes abnormalities of spinal reflex activity, which are routinely tested for during neurological examination. During development, descending control mechanisms suppress what may be regarded as ‘primitive’ reflex responses. However, these may be released or uncovered in certain pathological conditions, such as the plantar (Babinski’s) and grasp reflexes.
Stretch reflex
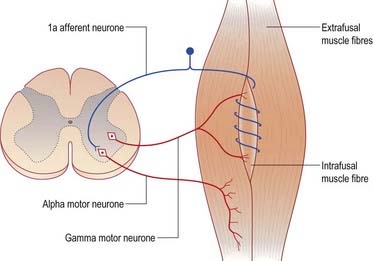
The stretch reflex is the mechanism by which stretch applied to a muscle elicits its reflex contraction. It is essential for the maintenance of both muscle tone and an upright stance (via innervation of the postural muscles of the neck, back and lower limbs). Anatomically, it is the simplest of reflexes—it consists of one afferent and one efferent neurone. The afferent component arises from stretch receptors associated with intrafusal muscle fibres located within muscle spindles; their primary or anulospiral endings give rise to primary afferent fibres, which enter the spinal cord and make excitatory synaptic contact directly on α motor neurones innervating the same muscle (Fig. 8.27). The α motor neurones of antagonistic muscles are simultaneously inhibited via collateral connections to inhibitory interneurones.
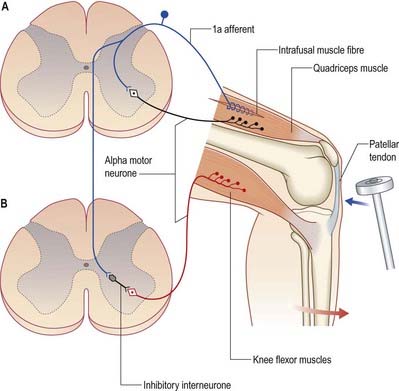
Gamma reflex
In addition to α motor neurones innervating extrafusal muscle fibres, muscles receive γ motor neurones that innervate intrafusal muscle fibres. Activation of γ motor neurones increases the sensitivity of the intrafusal fibres to stretch (Fig. 8.28). Therefore, changes in γ activity have a profound effect on the stretch reflex and on muscle tone. Like α motor neurones, γ motor neurones are under the influence of descending pathways from the brain stem and cerebral cortex. Changes in the activity of the stretch reflex and of muscle tone are commonly found in disorders of both the central and peripheral nervous systems.
Flexor reflex
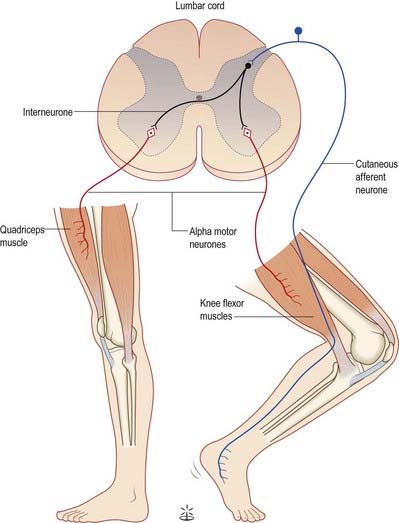
Painful stimulation of the limbs leads to reflex flexion withdrawal mediated by a polysynaptic reflex in which interneurones are interposed between afferent and efferent elements (Fig. 8.29). Thus, activation of nociceptive primary afferents indirectly causes activation of limb flexor motor neurones. Collateralization of fibres to nearby spinal segments mediates flexion of a limb at several joints, depending on the intensity of the stimulus. Decussating connections to the contralateral side of the cord activate α motor neurones innervating corresponding extensor muscles, which produces the so-called ‘crossed extensor reflex’. In principle, virtually any cutaneous stimulus has the potential to induce a flexor reflex, but with the exception of noxious stimuli, this response is normally inhibited by descending pathways. When descending influences are lost, even harmless cutaneous stimulation can elicit flexion of the limbs. The Babinski (extensor plantar) reflex, which is generally regarded as pathognomonic of damage to the corticospinal tract, at least in adults, is part of a flexion withdrawal of the lower limb in response to stimulation of the sole of the foot.
Spinal White Matter
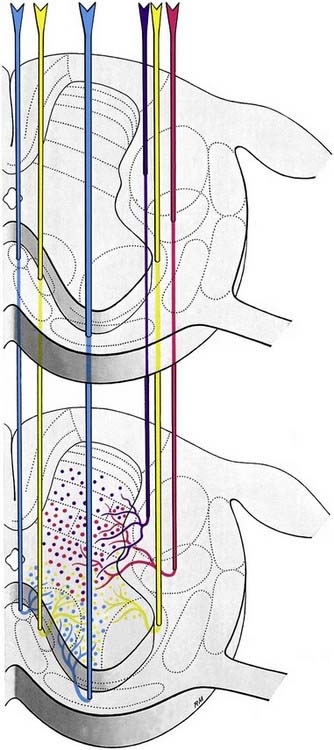
The spinal white matter surrounds the central core of grey matter. It contains nerve fibres, neuroglia and blood vessels. Most of the nerve fibres run longitudinally. They are arranged in three large masses—the dorsal, lateral and ventral funiculi—on either side of the cord (see Fig. 8.5). Fibres of related function and those with common origins or destinations are grouped to form ascending and descending tracts within the funiculi. Narrow dorsal and ventral white commissures run between the two halves of the cord. Here, the tracts are considered under three main headings: ascending, descending and propriospinal. Ascending tracts contain primary afferent fibres, which enter by dorsal roots, and fibres derived from intrinsic spinal neurones, which carry afferent impulses to supraspinal levels. Descending tracts contain long fibres, which descend from various supraspinal sources to synapse with spinal neurones. Propriospinal tracts, both ascending and descending, contain the axons of neurones that are localized entirely to the spinal cord and link nearby and distant spinal segments. They mediate intrasegmental and intersegmental coordination.
Although the ascending and descending tracts are to a large extent discrete and regularly located, significant overlap between adjacent tracts occurs. The following account of spinal tracts is concerned with the human cord; findings in animals are discussed only when adequate clinicopathological data are unavailable in humans. The general disposition of the major tracts is shown in Figure 8.17 and is depicted in greater detail at two transverse levels in Figure 8.18. Other features are summarized in Figures 8.22 and 8.30.
CASE 3 Subacute Combined Degeneration
Discussion: Weakness, spasticity and hyperreflexia in the legs suggest involvement of the corticospinal tracts within the spinal cord (Fig. 8.31). Impaired vibratory and position sense indicates involvement of the posterior columns, with relative sparing of the spinothalamic sensory pathways. This is a typical clinical presentation of subacute combined degeneration of the cord due to vitamin B12 deficiency, documented with appropriate testing. The mild impairment of cutaneous sensibility indicates the coexistence of a polyneuropathy, similarly due to vitamin B12 deficiency. Dementia and visual impairment due to involvement of the optic nerves and cerebral white matter may occur.
Ascending Tracts
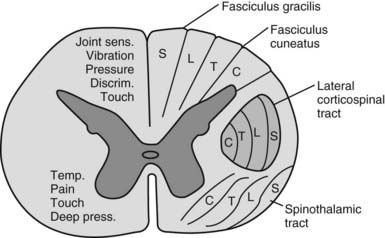
Dorsal columns
The dorsal funiculus on each side of the cord consists of two large ascending tracts—the fasciculus gracilis and fasciculus cuneatus (Fig. 8.32)—separated by a posterointermediate septum. They are also known as the dorsal columns. The dorsal columns contain a high proportion of myelinated fibres carrying proprioception (position sense and kinaesthesia) and exteroceptive (touch-pressure) information, including vibratory sensation, to higher levels. These fibres come from several sources: long primary afferent fibres that enter the cord in the dorsal roots of spinal nerves and ascend to the dorsal column nuclei in the medulla oblongata, shorter primary afferent fibres projecting to neurones of Clarke’s column and other spinal neurones and axons from secondary neurones of the spinal cord ascending to the dorsal column nuclei. The dorsal columns also contain axons of propriospinal neurones.
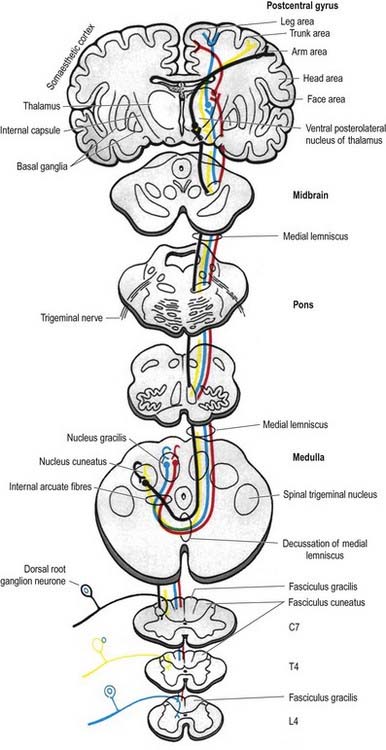
The fasciculus gracilis lies medial to the fasciculus cuneatus in the upper spinal cord (see Fig. 8.18). At upper cervical levels, the fasciculus gracilis contains a larger proportion of afferents from cutaneous receptors than from deep proprioceptors because many of the latter leave the fasciculus at lower segments to synapse in Clarke’s column. Indeed, proprioception from the lower limb reaches the thalamus mostly by relaying in Clarke’s column and again in the nucleus Z. Axons of the fasciculus gracilis, from both primary and secondary neurones, terminate in the nucleus gracilis of the dorsal medulla.
The fasciculus cuneatus (see Fig. 8.32) begins at mid-thoracic level and lies lateral to the fasciculus gracilis. It is composed mostly of primary afferent fibres of the upper thoracic and cervical dorsal roots. At upper cervical levels, it contains a large population of afferents from both deep and cutaneous receptors of the upper limb. In addition, some of its axons arise from secondary neurones in laminae IV to VI of the ipsilateral dorsal horn. Many axons (both primary and secondary) that ascend in the fasciculus cuneatus terminate in the nucleus cuneatus of the dorsal medulla. Some also end in the lateral (external or accessory) cuneate nucleus, whose neurones project to the cerebellum via the cuneocerebellar pathway.
Many ascending fibres of the fasciculus gracilis and fasciculus cuneatus terminate by synapsing on neurones of the dorsal column nuclei (nucleus gracilis and nucleus cuneatus, respectively) in the medulla oblongata. (The connections of the dorsal column nuclei are described further with the medulla oblongata in Chapter 10.) Axons arising from neurones in the dorsal column nuclei arch ventromedially around the central grey matter of the medulla as internal arcuate fibres (see Fig. 10.6) and decussate in the sensory or lemniscal decussation. At the medial lemniscus, they ascend to the ventral posterolateral nucleus of the thalamus; from there, neurones project to the somatosensory cortex in the postcentral gyrus of the parietal lobe (areas 3, 1 and 2). Some neurones of the dorsal column nuclei form posterior external arcuate fibres, which enter the cerebellum.
Spinocerebellar tracts
There are two principal spinocerebellar tracts: dorsal or posterior, and ventral or anterior. They occupy the periphery of the lateral aspect of the spinal white matter and carry proprioceptive and cutaneous information to the cerebellum for the coordination of movement (Fig. 8.33). Both tracts contain large-diameter myelinated fibres, but there are more in the posterior tract. Finer-calibre fibres are associated with the anterior tract.
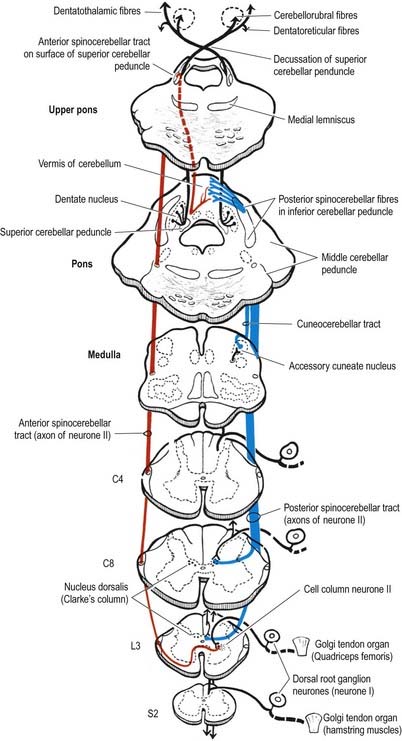
The dorsal spinocerebellar tract lies lateral to the lateral corticospinal tract (see Fig. 8.17). It begins at about the level of the second or third lumbar segment and enlarges as it ascends. Axons of the tract originate ipsilaterally from the larger neurones of Clarke’s column, in lamina VII throughout spinal segments T1 to L2. Clarke’s column receives input from collaterals of long ascending primary afferents of the doral columns and terminals of shorter ascending primary afferents of the dorsal columns. Many of these afferent fibres ascend from segments caudal to L2. In the medulla, the dorsal spinocerebellar tract passes through the inferior cerebellar peduncle to terminate ipsilaterally in the rostral and caudal parts of the cerebellar vermis.
The ventral spinocerebellar tract (see Fig. 8.33) lies immediately ventral to the dorsal spinocerebellar tract (see Fig. 8.17). The cells of origin are in laminae V to VII of the lumbosacral cord, and the tract carries information from the lower limb. Axons forming the tract mostly decussate; a few remain ipsilateral. The tract begins in the upper lumbar region and ascends through the medulla oblongata to reach the upper pontine level; it then descends in the dorsal part of the superior cerebellar peduncle and terminates, mainly contralaterally, in the anterior cerebellar vermis.
Because Clarke’s column diminishes rostrally (see Fig. 8.23) and does not extend above the lowest cervical segment, it follows that the dorsal spinocerebellar tract carries information from the trunk and lower limb. Proprioceptive and exteroceptive information from the upper limb travels in primary afferent fibres of the fasciculus cuneatus. These fibres end somatotopically in the accessory (external or lateral) cuneate nucleus and the adjoining part of the cuneate nucleus situated in the medulla oblongata. Cells of these nuclei give rise to the posterior external arcuate fibres that form the cuneocerebellar tract (see Fig. 8.33), which enters the cerebellum via the ipsilateral inferior cerebellar peduncle. The accessory cuneate nucleus and the lateral part of the cuneate nucleus are considered homologous to the cells of Clarke’s column. The cuneocerebellar tract is therefore functionally allied to the dorsal spinocerebellar tract and is its upper limb equivalent.
Axons of all the spinocerebellar tracts and the cuneocerebellar tract form part of the ‘mossy-fibre’ system. They end in the cerebellar cortex in a highly organized, somatotopic and functional pattern (Ch. 13).
CASE 4 Friedreich’s Ataxia
Discussion: As with all the so-called spinocerebellar ataxias, the core clinical deficit in Friedreich’s ataxia is a progressively incapacitating cerebellar deficit. Sensory changes involving especially proprioception are important and contribute to, if not actually causing, the ataxia (so-called sensory ataxia). The disease is clearly not confined to the cerebellar proprioceptive pathways, given the widespread clinical deficits such as areflexia, extensor plantar responses, optic atrophy and occasional deafness, along with non-neurological issues such as scoliosis, pes cavus and cardiomyopathy. Neuropathological changes include marked degeneration in Clarke’s column of the spinal cord as well as in the posterior columns, often with gross shrinkage of the spinal cord, degeneration in the corticospinal tracts and variable changes in other ascending and descending pathways (Fig. 8.34). The cerebellum itself is often normal. Generally inherited as an autosomal recessive trait, Friedreich’s ataxia is characterized by expanded trinucleotide (GAA) repeats.
Spinothalamic tracts
The lateral spinothalamic tract (Fig. 8.35) is sited in the lateral funiculus, lying medial to the ventral spinocerebellar tract (see Fig. 8.17). Clinical evidence indicates that it subserves pain and temperature sensations. The ventral spinothalamic tract (Fig. 8.36) lies in the anterior funiculus medial to the point of exit of the ventral nerve roots and dorsal to the vestibulospinal tract (see Fig. 8.17), which it overlaps. On the basis of clinical evidence, it subserves coarse tactile and pressure modalities.
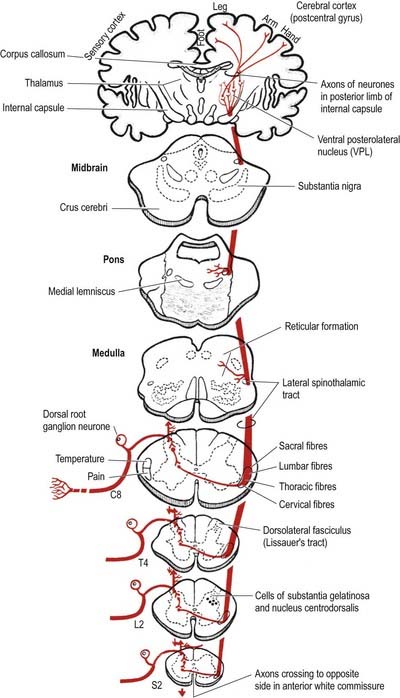
Fig. 8.35 Lateral spinothalamic tract.
(Reprinted by permission from Carpenter, M.B., 1991. Core Text of Neuroanatomy, 4th ed. Baltimore, Williams and Wilkins.)
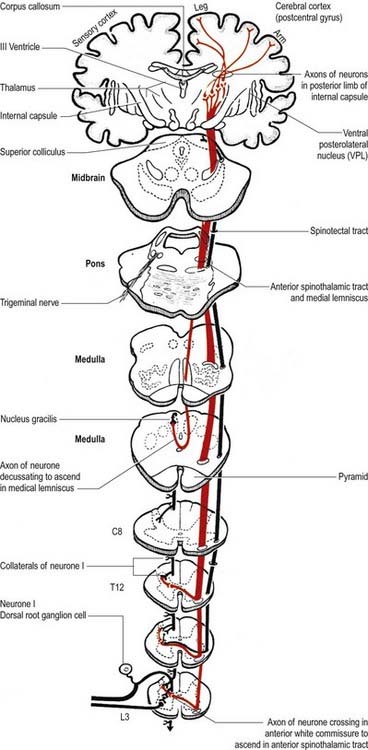
Fig. 8.36 Ventral (anterior) spinothalamic tract.
(Reprinted by permission from Carpenter, M.B., 1991. Core Text of Neuroanatomy, 4th ed. Baltimore, Williams and Wilkins.)
There is clear somatotopic organization of the fibres in the spinothalamic tracts throughout their extent. Fibres crossing at any cord level join the deep aspect of those that have already crossed, which means that both tracts are segmentally laminated (Fig. 8.37). Somatotopy is maintained throughout the medulla oblongata and pons. In the midbrain, fibres in the spinal lemniscus conveying pain and temperature sensation from the lower limb extend dorsally, whereas those from the trunk and upper limb are more ventrally placed. Both lemnisci ascend to end in the thalamus (see Figs 8.35, 8.36). The major spinothalamic projections in humans are to the ventral posterolateral nucleus and to the centrolateral intralaminar nucleus of the thalamus.
(Modified from Brodal A: Neurological Anatomy, 2e. New York, Oxford Univ. Press, 1969, Figure 2-8. By permission of Oxford University Press.)
CASE 5 Syringomyelia
Discussion: The disassociated sensory loss, of which the patient was originally unaware, along with unilateral muscular atrophy and loss of reflexes is the classic expression of syringomyelia. Early lesions are typically found within the cervical cord. The cyst generally appears at the base of the posterior horn and extends into the central grey and anterior commissure of the cord, thus involving the decussating spinothalamic fibres (Fig. 8.38). When the cyst extends into the anterior horn, lower motor neurone signs such as atrophy, weakness and reflex loss appear. Horner’s syndrome indicates spread of the lesion into the intermediolateral column. With the passage of time, the cystic lesion enlarges and may involve much of the remainder of the cord, with widespread motor and sensory deficits. The lesion commonly extends inferiorly in the cord and superiorly into the lower brain stem (syringobulbia).
Neurones of the spinothalamic tract
The population of deep dorsal column (laminae IV to VI) spinothalamic neurones of the lumbar cord contains wide dynamic range (60%), high-threshold (30%) and low-threshold (10%) units. They can accurately code both innocuous and noxious cutaneous stimuli. Some cells also respond to input from deep somatic and visceral receptors. In the lumbar cord their receptive fields are small or medium-sized—larger than the area of the foot but smaller than the entire leg. In the thoracic cord the fields of these laminar cells are larger and often include the entire upper limb plus part of the chest. Many of the deep dorsal column spinothalamic tract neurones in the thoracic segments receive convergent input from sympathetic afferent fibres. Laminae IV to VI spinothalamic tract units project to either the ventral posterolateral nucleus or the centrolateral nucleus of the thalamus, and sometimes to both. Units projecting to the ventral posterolateral nucleus receive input from all classes (Aβ, Aδ and C) of cutaneous fibres.
Spinoreticular pathway
Spinoreticular fibres are intermingled with those of the spinothalamic tracts and ascend in the ventrolateral quadrant of the spinal cord (Fig. 8.39). Evidence from animal studies suggests that the cells of origin occur at all levels of the spinal cord, particularly in the upper cervical segments. Most neurones are in lamina VII, some are in lamina VIII, and others are in the dorsal horn, especially lamina V. Most axons in the lumbar and cervical enlargements cross the midline, but there is a large uncrossed component in cervical regions. Most axons are myelinated. The pattern of anterograde degeneration, in both human postmortem studies and experimental animals following anterolateral cordotomy, indicates spinoreticular projections to many nuclei of the medial pontomedullary reticular formation. There is also a projection to the lateral reticular nucleus (a precerebellar relay nucleus). No somatotopic arrangement has been reported. Spinoreticular neurones respond to inputs from the skin or deep tissues. Innocuous cutaneous stimuli may inhibit or excite a particular cell, whereas noxious stimuli are often excitatory. A spinoreticulothalamocortical pathway has been proposed as an important route serving pain perception. Like other ascending pathways, the tract cells are influenced by descending control. For example, electrical stimulation of the periaqueductal grey matter inhibits the responses of certain spinoreticular cells to input from cardiopulmonary afferents. Stimulation of the reticular formation also alters the activity of spinoreticular neurones.
Pain mechanisms
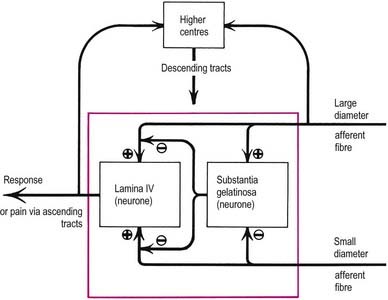
The gate control theory (Melzack and Wall 1965) defined a possible mechanism for modulating the inflow of information along nociceptive and other afferent pathways (Fig. 8.40). The proposition was that large-diameter afferents (e.g. from hairs and touch corpuscles) are excitatory to the large neurones of lamina IV, from which spinothalamic fibres arise, and to interneurones in the substantia gelatinosa. In contrast, fine non-myelinated afferents are excitatory to tract cells but inhibitory to interneurones. The axons of substantia gelatinosa interneurones are presumed to presynaptically inhibit the terminals of all afferents that synapse with tract cells. In such a system, low activity in the fine afferents inhibits the interneurones and thus prevents them from inhibiting tract cells; hence the ‘gate’ to T cells in lamina IV is open to transmit intermittent small volleys of impulses from the large fibres. A prolonged high-frequency volley of impulses in the large-diameter afferents would be transmitted to lamina IV tract cells initially, but this would soon cease as activity in the interneurones closed the gate. Conversely, persistent high activity in the fine afferents would open the gate, resulting in massive bombardment of neurones of lamina IV (which include some high-threshold neurones that are activated only by such bombardment). It was assumed that onward transmission in the lateral spinothalamic tract would evoke pain at supraspinal centres. Pain would therefore result from an imbalance between the varieties of afferent impulses when there was disproportionately large traffic along the fine afferents.
Descending Tracts
Descending pathways to the spinal cord originate primarily in the cerebral cortex and in numerous sites within the brain stem (Figs 8.39, 8.41, 8.42). They are concerned with the control of movement, muscle tone and posture; the modulation of spinal reflex mechanisms; and the transmission of afferent information to higher levels. They also mediate control over spinal autonomic neurones.
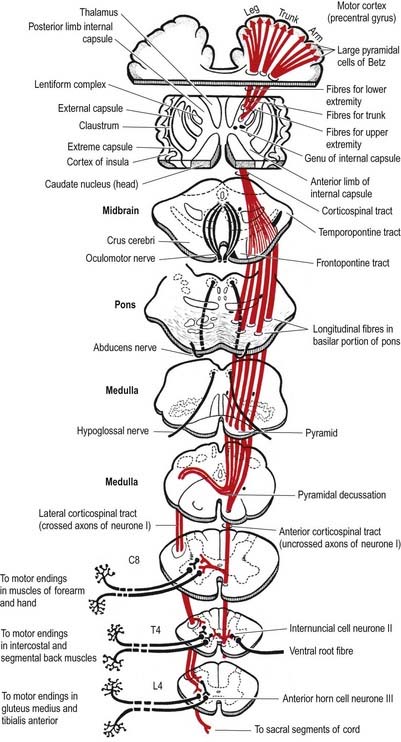
Fig. 8.41 Corticospinal tracts.
(Reprinted by permission from Carpenter, M.B., 1991. Core Text of Neuroanatomy, 4th ed. Baltimore, Williams and Wilkins.)
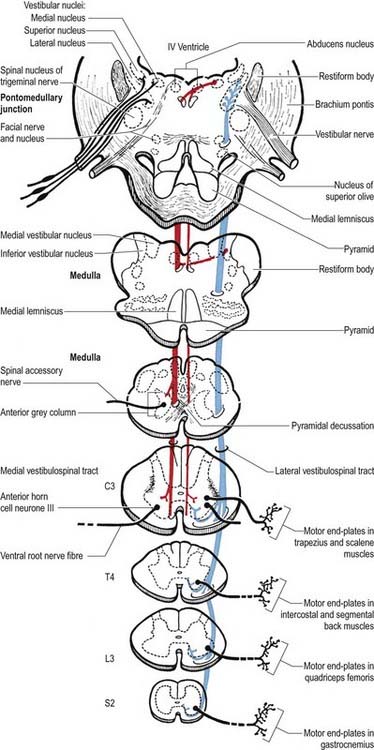
Fig. 8.42 Vestibulospinal tracts.
(Reprinted by permission from Carpenter, M.B., 1991. Core Text of Neuroanatomy, 4th ed. Baltimore, Williams and Wilkins.)
Corticospinal tract
Corticospinal fibres arise from neurones of the cerebral cortex. They project, in a somatotopically organized fashion, to neurones that are located mostly in the contralateral side of the spinal cord (see Fig. 8.41). The majority of corticospinal fibres arise from cells situated in the upper two-thirds of the precentral motor cortex (area 4) and from the premotor cortex (area 6). A small contribution of fibres comes from cells of the postcentral gyrus (somatosensory cortex, areas 1, 2, and 3) and the adjacent parietal cortex (area 5). In the monkey, 30% of corticospinal fibres arise from area 4, 30% from area 6 and 40% from the parietal regions. Cells of origin of corticospinal fibres vary in size in the different cortical areas and are clustered into groups or strips. The largest cells (giant pyramidal neurones, or Betz cells) are in the precentral cortex.
Corticospinal fibres descend first through the subcortical white matter and enter the posterior limb of the internal capsule. They then pass through the ventral part of the midbrain in the cerebral peduncle or crus cerebri. As they continue caudally through the pons, they are separated from its ventral surface by transversely running pontocerebellar fibres. In the medulla oblongata they form a discrete bundle, the pyramid (see Fig. 10.3), which forms a prominent longitudinal column on the ventral surface of the medulla. Thus, the corticospinal tracts are also referred to as the pyramidal tracts. However, this term is often used to denote not only corticospinal fibres but also corticobulbar fibres, which diverge above this level and end in association with cranial motor nuclei. Each pyramid contains about a million axons of varying diameters. The majority (70%) are myelinated; most (90%) have a diameter of 1 to 4 mm, 9% have diameters of 5 to 10 mm and less than 2% have diameters of 11 to 22 mm. The largest-diameter axons arise from the giant Betz cells.
Just rostral to the level of the spinomedullary junction, approximately 75% to 90% of the corticospinal fibres in the pyramid cross the median plane in the pyramidal decussation (decussation of the pyramids) and continue caudally as the lateral corticospinal tract. The rest of the fibres continue uncrossed as the ventral corticospinal tract. The lateral tract also contains some uncrossed corticospinal fibres. The lateral corticospinal tract (see Fig. 8.41) descends in the lateral funiculus throughout most of the length of the spinal cord. It occupies an oval area, ventrolateral to the dorsal horn and medial to the dorsal spinocerebellar tract (see Fig. 8.18). In the lumbar and sacral regions, where the dorsal spinocerebellar tract is absent, the lateral corticospinal tract reaches the dorsolateral surface of the cord. As it descends, the lateral corticospinal tract progressively diminishes in size until about the fourth sacral spinal segment. Its axons terminate on ipsilateral spinal neurones.
The smaller ventral corticospinal tract (see Fig. 8.41) descends in the ventral funiculus. It lies close to the ventral median fissure and is separated from it by the sulcomarginal fasciculus (see Fig. 8.18). The ventral corticospinal tract diminishes as it descends and usually disappears completely at mid-thoracic cord levels. It may be absent or, very rarely, contain almost all the corticospinal fibres. Near their termination, most fibres of the tract cross the median plane in the ventral white commissure to synapse with contralateral neurones. The vast majority of corticospinal fibres, irrespective of the tract in which they descend, terminate in the spinal cord on the side contralateral to their cortical origin.
Knowledge of the detailed termination of corticospinal fibres is based largely on animal studies but is supplemented by data from postmortem studies on human brains using anterograde degeneration methods. Most corticospinal fibres are believed to terminate contralaterally on interneurones in the lateral parts of laminae IV to VI and both lateral and medial parts of lamina VII. Some are also distributed to lamina VIII bilaterally. Terminals are associated with contralateral motor neuronal cell groups in lamina IX, in the dorsolateral group and in the lateral parts of both the central and ventrolateral groups (see Fig. 8.30).
Rubrospinal tract
The rubrospinal tract arises from neurones in the caudal magnocellular part of the red nucleus, an ovoid mass of cells situated centrally in the midbrain tegmentum (Ch. 10). This part of the nucleus contains some 150 to 200 large neurones, interspersed with smaller neurones.
The origin, localization, termination and functions of rubrospinal connections are poorly defined in humans, and the tract appears to be rudimentary. Rubrospinal fibres cross in the ventral tegmental decussation and descend in the lateral funiculus of the cord, where they lie ventral to, and intermingled with, fibres of the lateral corticospinal tract (see Fig. 8.18). In animals, the tract descends as far as lumbosacral levels, whereas in humans it appears to project only to the upper three cervical cord segments. Rubrospinal fibres are distributed to the lateral parts of laminae V and VI and the dorsal part of lamina VII of the spinal grey matter. The terminal zones of the tract correspond to those of corticospinal fibres from the motor cortex. Animal studies demonstrate that the effects of rubrospinal fibres on α and γ motor neurones are similar to those of corticospinal fibres.
Tectospinal tract
The tectospinal tract arises from neurones in the intermediate and deep layers of the superior colliculus of the midbrain. It crosses ventral to the periaqueductal grey matter in the dorsal tegmental decussation and descends in the medial part of the ventral funiculus of the spinal cord (see Fig. 8.17). Fibres of the tract project only to the upper cervical cord segments, ending in laminae VI to VIII. They make polysynaptic connections with motor neurones serving muscles in the neck, facilitating those that innervate contralateral muscles and inhibiting those that innervate ipsilateral ones. In animals, turning of the head to the contralateral side results from unilateral electrical stimulation of the superior colliculus and is effected mainly through the tectospinal tract.
Vestibulospinal tracts
The large vestibular nuclear complex lies in the lateral part of the floor of the fourth ventricle around the pontomedullary junction of the brain stem. It gives rise to the lateral and ventral vestibulospinal tracts, which are functionally and topographically distinct (see Fig. 8.42).
The lateral vestibulospinal tract arises from small and large neurones of the lateral vestibular nucleus (Deiters’ nucleus). It descends ipsilaterally, initially in the periphery of the ventrolateral spinal white matter but subsequently shifting into the medial part of the ventral funiculus at lower spinal levels. Fibres of this tract are somatotopically organized. Thus, fibres projecting to the cervical, thoracic and lumbosacral segments of the cord arise from neurones in the rostroventral, central and dorsocaudal parts, respectively, of the lateral vestibular nucleus. Lateral vestibulospinal fibres end ipsilaterally, mostly in the medial part of the ventral horn in lamina VIII and the medial part of lamina VII. Axons of the lateral vestibulospinal tract excite, through mono- and polysynaptic connections, motor neurones of extensor muscles of the neck, back and limbs; γ motor neurones are probably facilitated as well. Lateral vestibulospinal tract axons also inhibit, disynaptically, motor neurones of flexor limb muscles via 1a inhibitory interneurones.
The medial vestibulospinal tract arises mainly from neurones in the medial vestibular nucleus, but some are also located in the inferior and lateral vestibular nuclei. The medial vestibulospinal tract (see Fig. 8.42) descends in the medial longitudinal fasciculus into the ventral funiculus of the spinal cord, where it lies close to the midline in the so-called sulcomarginal fasciculus (see Fig. 8.18). Unlike the lateral tract, it contains both crossed and uncrossed fibres and does not extend beyond the mid-thoracic cord level. Fibres of the medial tract project mainly to the cervical cord segments, ending in lamina VIII and the adjacent dorsal part of lamina VII. Data from stimulation of the vestibular nuclei in animals indicate that axons of the medial tract monosynaptically inhibit the motor neurones that innervate axial muscles of the neck and upper part of the back.
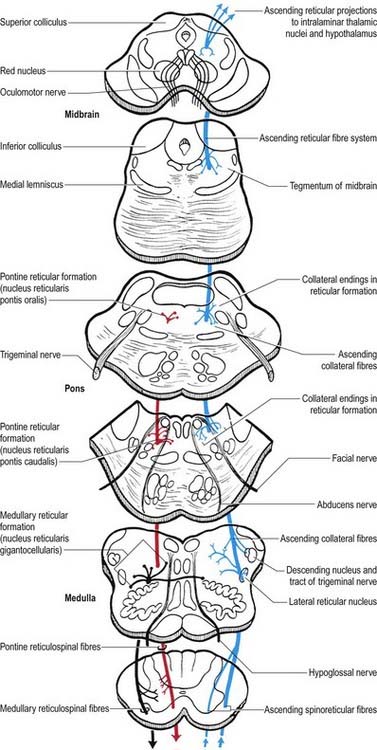
Reticulospinal tracts
The medial reticulospinal tract (see Fig. 8.39) originates from the medial tegmental fields of the pons and medulla. The main sources are the oral and caudal pontine reticular nuclei and the gigantocellular reticular nucleus in the medulla. Pontine fibres descend, mainly ipsilaterally, in the ventral funiculus of the cord. Medullary fibres descend, both ipsilaterally and contralaterally, in the ventral funiculus and ventral part of the lateral funiculus. These fibres have many collaterals, and two-thirds of the reticulospinal neurones that reach the cervical cord also descend to lumbosacral levels. The terminals of reticulospinal fibres are distributed to lamina VIII and the central and medial parts of lamina VII. The medullary reticulospinal terminals are more widely distributed, ending additionally in the lateral parts of laminae VI and VII and directly on motor neurones. From animal studies, it appears that terminations of reticulospinal fibres that originate in the medulla are, in general, more dorsally placed than those that originate in the pons, although there is considerable overlap.
The lateral reticulospinal tract lies in the lateral funiculus of the spinal cord, closely associated with the rubrospinal and lateral corticospinal tracts (see Fig. 8.18). Its fibres arise from neurones of the ventrolateral tegmental field of the pons. The fibres cross in the rostral medulla oblongata and project, with a high degree of collateralization, throughout the length of the spinal cord. Axons of this tract terminate in laminae I, V and VI and also bilaterally in the lateral cervical nucleus. Evidence suggests that this pathway is involved in the control of pain perception and in motor functions.
Propriospinal Tracts
Propriospinal pathways, or tracts, are sometimes referred to as the fasciculi proprii. Propriospinal neurones are confined to the spinal cord; that is, their ascending and descending fibres begin and end within the spinal grey matter. They connect neurones within the same segment or other neurones in more distant segments of the spinal cord and thus subserve intrasegmental and intersegmental integration and coordination. The majority of spinal neurones are propriospinal neurones, most of which lie in laminae V to VIII. Propriospinal fibres are concentrated mainly around the margins of the grey matter (see Fig. 8.17) but are also dispersed diffusely in the white funiculi.
Tract of Lissauer
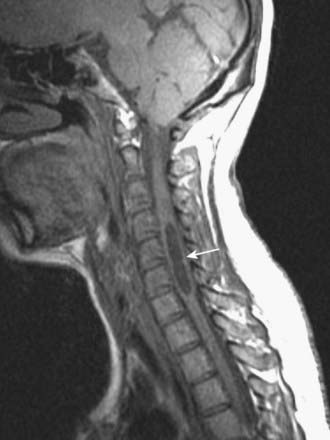
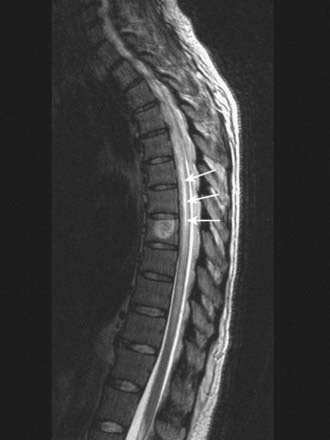
CASE 6 Acute Transverse Myelitis
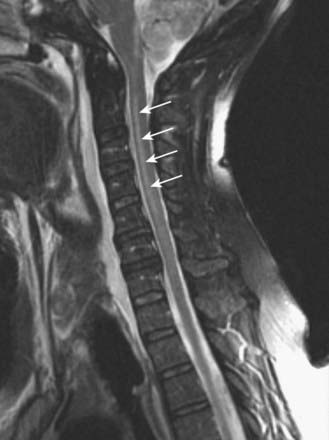
Discussion: Acute transverse myelitis is an acute or subacutely evolving idiopathic inflammatory disorder of the spinal cord, usually presenting as an acute sensorimotor segmental myelopathy. The lesions often involve several adjacent segments of the spinal cord; both grey and white matter are affected. The diagnosis is confirmed by MRI and spinal fluid analysis (Fig. 8.43).
Spinal Cord Injury and Vertebral Column Injury
CASE 7 Brown–Séquard Syndrome
A 30-year-old man is involved in a street brawl and is stabbed in the neck, with the knife penetrating at approximately the C4 level. Examination in the emergency room demonstrates a flaccid paralysis of the right arm and leg, with loss of reflexes; loss of vibratory and position sense below and ipsilateral to the level of the injury; and contralateral loss of pain and temperature sensation, also below the level of the lesion. MRI imaging demonstrates the wound penetrating the substance of the spinal cord (Fig. 8.44).
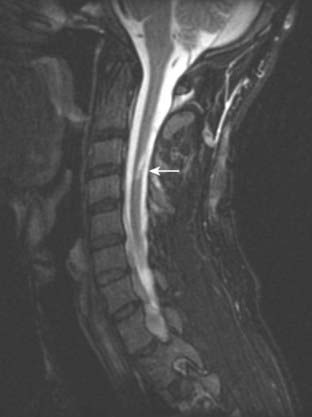
Lesions of the Spinal Roots, Nerves and Ganglia
Spinal Cord Lesions
Mechanical compression and secondary ischaemic damage to underlying nervous tissue cause surgically relevant spinal cord disease (myelopathy). The site and the level of cord damage determine the particular clinical syndrome—for example, whether the lesion involves the upper or lower cervical, thoracic or lumbosacral spinal cord. At each of these levels, symptoms and signs are determined by direct destruction of segmental tissue, or transversely distributed damage, and by disconnection of suprasegmental ascending and descending tracts above and below the level of a lesion, or longitudinally distributed damage (Fig. 8.45). For example, a lower cervical spinal cord lesion damages the segmental sensory and motor contributions to the nerve roots and brachial plexus, causing sensory loss, weakness and wasting of the muscles and loss of tendon reflexes in the upper limbs. Disruption of the ascending sensory pathways in the lateral and dorsal columns of the cervical spinal cord leads to loss of pain and temperature sensation (lateral spinothalamic tracts) and touch and proprioception (dorsal fasciculi) below the ‘sensory level’ corresponding to the spinal cord segment. Damage to the descending corticospinal tracts in the lateral columns of the spinal cord produces a spastic paraparesis, with increased muscle tone, weakness of flexion movements, exaggerated tendon reflexes and abnormal superficial reflexes (e.g. extensor plantar responses, absent abdominal reflexes). Descending pathways to the bladder are interrupted, producing a ‘neurogenic bladder.’ The same principles apply to lesions at other levels of the spinal cord, as illustrated in Figure 8.45.
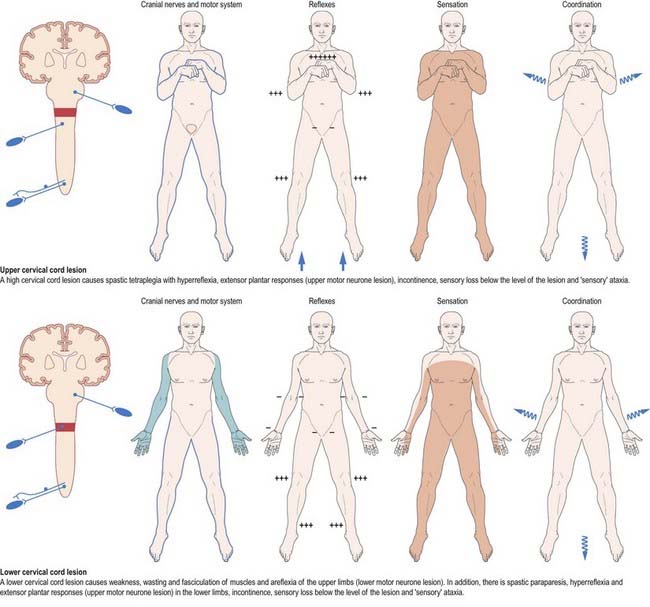
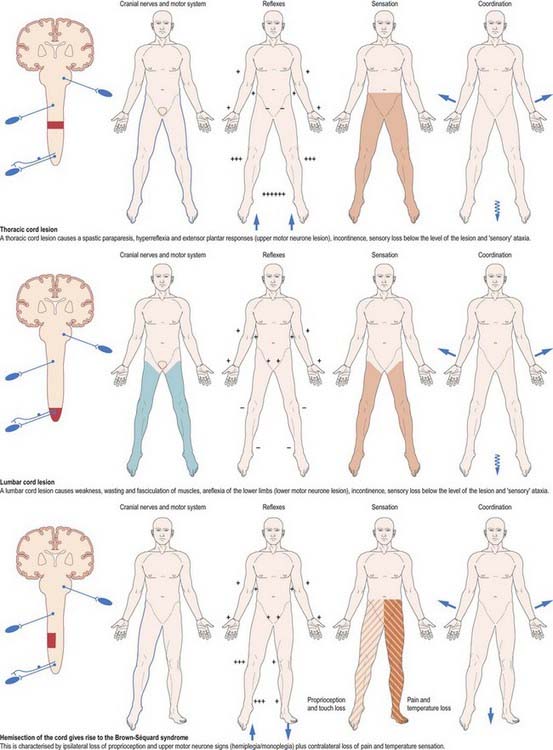
Fig. 8.45 Lesions of the spinal cord.
(By permission from Crossman, A.R., Neary, D., 2000. Neuroanatomy, 2nd ed. Churchill Livingstone, Edinburgh.)
The precise clinical syndrome is determined by anatomical site alone, not by pathology. However, it is of practical use to classify lesions on the basis of their anatomical relationship to the spinal cord and meninges—that is, whether they are extradural, intradural or intramedullary (Table 8.1). This anatomical classification provides a guide to the diagnostic probabilities as well as an aid to neuroradiological interpretation before neurosurgical intervention. For example, neurofibromas are common in the cervical spinal canal, meningiomas in the thoracic spinal canal and ependymomas in the lumbosacral spinal canal. Degenerative disease of the vertebral column is common in the cervical and lumbosacral vertebrae but rare in the thoracic vertebrae. Discrete anterior and central intramedullary lesions, such as those due to syringomyelia and angioma, respectively, preferentially destroy the spinothalamic pathways in the anterolateral columns and central areas of the spinal cord. This leads to a characteristic ‘dissociated’ sensory loss, with loss of pain and temperature sensation but preservation of touch sensation and proprioception at and below the level of the lesion.
Table 8.1 Classification of lesions based on anatomical relationship to the spinal cord and meninges
| Extradural Lesions |
CASE 8 Spinal Cord Infarction
Discussion: The anterior spinal artery supplies the anterior two-thirds of the spinal cord. Arising from the vertebral artery in the cervical region, it receives contributions from the medullary segmental arteries, the largest of which, the artery of Adamkiewicz, is in the thoracic region. Because there is little collateral circulation in the spinal cord, it is particularly sensitive to ischaemia; because the posterior one-third of the cord receives its blood supply from the posterior spinal arteries, posterior column function (e.g. proprioception) is often intact in the setting of anterior spinal artery disease. Spinal cord infarcts, which are most common in the thoracic region, may be due to systemic hypotension, with infarction at ‘border zones’ between vascular territories; surgery, particularly in the thoracic region, which may compromise the aorta or medullary arteries; direct injury to the anterior spinal artery with extension injury of the neck; rarely, fibrocartilaginous embolization in patients with preexisting spinal disc disease; or after relatively minor spinal trauma. See Figure 8.46.
Lesions of the Conus and Cauda Equina
Lesions of the conus and cauda equina, such as tumours, cause bilateral deficits, often with pain in the back extending into the sacral segments and to the legs. Loss of bladder and erectile function can be early features. There are lower motor neurone signs in the legs, with fasciculation and muscle atrophy. Sensory loss usually involves the perineal or ‘saddle area’ as well as other lumbar and sacral dermatomes. There may be congenital abnormalities, including spina bifida, lipoma or diastematomyelia, and the conus may extend below the lower border of L1, often with a tethered filum terminale. Extramedullary lesions include prolapsed intervertebral discs. A midline (central) disc protrusion in the lumbar region may present with involvement of only the sacral segments.
Barson A.J., Sands J. Regional and segmental characteristics of the human adult spinal cord. J. Anat.. 1977;123:797-803.
Bogduk N. Clinical Anatomy of the Lumbar Spine and Sacrum, third ed. Edinburgh: Churchill Livingstone; 1997.
Dean N.A., Mitchell B.S. Anatomic relation between the nuchal ligament (ligamentum nuchae) and the spinal dura mater in the craniocervical region. Clin. Anat.. 2002;15:182-185.
Fraher J.P. The transitional zone and CNS regeneration. J. Anat.. 2000;196:137-158.
Gordon P.H., Cheng B., Katz I.B., et al. The natural history of primary lateral sclerosis. Neurology. 2006;66:647-653.
Haines, Harkey, Al-Mefty, 1993Haines D.E. Harkey H.L. Al-Mefty O. The ‘subdural’ space: a new look at an outdated concept. Neurosurgery. 32:1993;111-120.
Kubik, Müntener, 1969Kubik S. Müntener M. Zur Topographie der spinalen Nervenwurzeln. II Der Einfuss des Wachstums des Duralsackes, sowie der Krümmagen und der Bewegungen der spinalen Nervenwurzeln. Acta. Anat.. 74:1969;149-168.
Kuypers H.G.J.M. Anatomy of descending pathways. In: Brookhart J.M., Mountcastle V.B., et al, editors. Handbook of Physiology, vol. 2. Bethesda, MD: American Physiological Society; 1981:597-666. Motor Control, pt 1
Mancall E.L., Rosales R.K. Necrotizing myelopathy associated with visceral carcinoma. Brain. 1964;87:639-656.
Melzack R., Wall P.D. Pain mechanisms: a new theory. Science. 1965;150:971-979.
Neidre A., Macnab I. Anomalies of the lumbosacral nerve roots. Spine. 1983;8:294-299.
Newell R.L.M. The spinal epidural space. Clin. Anat.. 1999;12:375-379.
Rothstein J.D. Current hypotheses for the underlying biology of amyotrophic lateral sclerosis. Ann. Neurol.. 2009;65:S3-S9.
Steele J.C., McGeer P.L. The ALS/PDS syndrome of Guam and the cycad hypothesis. Neurology. 2008;70:1984-1990.