CHAPTER 1 Phylogeny
EVOLUTION OF THE VERTEBRATE ELBOW
The distal humerus of pelycosaurs, the late Paleozoic (255 to 235 mya) reptiles that probably gave rise to more advanced mammal-like reptiles, possessed a bulbous capitellum laterally and medially. The articulation with the ulna was formed by two distinct surfaces: a slightly concave ventral surface and a more flat dorsal surface (Fig. 1-1).11 The proximal articular surface of the ulna was similarly divided into two surfaces separated by a low ridge. Reconstruction of the forelimb of these reptiles suggests that they walked with limbs splayed out to the side. The humerus was held more or less horizontal, the elbow flexed to 90 degrees, and the forearm was sagittally oriented. Forward motion was brought about by rotation of the humerus around its long axis, which propelled the body forward relative to the fixed forefoot. Elbow flexion and extension probably were useful only in side-to-side motions. The ulnohumeral joint, with its dual articular surfaces, was well suited to resist the valgus/varus stress produced by humeral rotation, and the proximal end of the radius was flat and triangular, precluding pronosupination. It appears, therefore, that stability rather than mobility was the major functional characteristic of the elbow of these late Paleozoic reptiles.
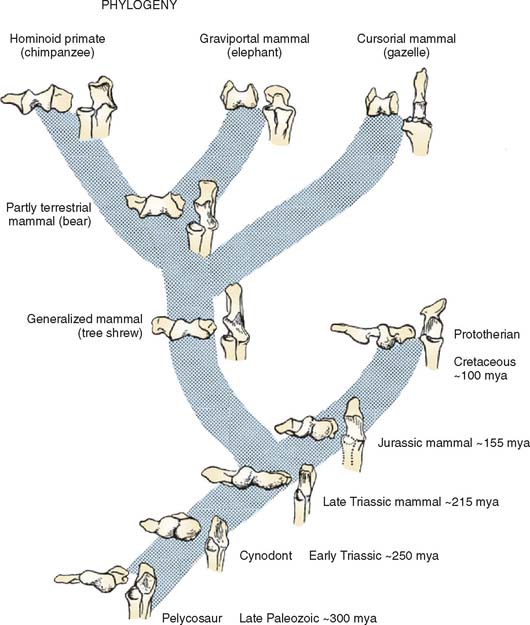
Cynodonts, the more immediate ancestors of mammals from the Permo-Triassic period (235 to 160 mya), had their limbs underneath their bodies rather than at the sides. The distal humeral articular surface consisted of radial and ulnar condyles separated by a shallow groove (see Fig. 1-1). The proximal ulnar articular surface was an elongate spoon shape for articulation with the humeroulnar condyle. The lateral flange on the ulna for articulation with the radius was separated from this surface by a low ridge. This ridge articulated with the groove between the radial and ulnar condyles displaying some features in common with the “tongue and groove” (trochleariform) type of humeroulnar articulation characteristic of many modern mammals.
Early mammals from the Triassic (210 to 160 mya) and Jurassic (160 to 130 mya) periods also had radial and ulnar condyles. However, the radial condyle was more protuberant than the ulnar, and the ulnar condyle was more linear and obliquely oriented (see Fig. 1-1). The two condyles were separated by an intercondylar groove. The ulnar notch had articular surfaces for both the ulnar and the radial condyles, each matching the configuration of the corresponding humeral surface. The oblique orientation of the humeroulnar joint resembled a spiral configuration, which helped to keep forearm movement in a sagittal plane as the humerus underwent a compound motion involving adduction, elevation, and rotation during propulsion.
The trochleariform distal humeral articular surface in modern mammals largely came about by widening the intercondylar groove and the development of a ridge within it (see Fig. 1-1, bear). The articular surface on the proximal ulna is oblique in orientation, and the distal half retains an articulation with the ulnar condyle. This spiral trochlear configuration allows the forearm to move in a sagittal plane while maintaining the stability of ulnohumeral contact through the cam effect of the ulnar condyle during humeral rotation.
COMPARATIVE PRIMATE ANATOMY OF THE ELBOW REGION
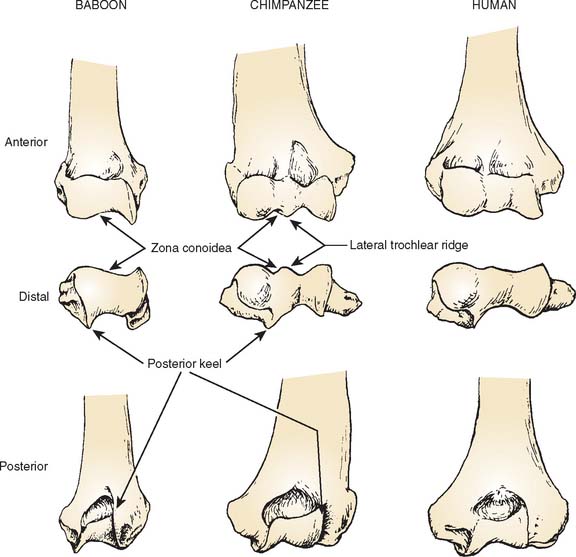
Much of what follows is taken from the detailed studies of Rose.20,21 The humeral trochlea may be cylindrical, conical, or trochleariform in nonhuman primates.21 The trochlea is conical in some prosimians, but a cylindrical trochlea seems to be the most common shape and is observed in most prosimians and New World monkeys. The trochlea is also cylindrical in most Old World monkeys but with a pronounced medial flange or keel that is best developed anterodistally (Fig. 1-2). Only in apes and humans is the trochlea truly trochleariform, possessing medial and lateral ridges all around the trochlear margins, which contribute to the stability of the ulnohumeral joint, substituting for the radiohumeral joint, which is freed for pronosupination throughout the flexion range.11,20 In most species, the articular surface of the trochlea expands posteriorly to the area behind the capitellum. In larger monkeys, the lateral edge of the posterior trochlear surface projects to form a keel that extends up the lateral wall of the olecranon fossa (see Fig. 1-2). In hominoids, this keel is a continuation of the lateral trochlear ridge and helps form a sharp lateral margin of the olecranon fossa, providing resistance to varus and internal rotation in extension.20.21
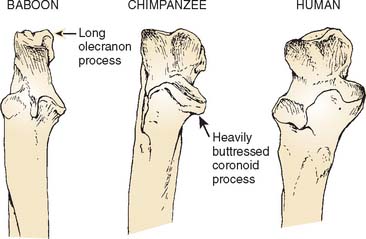
The trochlear notch of the ulna generally mirrors the shape of the humeral trochlea. In humans and apes, the notch has medial and lateral surfaces separated by a ridge that articulates with the trochlear groove (Fig. 1-3).20,21
The differences seen in the configuration of the humeroulnar joint across primate species reflect contrasting requirements for stabilization with different forms of limb use. In most monkeys, the humeroulnar joint is in its most stable configuration in a partially flexed position owing to the development of the medial trochlear keel anterodistally and the lateral keel posteriorly.20
The great apes (chimpanzees, gorillas, and orangutans) and the lesser apes (gibbons) move about in a much less stereotypical fashion than do monkeys. To accommodate this more varied form of limb use, the hominoid humeroulnar joint, with its deeply socketed articular surfaces and well-developed medial and lateral trochlear ridges all around the joint margins, is designed to provide maximum stability throughout the flexion-extension range.20–22 The use of overhead suspensory postures and locomotion in apes has led to the evolution of the capacity for complete elbow extension. Apes even keep their elbows extended during quadrupedal locomotion. The ideal joint configuration for resistance of transarticular stress with fully extended elbows during quadrupedal postures would be to have a trochlear notch that was proximally directed. It could then act as a cradle to support the humerus during locomotion. However, a proximal orientation of the trochlear notch would severely limit elbow flexion by impingement of the coronoid process within its fossa. The anteroproximal orientation of the trochlear notch in apes thus represents a compromise that safely supports the humerus on the ulna in extended elbow positions during locomotion without unduly sacrificing elbow flexion.1
On the lateral side of the elbow, the articular surface on the capitellum extends farther posteriorly in apes and humans than in monkeys, allowing the radius to move with the ulna into full extension of the elbow. In addition, the capitellum of apes and humans is uniformly rounded, reflecting versatility rather than stereotypy in forelimb usage (Fig. 1-4).
The gutter-like region between the trochlea and capitellum—the zona conoidea—is a relatively flat plane that terminates distally in most monkeys. In the hominoids, it continues posteriorly (see Fig. 1-1).20,21 The zona conoidea articulates with the bevel of the radial head, and differences in its configuration reflect differences in the shape of the radial head.
The radial head of hominoid primates is nearly circular, and the peripheral rim is symmetrical and beveled all around the circumference of the radial head for articulation with the zona conoidea (Fig. 1-5). This configuration provides good contact to resist dislocation of the radial head from the humerus under the varied loading regimes experienced by the hominoid elbow and can stabilize the radial head in all positions of pronosupination.20,21
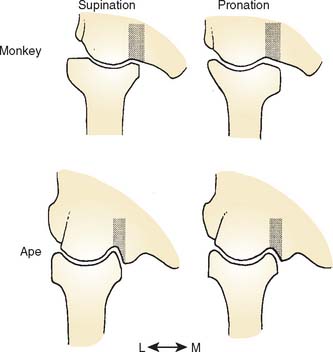
In most monkeys and prosimians, the radial head is ovoid and the proximal radioulnar joint articulation is restricted to the anterior and medial surfaces; as a result, the joint becomes close packed for stability in pronation (Fig. 1-6). In apes and humans, on the other hand, this articular surface extends almost all the way around the head, implying a greater range of pronosupination.20 The radial notch of the ulna in most monkeys and prosimians faces either anterolaterally or directly anteriorly, whereas in hominoids, it faces more laterally.20,21 The configuration observed in apes and humans emphasizes a broad range of pronosupination with a nearly equal degree of stability in all positions.20,21
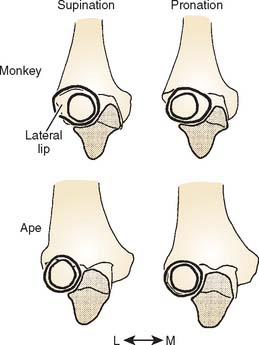
In general terms, most of the differences in elbow joint morphology between quadrupedal monkeys and the apes can be related to the development of a position of particular stability in monkeys versus more universal stability in apes.
A few additional features of the human elbow are shared with apes, such as a more distal biceps tuberosity (longer radial neck) relative to their body size.21 In apes, this is probably related to the demands for powerful elbow flexion to raise the center of mass of the body during climbing and suspensory postures and locomotion. Although the radial tuberosity faces more or less anteriorly in most primates, it faces more medially in apes and humans, reflecting their greater range of pronosupination.17 Extreme supination is an important component of suspensory locomotion in apes, and the medially placed tuberosity provides maximum supination torque near full supination.14,30 Apes and humans share a relatively short lever arm for triceps compared with that of most other primates, which is generally attributed to the demands for rapid elbow extension during suspensory locomotion. Finally, apes and humans are distinguished from other primate species in possessing a biomechanical carrying angle at the elbow. Sarmiento22 has argued that the evolution of a carrying angle in apes is related to the need to bring the center of mass of the body beneath the supporting hand during suspensory locomotion in a manner similar to that in which the valgus knee of humans brings the foot nearer the center of mass of the body during the single limb support phase of walking (Fig. 1-7).

The morphology of the modern human elbow is not identical to that of the ape elbow, however. In some cases, the differences are simply a matter of degree. For example, although both apes and humans are distinguished from other primates in the medial orientation of the radial tuberosity, it is more extreme in position in the ape; in the human it is typically slightly anterior to true medial. In addition, although the olecranon is short in both humans and apes compared with most monkeys, it is slightly longer in humans than in apes and also shaped to maintain this length throughout the range of flexion—both of which are advantageous for powerful manipulatory activities.4
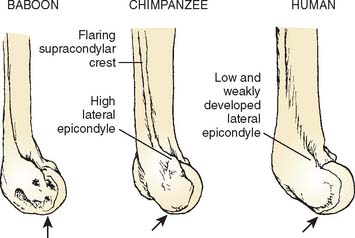
Other differences between the elbow morphology of humans and that of apes can be related to the fact that the human forelimb has no role in locomotion. These differences include a less robust coronoid process and a relatively narrower, proximally oriented trochlear notch in humans, indicating relative stability in flexion rather than the need to support the weight of the body during quadrupedal locomotion in extension.1,13 Humans possess a smaller and more distally placed lateral epicondyle and a less well-developed supracondylar crest than is seen in the apes, reflecting diminished leverage of the wrist extensors and brachioradialis.23–25 Humans have no bowing of the ulna that is related to enhancing the leverage of the forearm pronators and supinators in apes.1 Finally, a diminution in the prominence of the trochlear ridges and steep lateral margin of the olecranon fossa in humans can be related to the overall reduction in stresses at the human elbow and the concomitant relaxation on the demands for strong stabilization in all positions.20,21
FOSSIL EVIDENCE
Dendropithecus macinnesi, Limnopithecus legetet, and Proconsul heseloni (all from Africa) are among the earliest known hominoid species dated to the early part of the Miocene epoch (23 to 16 mya) for which postcranial material is known. Overall, the distal humeri of the first two of these forms resemble generalized New World monkeys such as Cebus (capuchin monkeys). The trochlea does not display a prominent lateral ridge, and the zona conoidea is relatively flat. The trochlear notch faces anteriorly, and the head of the radius is oval in outline with a well-developed lateral lip. These features generally are considered to be primitive for higher primates (monkeys, apes, and humans).8,9,20
P. heseloni, on the other hand, does display some features characteristic of extant hominoids. It has a globular capitellum, well-developed medial and lateral trochlear ridges, and a deep zona conoidea forming the medial wall of a recessed gutter between the capitellum and trochlea.20 In general, the elbow region of Proconsul resembles that of extant hominoids in features related to general stability and range of pronosupination; yet full pronation remained a position of particular stability.20
The limited fossil material that is available from the late Miocene epoch (16 to 5 mya) suggests that many hominoid species, including members of the genera Dryopithecus (from Europe), Sivapithecus (from Europe and Asia), and Oreopithecus (from Europe), displayed the features characteristic of the modern hominoid elbow. Although it is possible that these features arose in parallel in different genera, the more parsimonious explanation is that they inherited this morphology from an early to middle Miocene common ancestor, possibly similar to P. heseloni.16,29,31 Assuming that the characteristic features of the hominoid elbow are shared derived traits, that is, traits inherited from a single common ancestor, we can say that the elbow morphology of modern apes and humans can be dated to roughly 15 to 20 mya.
The majority of paleoanthropologists agree that humans are most closely related to the African apes (chimpanzees and gorillas) and that the two lineages arose in the late Miocene or earliest Pliocene period (between 10 and 4 mya).8 The earliest known fossils of the human lineage (hominids) date from the early Pliocene era, approximately 4 to 5 mya. There are three genera of these earliest hominids currently recognized, Ardipithecus, Paranthropus, and Australopithecus. The latter is the best known and most widespread genus, and includes the famous “Lucy” skeleton from Hadar, Ethiopia (A. afarensis).5,12 The genus Homo, to which our own species belongs, first appeared about 2.5 to 2 mya in East Africa with its earliest member species, H. habilis.
All of the hominids from the Pliocene period were bipedal, although some probably spent significant time climbing trees.23–2628 The development of bipedalism freed the upper extremity from the requirements of locomotion, placing greater emphasis on increasing mobility. The ability to supinate and pronate was an immense advantage to hominids in caring for their young, defending themselves, and gathering food. It was also critical in efficient tool handling, which developed approximately 2 mya, at about the same time as H. habilis, although there is debate about which species of early hominid was responsible for making them.27
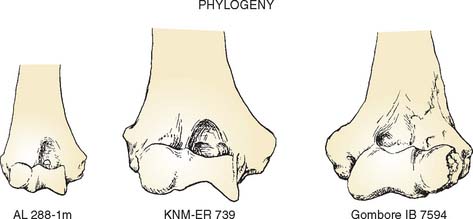
Several distal humeri are known from these early hominid species. All of the early hominid distal humeri lack the steep lateral margin of the olecranon fossa that is characteristic of chimpanzees and gorillas. However, they do show a considerable amount of morphologic variation in other characteristics (Fig. 1-8). On the basis of the contour of the distal end of the humeral shaft, the placement of the epicondyles, and the configuration of the articular surface, the fossil distal humeri have been divided into two groups. The first group is characterized by a weakly projecting lateral epicondyle that is placed low, at about the level of the capitellum, and by a moderately developed lateral trochlear ridge.23,24 These are features shared with modern humans, and consequently, this group generally is referred to as early Homo. The second group includes the Australopithecus and Paranthropus species and is characterized by a well-developed lateral epicondyle that is high relative to the capitellum. These features are similar to those of modern apes.
A number of fragments of early hominid proximal radii have been recovered representing each of the currently recognized species. The proximal radial fragments that have been attributed to early Homo display a much narrower bevel around the capitellar fovea than that of the modern apes and the earlier hominin group. This provides for articulation with a more shallow zona conoidea and a more vertical and uniformly wide surface on the side of the head for articulation with the ulna favoring pronosupination over stability. Other primitive hominoid features include thick cortices, a relatively long and angulated radial neck (lower neck shaft angle), and a more anteromedially (rather than medially) placed biceps tuberosity. Many of these features are still present in a small percentage of modern humans, limiting the functional conclusions that can be drawn and suggesting a mosaic pattern of evolution.18,19
Some early hominid ulnae that have been recovered appear to retain many primitive features including a longer more curved shaft, greater mediolateral width proximally, and a nonprominent interosseous border.1,2,10 However, early human ulnae attributed to the genus Homo are similar to those of modern humans in having a prominent interosseous border, a supinator crest, and a well-marked hollow for the play of the tuberosity of the radius.6,7,15 It appears, therefore, that many of the characteristics that distinguish the human elbow from that of the ape can be found in the earliest members of our genus.
In overview, the combination of comparative anatomy and the fossil record indicates that the modern human elbow owes its beginnings to our hominoid ancestry. Current evidence suggests that many of the characteristic features of the human distal humerus and proximal radius and ulna can be projected back approximately 15 to 20 mya to a common ancestor of extant apes and humans. Functional analysis suggests that this morphologic structure arose in hominoid primates in response to the need for stabilization throughout the flexion-extension and pronosupination ranges of motion to permit a more versatile form of forelimb use. This morphology was still largely intact following the evolution of upright posture and bipedal locomotion in the earliest known hominids. However, as the forelimb became less and less involved in locomotion, the hominid elbow underwent additional modifications, relaxing some of the emphasis on stabilization and increasing performance throughout the range of movement. The fossil record indicates that the distinct form of the modern human elbow probably first appeared about 2 mya in our ancestor H. habilis. This morphology has changed only subtly during all subsequent stages of human evolution.
1 Aiello L.C., Dean M.C. An Introduction to Human Evolutionary Anatomy. London: Academic Press, 1990.
2 Churchill S.E., Pearson O.M., Grine F.E., Trinkaus E., Holliday T.W. Morphological affinities of the proximal ulna from Klasies River main site: archaic or modern? J. Hum. Evol. 1996;31:213.
3 Conroy G.C. Primate Evolution. New York: W. W. Norton & Co., 1990.
4 Drapeau M.S. Functional anatomy of the olecranon process in hominoids and Plio-Pleistocene hominins. Am. J. Phys. Anthropol.. 2004;124:297.
5 Drapeau M.S., Ward C.V., Kimbel W.H., Johanson D.C., Rak Y. Associated cranial and forelimb remains attributed to Australopithecus afarensis from Hadar, Ethiopia. J. Hum. Evol.. 2005;48:593.
6 Day M.H. Functional interpretations of the morphology of postcranial remains of early African hominids. In: Jolly C.J., editor. Early Hominids of Africa. London: Duckworth; 1978:311.
7 Day M.H., Leakey R.E.F. New evidence for the genus Homo from East Rudolf, Kenya III. Am. J. Phys. Anthropol.. 1974;39:367.
8 Fleagle J.G. Primate Adaptation and Evolution, 2nd ed. New York: Academic Press, 1999.
9 Harrison T. The phylogenetic relationships of the early catarrhine primates: a review of the current evidence. J. Hum. Evol.. 1987;16:41.
10 Howell F.C., Wood B.A. Early hominid ulna from the Omo Basin, Ethiopia. Nature. 1974;249:174.
11 Jenkins F.A.Jr. The functional anatomy and evolution of the mammalian humeroulnar articulation. Am. J. Anat.. 1973;137:281.
12 Johanson D.C., Lovejoy C.O., Kimbel W.H., White T.D., Ward S.C., Bush M.E., Latimer B.M., Coppens Y. Morphology of the Pliocene partial hominid skeleton (A.L. 288-1) from the Hadar Formation, Ethiopia. Am. J. Phys. Anthropol.. 1982;57:403.
13 Knussmann R.. Humerus, Ulna and Radius der Simiae. Bibliotheca Primatologica, Vol. 5. S. Karger, Basel, 1967.
14 Larson S.G. Subscapularis function in gibbons and chimpanzees: implications for interpretation of humeral head torsion in hominoids. Am. J. Phys. Anthropol.. 1988;76:449.
15 Leakey R.E.F. Further evidence of lower Pleistocene hominids from East Rudolf, Northern Kenya. Nature. 1972;237:264.
16 Martin L., Andrews P.I. Cladistic relationships of extant and fossil hominoids. J. Hum. Evol.. 1987;16:101.
17 O’Connor B.L., Rarey K.E. Normal amplitudes of radioulnar pronation and supination in several genera of anthropoid primates. Am. J. Phys. Anthropol.. 1979;51:39.
18 Patel B.A. The hominoid proximal radius: re-interpreting locomotor behaviors in early hominins. J. Hum. Evol.. 2005;48:415.
19 Pearson O.M., Grine F.E. Re-analysis of the hominid radii from Cave of Hearths and Klasies River Mouth, South Africa. J. Hum. Evol.. 1997;32:577.
20 Rose M.D. Another look at the anthropoid elbow. J. Hum. Evol.. 1988;17:193.
21 Rose M.D. Functional anatomy of the elbow and forearm in primates. In: Gebo D., editor. Postcranial Adaptation in Nonhuman Primates. DeKalb, IL: Northern Illinois Press; 1993:70.
22 Sarmiento, E. E.: Functional Differences in the Skeleton of Wild and Captive Orang-Utans and Their Adaptive Significance. Ph.D. Thesis, New York University, 1985.
23 Senut B. Outlines of the distal humerus in hominoid primates: application to some Plio-Pleistocene hominids. In: Chiarelli A.B., Corruccini R., editors. Primate Evolutionary Biology. Berlin: Springer Verlag; 1981:81.
24 Senut B. Humeral outlines in some hominoid primates and in Plio-Pleistocene hominids. Am. J. Phys. Anthropol.. 1981;56:275.
25 Senut B., Tardieu C. Functional aspects of Plio-Pleistocene hominid limb bones: implications for taxonomy and phylogeny. In: Delson E., editor. Ancestors: The Hard Evidence. New York: A. Liss; 1985:193.
26 Stern J.T.Jr., Susman R.L. The locomotor anatomy of Australopithecus afarensis. Am. J. Phys. Anthropol.. 1983;60:279.
27 Susman R.L. Fossil evidence for early hominid tool use. Science. 1994;265:1570.
28 Susman R.L., Stern J.T.Jr., Jungers W.L. Arboreality and bipedality in Hadar hominids. Folia Primatol.. 1984;43:113.
29 Szalay F.S., Delson E. Evolutionary History of the Primates. New York: Academic Press, 1979.
30 Trinkaus E., Churchill S.E. Neandertal radial tuberosity orientation. Am. J. Phys. Anthropol.. 1988;75:15.
31 Ward C.V., Walker A., Teaford M.F. Proconsul did not have a tail. J. Hum. Evol.. 1991;21:215.