Parasitic infections
PROTOZOAL INFECTIONS

AMEBIC INFECTIONS
CEREBRAL AMEBIC ABSCESS
 PATHOGENESIS OF CEREBRAL AMEBIC ABSCESS
PATHOGENESIS OF CEREBRAL AMEBIC ABSCESS



 CEREBRAL AMEBIC ABSCESS
CEREBRAL AMEBIC ABSCESS


MICROSCOPIC APPEARANCES
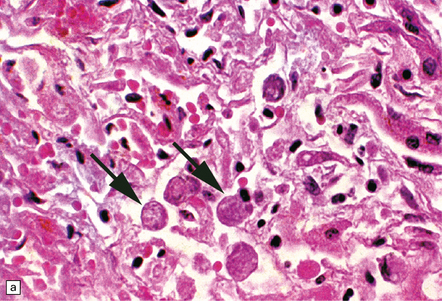
Cerebral amebic abscesses contain inflamed necrotic tissue and it may be difficult to distinguish E. histolytica trophozoites within this tissue from macrophages. The trophozoites are spherical or oval, 15–25 μm in diameter, and have vacuolated cytoplasm and a single nucleus. Occasionally pseudopodia can be seen. In routinely stained sections, the nuclei are round and have a small central karyosome and peripheral chromatin (Fig. 18.1). The cytoplasm contains abundant glycogen.
PRIMARY AMEBIC MENINGOENCEPHALITIS
 PATHOGENESIS OF PRIMARY AMEBIC MENINGOENCEPHALITIS
PATHOGENESIS OF PRIMARY AMEBIC MENINGOENCEPHALITIS



 PRIMARY AMEBIC MENINGOENCEPHALITIS
PRIMARY AMEBIC MENINGOENCEPHALITIS

MACROSCOPIC APPEARANCES
The brain is swollen and there is a hemorrhagic exudate in the meninges over the cerebrum, brain stem, and cerebellum. The olfactory bulbs and tracts and the adjacent parts of the frontal and temporal lobes contain areas of hemorrhagic necrosis (Fig. 18.2).
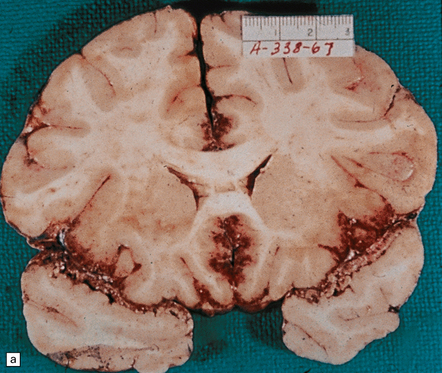
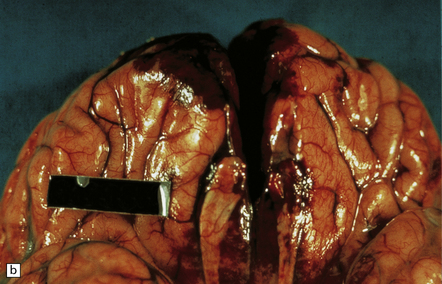
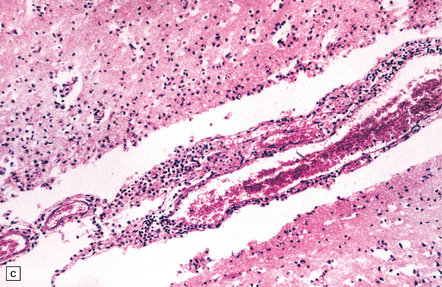
18.2 Primary amebic meningoencephalitis. (a) A hemorrhagic exudate is present in the meninges. There is also hemorrhagic necrosis of the underlying temporal, inferior frontal, and cingulate cortex. (Courtesy of Dr A J Martinez, University of Pittsburgh, USA.) (b) Hemorrhagic necrosis of the frontal poles, olfactory bulbs and tracts, and the underlying inferior frontal cortex. (Courtesy of Dr A J Martinez, University of Pittsburgh, USA.) (c) Scanty inflammatory infiltrates are present in the meninges.
MICROSCOPIC APPEARANCES
A scanty mononuclear inflammatory infiltrate (Fig. 18.2) with focal hemorrhage is seen in the meninges, and there is usually extensive necrosis of brain parenchyma. N. fowleri amebae are present in the subarachnoid space and around vessels in the necrotic parenchyma (Fig. 18.3). Their diameter, of 8–15 μm, is slightly less than that of E. histolytica. They resemble macrophages, but can be distinguished from them by their vesicular nucleus with its large central nucleolus.
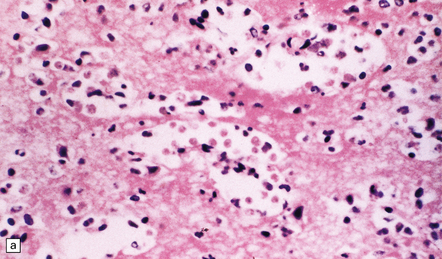
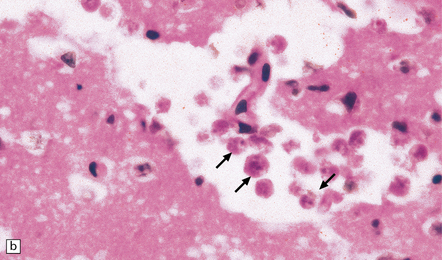
GRANULOMATOUS AMEBIC ENCEPHALITIS
 PATHOGENESIS OF GRANULOMATOUS AMEBIC ENCEPHALITIS
PATHOGENESIS OF GRANULOMATOUS AMEBIC ENCEPHALITIS



 GRANULOMATOUS AMEBIC ENCEPHALITIS
GRANULOMATOUS AMEBIC ENCEPHALITIS

MACROSCOPIC APPEARANCES
The brain is usually swollen, covered by a diffuse leptomeningeal exudate, and includes foci of softening, hemorrhage, and necrosis, particularly in the anterior part of the cerebral hemispheres, the thalamus (Fig. 18.4), brain stem, and cerebellum (Fig. 18.5).
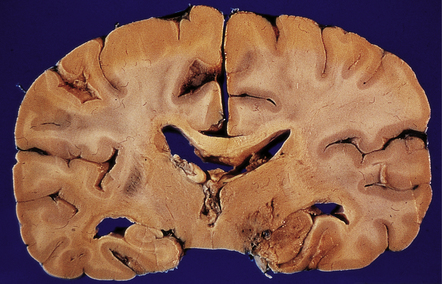
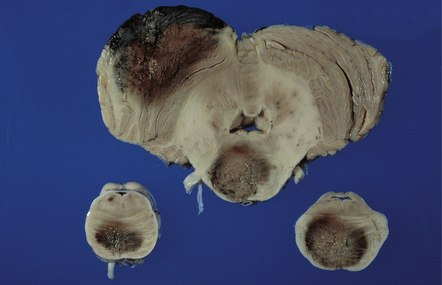
MICROSCOPIC APPEARANCES
The brain shows foci of chronic inflammation centered around arteries and veins. The inflammation is typically granulomatous and includes lymphocytes, macrophages, plasma cells, and multinucleated giant cells, but may be necrotizing. There is also a chronic inflammatory infiltrate in the meninges (Fig. 18.6). Acanthamoeba or Balamuthia trophozoites and cysts may be found in and around the walls of affected blood vessels (Fig. 18.6), and also in areas relatively free of inflammation (Fig. 18.6). The amebae are 15–40 μm in diameter and have a prominent vesicular nucleus with a dense central nucleolus. The cysts are surrounded by a double membrane (Fig. 18.6).
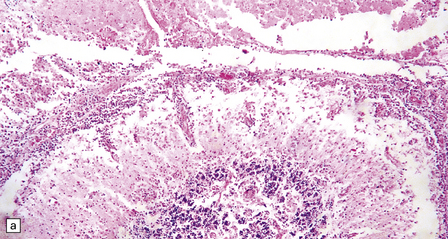
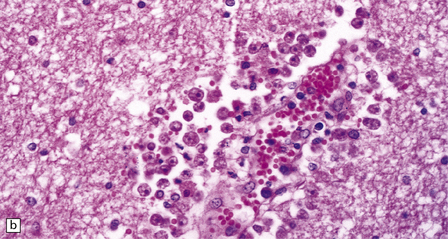
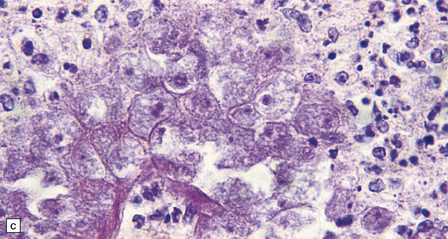
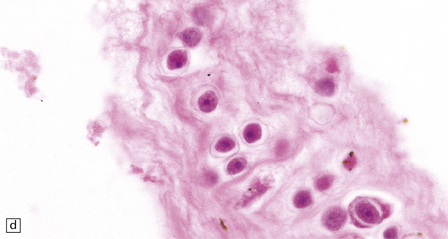
18.6 Granulomatous amebic encephalitis. (a) Meningeal and perivascular inflammation with necrosis of adjacent cerebellar tissue. (b) Perivascular Acanthameba trophozoites and scanty mononuclear inflammatory cells in the cerebral white matter. The amebae have a prominent vesicular nucleus with a dense central nucleolus. (c) This section shows perivascular Balamuthia trophozoites and cysts with surrounding inflammation and necrosis. (d) Encysted amebae in the wall of a leptomeningeal blood vessel.
CEREBRAL MALARIA
 CEREBRAL MALARIA
CEREBRAL MALARIA





MACROSCOPIC APPEARANCES
The brain is usually swollen with congested leptomeninges. The cerebral cortex may appear dusky pink color due to marked congestion, or slate gray due to the presence of abundant malarial pigment (Fig. 18.8). The white matter often contains petechial hemorrhages.
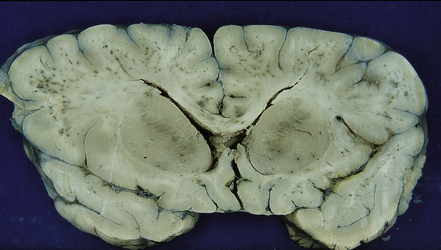
18.8 Slate gray brain in cerebral malaria. Blood vessels in the white matter are congested and some are surrounded by petechial hemorrhages.
 PATHOGENESIS OF CEREBRAL MALARIA
PATHOGENESIS OF CEREBRAL MALARIA
 Malaria is caused by four species of Plasmodium: P. falciparum, P. vivax, P. malariae, and P. ovale, the first two being responsible for 95% of cases.
Malaria is caused by four species of Plasmodium: P. falciparum, P. vivax, P. malariae, and P. ovale, the first two being responsible for 95% of cases.

 Sickle cell trait or disease provides some protection.
Sickle cell trait or disease provides some protection.
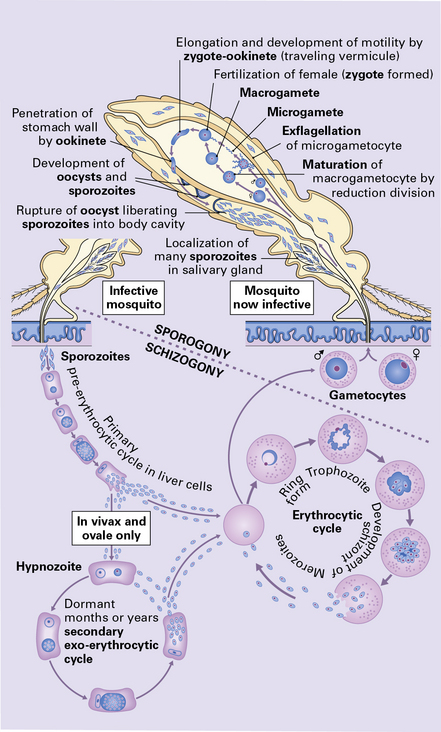
 Malaria is acquired by the bite of an infected Anopheles mosquito, which inoculates the sporozoites into the blood stream. The sporozoites are then carried to the liver, where they penetrate hepatocytes and develop into merozoites, which rupture the hepatocytes, enter the blood stream, and invade red blood cells (Fig. 18.7). The merozoites develop into trophozoites, which subsequently produce schizonts. After maturation, the schizont splits into merozoites. The red blood cells rupture liberating the merozoites which enter other red blood cells to repeat the schizogonic cycle.
Malaria is acquired by the bite of an infected Anopheles mosquito, which inoculates the sporozoites into the blood stream. The sporozoites are then carried to the liver, where they penetrate hepatocytes and develop into merozoites, which rupture the hepatocytes, enter the blood stream, and invade red blood cells (Fig. 18.7). The merozoites develop into trophozoites, which subsequently produce schizonts. After maturation, the schizont splits into merozoites. The red blood cells rupture liberating the merozoites which enter other red blood cells to repeat the schizogonic cycle.




MICROSCOPIC APPEARANCES
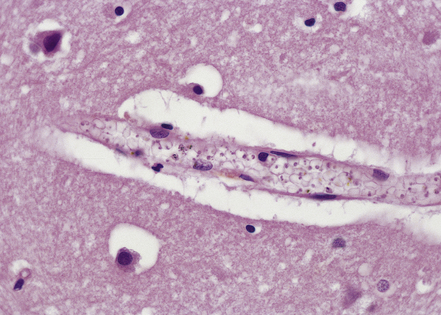
The small vessels are engorged by red blood cells, which may have a ghost-like appearance with poor staining of hemoglobin (Fig. 18.9). Many of these cells, particularly in the gray matter, contain malaria parasites and/or granules of dark malarial pigment, which is related to hematin. Marginated aggregates of red blood cells may appear to be adherent to the vascular endothelium (Fig. 18.10).
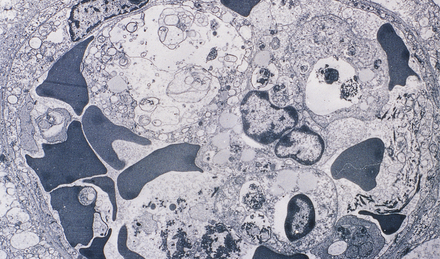
18.10 Endothelial adherence of parasitized red blood cells.
Electron micrograph showing a capillary occluded by macrophages with osmiophilic granular pigment and red blood cells containing P. falciparum schizonts. Note that the parasitized red blood cells have knob-like protrusions that are attached to the endothelium. (Courtesy of Dr Carmen L P Lancellotti, Department of Pathology, Santa Casa de Sao Paulo, Brazil.)
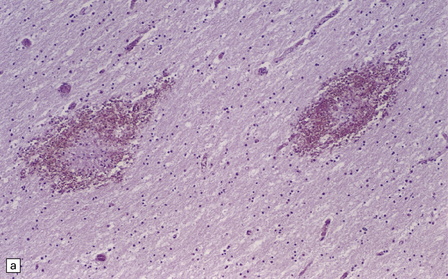
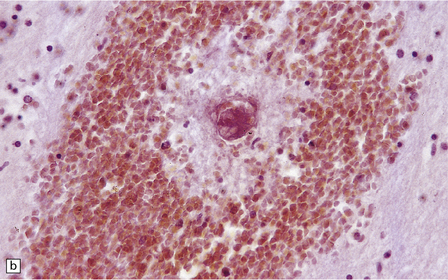
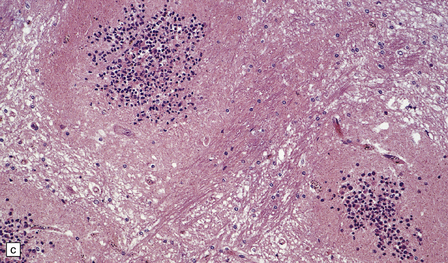
Edema, capillary necrosis, and perivascular hemorrhages are usually evident, and there may be parenchymal and meningeal infiltration by lymphocytes and macrophages. Petechial or larger hemorrhages can occur in any part of the brain, but are most common in the white matter and may surround necrotic arterioles and veins (Fig. 18.11). Patients with longer survival may harbor foci of softening and gliosis. Collections of microglia and astrocytes, the so-called Dürck granulomas, are probably related to resorption of ring hemorrhages (Fig. 18.11). The microglia contain iron pigment and lipid.
CEREBRAL TOXOPLASMOSIS
POSTNATALLY-ACQUIRED CEREBRAL TOXOPLASMOSIS
 CONGENITAL TOXOPLASMOSIS
CONGENITAL TOXOPLASMOSIS



MACROSCOPIC APPEARANCES
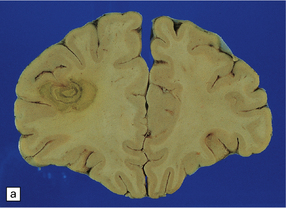
The brain typically contains multifocal necrotic lesions of variable size (Fig. 18.12). There may be associated hemorrhage. Older lesions are cystic due to resorption of necrotic material. The basal ganglia are often involved, but any part of the brain may be affected. Occasionally brain involvement results in an encephalitic process without obvious focal lesions on macroscopic examination.
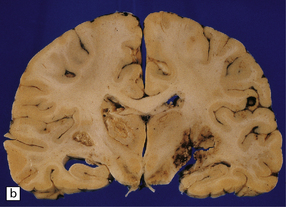
MICROSCOPIC APPEARANCES
Necrotizing abscesses or foci of coagulative necrosis are surrounded by mononuclear and polymorphonuclear inflammatory cells, newly formed capillaries, reactive astrocytes, and microglia (Fig. 18.13). Infiltrates of lymphocytes and macrophages surround the blood vessels. Scanty fibrous encapsulation may be evident. Other findings include intimal proliferation and thrombosis, fibrinoid necrosis (Fig. 18.13), and perivascular hemorrhage. The pathological findings depend partly on the degree of impairment of immune function: inflammation is less prominent and fibrosis usually absent in patients whose immune function is severely compromised.
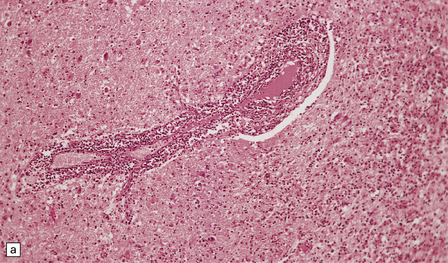
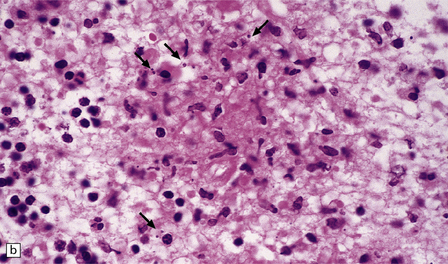
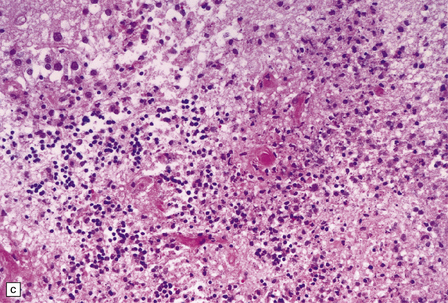
18.13 Histologic features of toxoplasmosis. (a) Inflammation adjacent to a Toxoplasma abscess. A dense perivascular and interstitial infiltrate of inflammatory cells, predominantly lymphocytes and macrophages, is present in the adjacent brain tissue. (b) Toxoplasmosis involving the cerebellar cortex. Note the proliferation of microglia and the presence of extracellular Toxoplasma tachyzoites (arrows). (c) Fibrinoid necrosis of blood vessels in the cerebellum.
 PATHOGENESIS OF CEREBRAL TOXOPLASMOSIS
PATHOGENESIS OF CEREBRAL TOXOPLASMOSIS






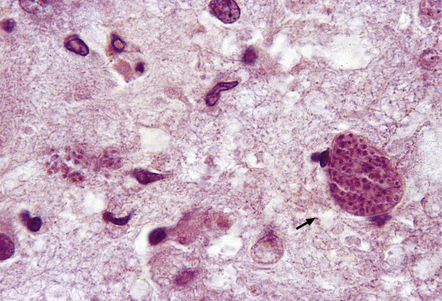
Intracellular and extracellular Toxoplasma tachyzoites (also known as endozoites or trophozoites) are usually abundant. They are oval- or crescent-shaped and measure 2–4 μm by 4–8 μm (Fig. 18.14). Those within cells may be clustered together (in vacuoles or larger pseudocysts) or may appear to lie free in the cell cytoplasm. They can be seen reasonably well when stained with hematoxylin and eosin, but are more readily identified and distinguished from other protozoa immunohistochemically (Fig. 18.15). Cysts measuring 20–100 μm in diameter and containing large numbers of bradyzoites (also known as cystozoites) may occur within, or at the periphery of, the necrotic areas (Figs 18.15, 18.16).
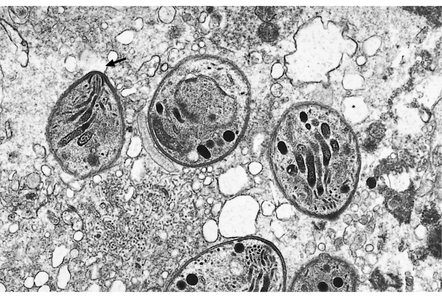
18.14 Ultrastructural appearance of Toxoplasma tachyzoites.
This electron micrograph shows several Toxoplasma tachyzoites, each surrounded by a double-layered pellicle. The conoid of one of the tachyzoites is clearly visible (arrow).
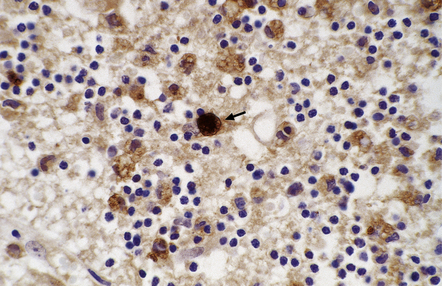
18.15 Immunohistochemical demonstration of Toxoplasma. Immunostained Toxoplasma protozoa are visible within a cyst (arrow) and in the cytoplasm of macrophages.
CONGENITAL TOXOPLASMOSIS
 CEREBRAL TOXOPLASMOSIS
CEREBRAL TOXOPLASMOSIS The clinical presentation of symptomatic disease of the CNS is variable and may include headaches, drowsiness, disorientation, non-specific features of raised intracranial pressure, and progressive focal neurologic deficits.
The clinical presentation of symptomatic disease of the CNS is variable and may include headaches, drowsiness, disorientation, non-specific features of raised intracranial pressure, and progressive focal neurologic deficits.
 CT scans and magnetic resonance imaging usually reveal multiple ring-enhancing lesions.
CT scans and magnetic resonance imaging usually reveal multiple ring-enhancing lesions.
MACROSCOPIC APPEARANCES
Macroscopic abnormalities occur in the more severe cases and include:
 Multifocal or confluent areas of necrosis, particularly in the periventricular and subpial regions (Fig. 18.17).
Multifocal or confluent areas of necrosis, particularly in the periventricular and subpial regions (Fig. 18.17).
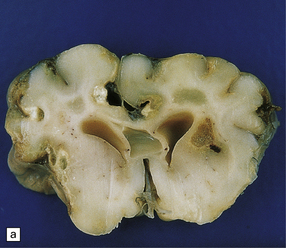
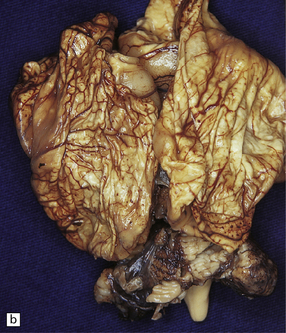
18.17 Congenital toxoplasmosis. (a) Foci of subpial and deep white matter necrosis are seen in this case of congenital toxoplasmosis. Some of the subpial foci are partly calcified. (b) Collapsed cerebral hemispheres of a severely hydrocephalic brain in congenital toxoplasmosis.
 Hydrocephalus or even hydranencephaly (Fig. 18.17). The brain may collapse after removal from the cranial cavity, because of extensive parenchymal destruction secondary to vascular thrombosis.
Hydrocephalus or even hydranencephaly (Fig. 18.17). The brain may collapse after removal from the cranial cavity, because of extensive parenchymal destruction secondary to vascular thrombosis.
 Foci of calcification scattered throughout the brain (Fig. 18.17) in contrast to the predominantly periventricular calcification of congenital cytomegalovirus encephalitis
Foci of calcification scattered throughout the brain (Fig. 18.17) in contrast to the predominantly periventricular calcification of congenital cytomegalovirus encephalitis

 PATHOGENESIS OF CONGENITAL TOXOPLASMOSIS
PATHOGENESIS OF CONGENITAL TOXOPLASMOSIS

MICROSCOPIC APPEARANCES
Active lesions in congenital toxoplasmosis show extensive coagulative necrosis. They are usually associated with lipid-laden macrophages, lymphocytes, and a few neutrophils, which are also present in the leptomeninges (Fig. 18.18). The adjacent brain tissue may contain microglial nodules. Toxoplasma tachyzoites and cysts can be seen in the meningeal exudate, around (rather than within) the necrotic lesions, and are particularly numerous near the ventricular cavities (Fig. 18.18).
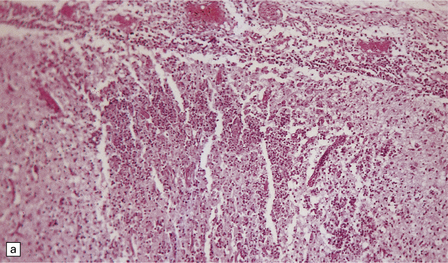
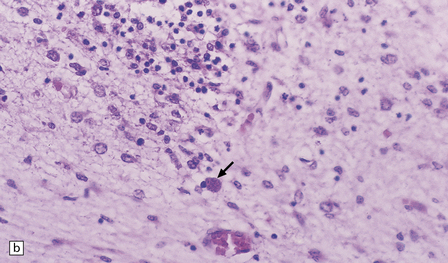
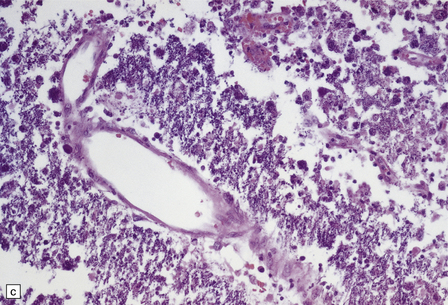
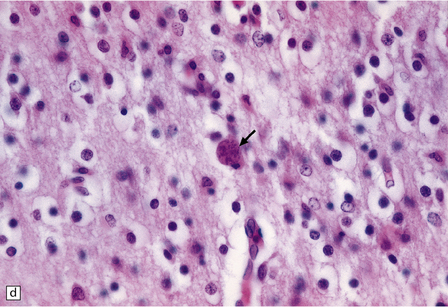
18.18 Congenital toxoplasmosis. (a) Focal cortical necrosis with inflammation of the adjacent brain tissue and overlying leptomeninges. (b) A cyst (arrow) and some lymphocytes are present at the periphery of a necrotic lesion. (c) The foci of necrosis tend eventually to undergo mineralization, as illustrated here. (d) Residual Toxoplasma cyst (arrow).
The foci of necrosis eventually tend to undergo mineralization (Fig. 18.18, see also Fig. 18.17). Residual Toxoplasma cysts can usually be found (Fig. 18.18). Ependymal granulations and gliosis may lead to aqueduct stenosis and obstructive hydrocephalus.
TRYPANOSOMIASIS
AFRICAN TRYPANOSOMIASIS (SLEEPING SICKNESS)
 AFRICAN TRYPANOSOMIASIS
AFRICAN TRYPANOSOMIASIS

MACROSCOPIC APPEARANCES
The leptomeninges may be cloudy and the brain swollen and congested. Sometimes the brain is macroscopically normal. Patients who have been treated with melarsoprol may develop acute hemorrhagic leukoencephalopathy (Fig. 18.19) (see Chapter 20).
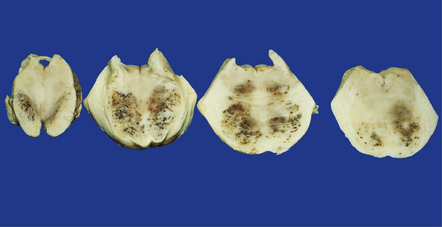
18.19 Acute hemorrhagic leukoencephalopathy may complicate melarsoprol treatment. Petechial hemorrhages and surrounding gray discoloration in the brain stem of a patient with acute hemorrhagic leukoencephalopathy. This was due to melarsoprol treatment of trypanosomiasis. (Courtesy of Professor J H Adams, Southern General Hospital, Glasgow.)
 PATHOGENESIS OF AFRICAN TRYPANOSOMIASIS
PATHOGENESIS OF AFRICAN TRYPANOSOMIASIS



MICROSCOPIC APPEARANCES
Lymphocytes, macrophages, and plasma cells surround the cerebral blood vessels and infiltrate the subarachnoid space (Fig. 18.20). Some of the plasma cells contain multiple intracytoplasmic globules of immunoglobulin. These ‘morular cells’ (Fig. 18.20) are characteris tic, but not specific. Other features include a reactive gliosis, and clusters of mononuclear inflammatory cells (Fig. 18.20). Typical microglial nodules may be abundant, especially in T. b. rhodesiense encephalitis, and are often associated with lymphophagocytosis. Trypanosomes are rarely demonstrable in the histologic sections.
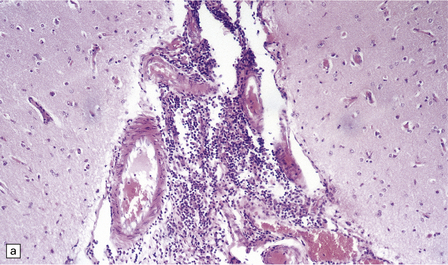
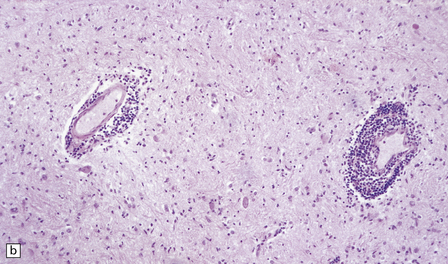
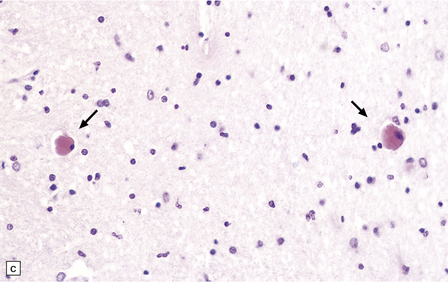
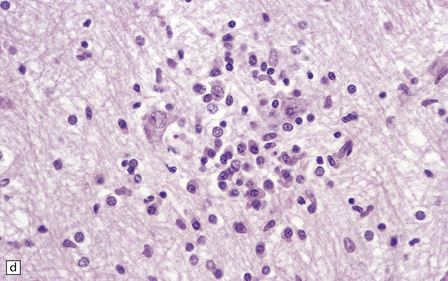
18.20 African trypanosomiasis. (a) Meningeal and (b) perivascular parenchymal infiltrates of mononuclear inflammatory cells in trypanosomiasis. Note the diffuse reactive gliosis. (c) Scattered morular cells (arrows) are characteristic. (d) Parenchymal aggregate of mononuclear inflammatory cells. (Courtesy of Professor J H Adams, Southern General Hospital, Glasgow.)
AMERICAN TRYPANOSOMIASIS (CHAGAS’ DISEASE)
 PATHOGENESIS OF AMERICAN TRYPANOSOMIASIS
PATHOGENESIS OF AMERICAN TRYPANOSOMIASIS


 AMERICAN TRYPANOSOMIASIS
AMERICAN TRYPANOSOMIASIS






MACROSCOPIC AND MICROSCOPIC APPEARANCES
Acute phase: The brain appears swollen and congested, with scattered petechial hemorrhages. Amastigote (Leishmania-like) forms of the parasites are present within glial cells (Fig. 18.21) or less frequently at the center of microglial nodules, which are scattered within the brain parenchyma (Fig. 18.21).
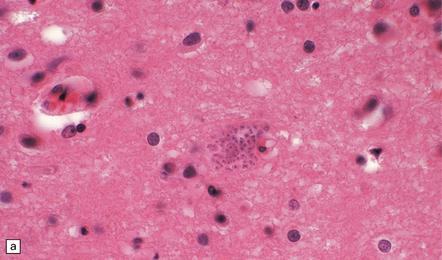
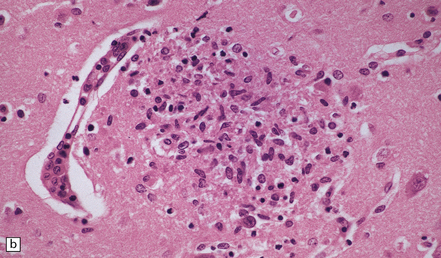
Reactivated disease takes the form of granulomatous or multifocal necrotizing encephalitis (Figs 18.22, 18.23) with abundant amastigote parasites (Fig. 18.23). The identity of the parasite can be confirmed immunohistochemically.
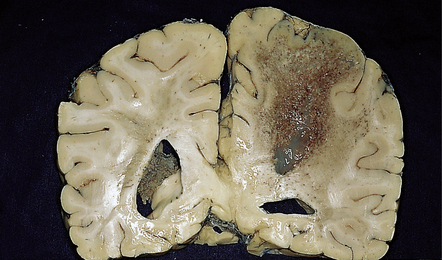
18.22 Necrotizing T. cruzi infection. Large necrotic lesion in the brain of a patient with AIDS and T. cruzi encephalitis.
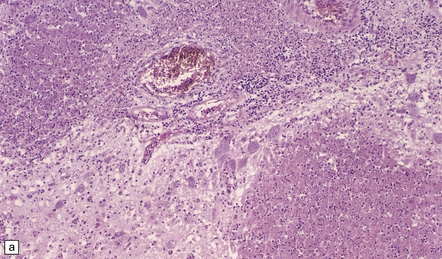
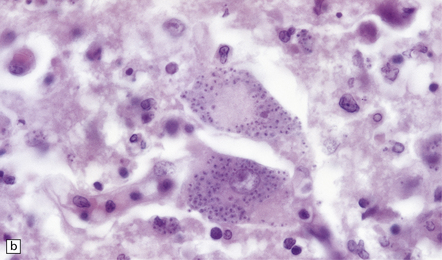
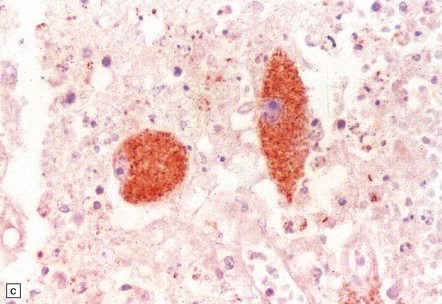
18.23 T. cruzi multifocal necrotizing encephalitis in AIDS. (a) This pattern of infection with necrosis is restricted to immunocompromised patients. (b) Abundant amastigote parasites, most of which appear to be in astrocytes and macrophages. (c) Immunohistochemical confirmation of the identity of T. cruzi parasites.






HELMINTHIC INFECTIONS
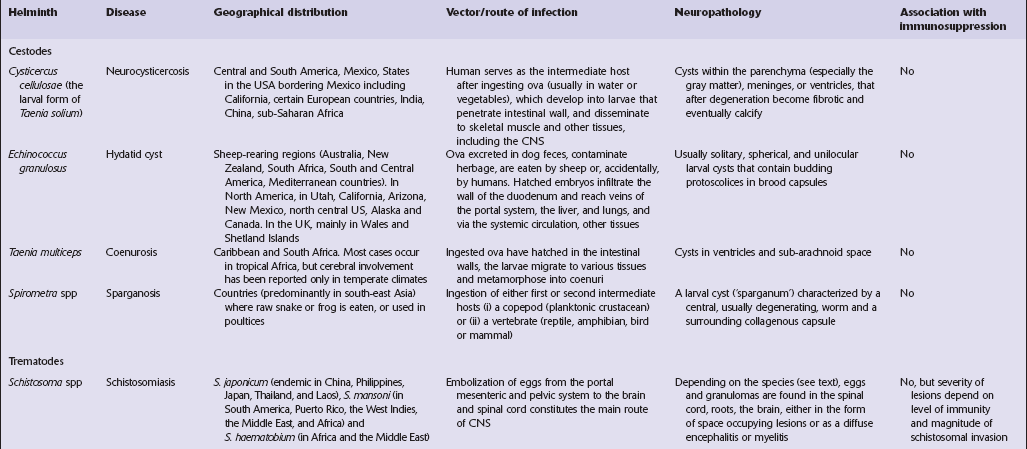
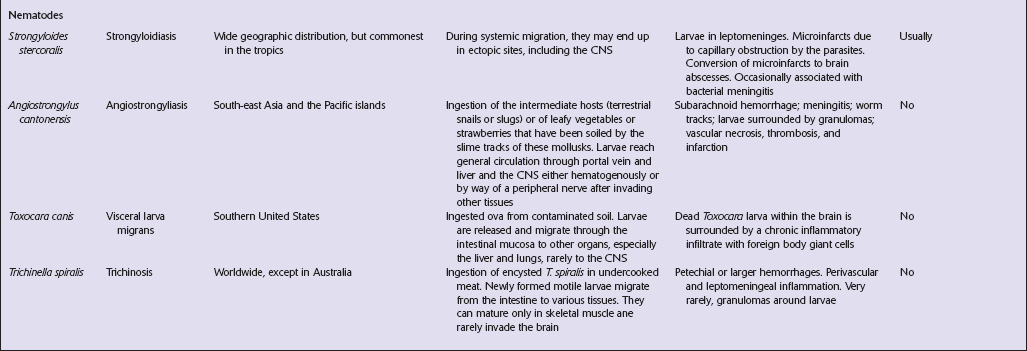
CYSTICERCOSIS
 CNS CYSTICERCOSIS
CNS CYSTICERCOSIS


MACROSCOPIC APPEARANCES
The number of cysts within the CNS varies from one to several hundred (Fig. 18.25). They occur in the parenchyma (especially the gray matter), meninges, or ventricles (Fig. 18.25). The viable intraparenchymal cysticerci are usually 1–2 cm in diameter and contain a single invaginated scolex. After degeneration they become fibrotic and represented by a firm white nodule (Fig. 18.25), which eventually calcifies. The spinal cord is rarely involved.
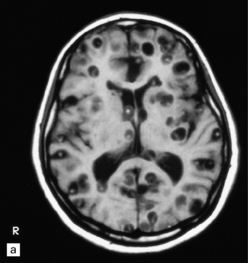
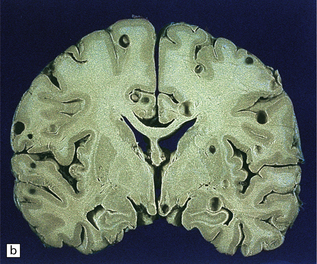
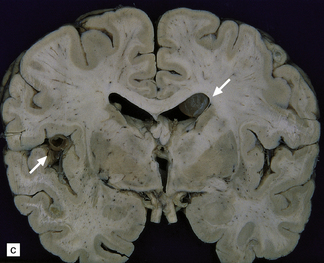
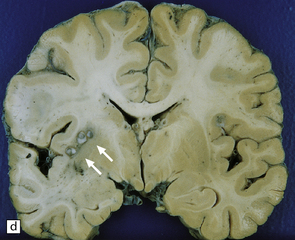
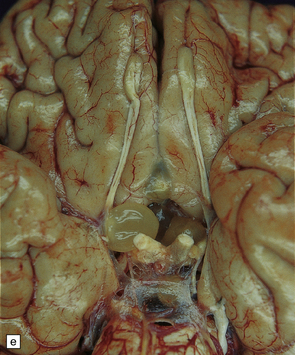
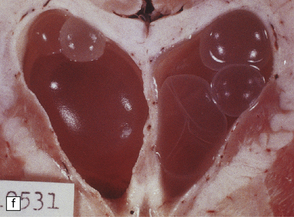
18.25 Cysticercosis. (a) Axial magnetic resonance image of the brain of a patient with cysticercosis. There are multiple visible cysticerci (low signal intensity lesions) and degenerating parasites (producing cysts with higher signal intensity). The small foci of high signal intensity in several of the cysts are caused by the scolex. (Courtesy of Dr B O Colli, Division of Neurosurgery, School of Medicine of Ribeirão Preto, University of São Paulo, Brazil.) (b) This coronal brain slice contains multiple cysts in the cortex and white matter, some of them with a scolex. (Courtesy of Dr J E H Pittella, Department of Pathology, School of Medicine, Federal University of Minas Gerais, Brazil.) (c) Intraventricular and leptomeningeal cysticerci (arrows). (d) Degenerate cysticerci in the brain parenchyma. After degeneration, the cysticerci become firm white or gray nodules (arrows) that are partly calcified. (e) Cysticerci in the leptomeninges anterior to the optic chiasm. (f) Racemose cysticerci. The cysticerci are forming grape-like clusters in the lateral ventricles. (Courtesy of Dr J E H Pittella, Department of Pathology, School of Medicine, Federal University of Minas Gerais, Brazil.)
The meningeal cysts are small and colorless. They adhere to the pia or float freely in the subarachnoid space (Fig. 18.25), particularly in the sylvian fissure. With time, basal cysts tend to shrink, and the meninges become thickened and fibrotic.
Racemose cysticerci are large multiloculated grape-like clusters of cysts that lack an invaginated scolex. They are usually found in the basilar cisterns or within the ventricular system (Fig. 18.25), especially the fourth ventricle.
 PATHOGENESIS OF CYSTICERCOSIS
PATHOGENESIS OF CYSTICERCOSIS The etiologic agent of cysticercosis is Cysticercus cellusosae, the larval form of the pork tapeworm, Taenia solium.
The etiologic agent of cysticercosis is Cysticercus cellusosae, the larval form of the pork tapeworm, Taenia solium.

 Proglottids are ingested by a suitable intermediate host, usually a pig. The ova then develop into larvae that penetrate the intestinal wall, invade the lymphatics and veins, and disseminate to skeletal muscle and other tissues, including the CNS (Fig. 18.24).
Proglottids are ingested by a suitable intermediate host, usually a pig. The ova then develop into larvae that penetrate the intestinal wall, invade the lymphatics and veins, and disseminate to skeletal muscle and other tissues, including the CNS (Fig. 18.24).
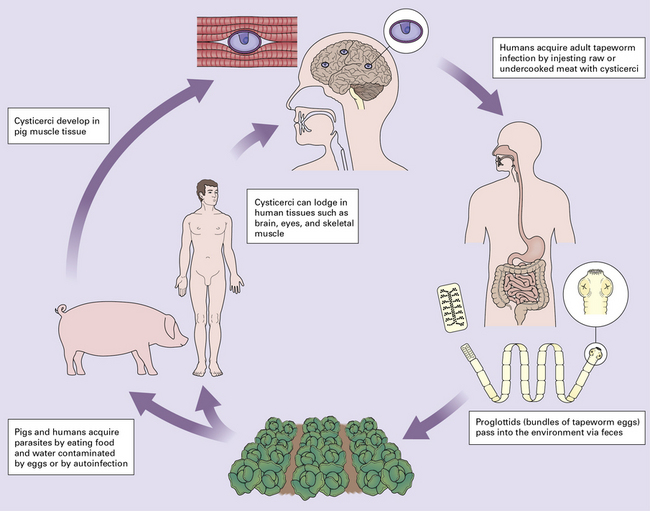
 Cysticercosis occurs when a human serves as the intermediate host.
Cysticercosis occurs when a human serves as the intermediate host.
Human acquisition of the larval forms that produce cysticercosis occurs:
MICROSCOPIC APPEARANCES
Each scolex has a rostellum with four suckers and a double row of 22–32 hooklets (Fig. 18.26). The cyst wall is sparsely cellular and consists of three histologically distinct layers (Fig. 18.26):
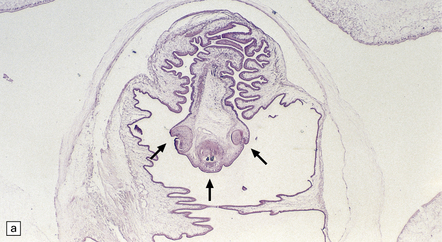
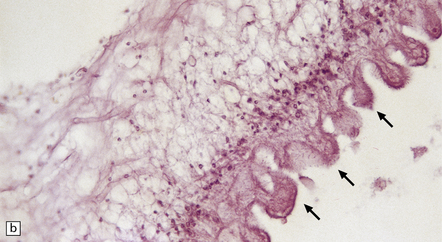
18.26 T. solium. (a) Scolex. The rostellum, with suckers (arrows), is clearly visible. (b) Section through T. solium cyst wall. This consists of a cuticular layer with hair-like protrusions (microtrichia) (arrows), a middle cellular layer, and an inner reticular layer.
 An outer or cuticular layer, which is about 3 μm thick, is eosinophilic, and has hair-like protrusions called microtrichia.
An outer or cuticular layer, which is about 3 μm thick, is eosinophilic, and has hair-like protrusions called microtrichia.

 An inner reticular or fibrillary layer, which may contain fine mineral concretions.
An inner reticular or fibrillary layer, which may contain fine mineral concretions.
While encysted larvae are viable, the surrounding parenchyma shows minimal reaction. Shortly after the cysticerci die, they are surrounded by neutrophils, lymphocytes, macrophages, foreign body giant cells, and eosinophils (Fig. 18.27), and then enclosed by a zone of granulation tissue, which eventually produces a dense collagenous capsule (Fig. 18.27). The lesion may include necrotic tissue and cholesterol clefts. Old nodules may be entirely fibrotic. Eventually some of the cysts become mineralized (Fig. 18.27).
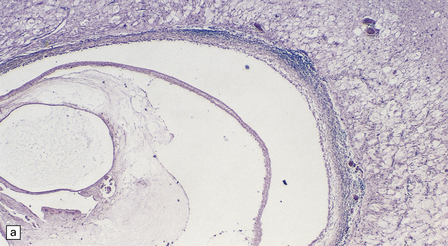
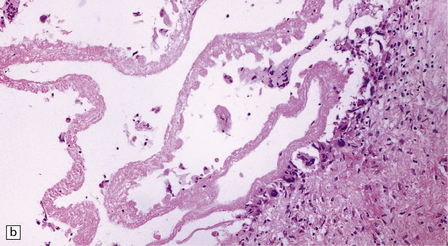
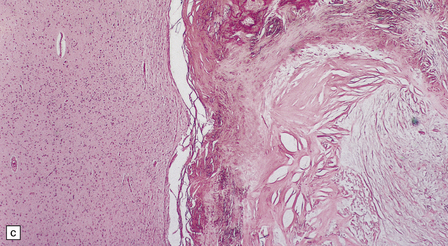
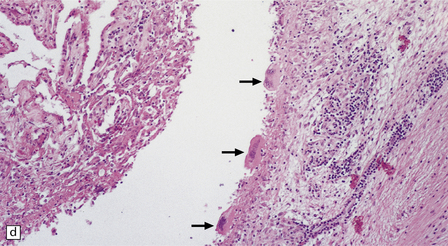
18.27 Dead cysticercus. (a) An intense inflammatory reaction is present around the dead parasite. (b) The necrotic cyst wall is abutted by a row of foreign body giant cells. There are scattered lymphocytes and microglia in the surrounding brain tissue. (c) In this case, the dead parasite is enclosed in a dense, focally calcified, collagenous capsule. (d) Degeneration of ventricular cysticerci has provoked florid granulomatous inflammation in the choroid plexus and periventricular tissue. Multinucleated giant cells (arrows) are present in the wall of the ventricle.
Ventricular cysticerci usually cause a granular ependymitis. Degeneration of racemose cysticerci in the ventricles or subarachnoid space may provoke a florid granulomatous inflammatory reaction (Fig. 18.27).
HYDATID DISEASE (ECHINOCOCCOSIS)
 HYDATIDOSIS
HYDATIDOSIS



MACROSCOPIC APPEARANCES
 PATHOGENESIS OF HYDATIDOSIS
PATHOGENESIS OF HYDATIDOSIS Hydatid disease is usually caused by larvae of Echinococcus granulosus, much less commonly E. multilocularis or E. vogeli. These are small tapeworms found primarily in dogs, coyotes, wolves, dingoes and jackals. The usual intermediate hosts are sheep. The disease occurs in sheep-rearing regions, especially in Australia, New Zealand, South Africa, South and Central America, and several Mediterranean countries. In North America, it occurs in sheep-raising regions of Utah, California, Arizona, New Mexico, north central US, Alaska and Canada. In the UK, hydatid disease occurs mainly in Wales and the Shetland Islands.
Hydatid disease is usually caused by larvae of Echinococcus granulosus, much less commonly E. multilocularis or E. vogeli. These are small tapeworms found primarily in dogs, coyotes, wolves, dingoes and jackals. The usual intermediate hosts are sheep. The disease occurs in sheep-rearing regions, especially in Australia, New Zealand, South Africa, South and Central America, and several Mediterranean countries. In North America, it occurs in sheep-raising regions of Utah, California, Arizona, New Mexico, north central US, Alaska and Canada. In the UK, hydatid disease occurs mainly in Wales and the Shetland Islands.


MICROSCOPIC APPEARANCES
The wall of the cyst (Fig. 18.28) consists of:
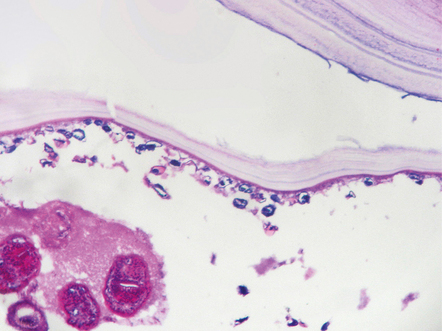
18.28 Hydatid (echinococcal) cyst. Section through the edge of a cyst, showing part of the cyst wall and an adjacent brood capsule within which are several protoscolices. The irregularly shaped dark blue fragments of calcified material just beneath the cyst wall are known as calcaneus corpuscles.
 An outer layer of fibrous tissue, derived from the host.
An outer layer of fibrous tissue, derived from the host.
 An intermediate laminated, chitinous or cuticular layer.
An intermediate laminated, chitinous or cuticular layer.
 A thin inner germinal layer to which oval brood capsules are usually attached.
A thin inner germinal layer to which oval brood capsules are usually attached.
The interior of the cyst contains numerous small protoscolices in brood capsules (Fig. 18.28). Apart from mild gliosis and occasional lymphocytic cuffing of blood vessels in the immediate vicinity, there is little other reaction to the cysts.
COENUROSIS
 PATHOGENESIS AND CLINICAL FEATURES OF COENUROSIS
PATHOGENESIS AND CLINICAL FEATURES OF COENUROSIS
 Coenurosis is a rare parasitic disease of humans caused by Coenurus cerebralis, the larvae of various dog tapeworms, especially Multiceps multiceps (Taenia multiceps), a parasite with a life cycle similar to that of E. granulosus. The disease has been recognized in the Caribbean and in South Africa. Most cases occur in tropical Africa, but cerebral involvement has been reported only in temperate climates.
Coenurosis is a rare parasitic disease of humans caused by Coenurus cerebralis, the larvae of various dog tapeworms, especially Multiceps multiceps (Taenia multiceps), a parasite with a life cycle similar to that of E. granulosus. The disease has been recognized in the Caribbean and in South Africa. Most cases occur in tropical Africa, but cerebral involvement has been reported only in temperate climates.


 CNS disease manifestations can include headache, transient hemiparesis, and seizures.
CNS disease manifestations can include headache, transient hemiparesis, and seizures.
 CNS SPARGANOSIS
CNS SPARGANOSIS

MACROSCOPIC AND MICROSCOPIC APPEARANCES
 PATHOGENESIS OF SPARGANOSIS
PATHOGENESIS OF SPARGANOSIS



NEMATODES
 NEMATODE INVOLVEMENT OF THE CNS
NEMATODE INVOLVEMENT OF THE CNS







ANGIOSTRONGYLIASIS
MACROSCOPIC AND MICROSCOPIC APPEARANCES

 Meningitis with lymphocytes, plasma cells, and eosinophils.
Meningitis with lymphocytes, plasma cells, and eosinophils.

 Identifiable larvae surrounded by granulomatous inflammation with giant cells.
Identifiable larvae surrounded by granulomatous inflammation with giant cells.

 PATHOGENESIS OF ANGIOSTRONGYLIASIS
PATHOGENESIS OF ANGIOSTRONGYLIASIS
 Angiostrongyliasis is caused by Angiostrongylus cantonensis, a nematode that in its adult form lives in the pulmonary arteries of rats and other rodents. It occurs in southeast Asia and the Pacific islands. The larvae migrate through the respiratory passages into the pharynx, are swallowed, and eliminated with the feces.
Angiostrongyliasis is caused by Angiostrongylus cantonensis, a nematode that in its adult form lives in the pulmonary arteries of rats and other rodents. It occurs in southeast Asia and the Pacific islands. The larvae migrate through the respiratory passages into the pharynx, are swallowed, and eliminated with the feces.


 The life cycle cannot be completed in the accidental human hosts.
The life cycle cannot be completed in the accidental human hosts.
TRICHINOSIS
 PATHOGENESIS OF TRICHINOSIS
PATHOGENESIS OF TRICHINOSIS







STRONGYLOIDIASIS
MACROSCOPIC AND MICROSCOPIC APPEARANCES
Pathologic findings may include:
 The presence of larvae in the leptomeninges (Fig. 18.29).
The presence of larvae in the leptomeninges (Fig. 18.29).
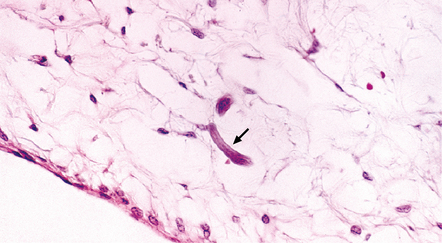
18.29 CNS strongyloidiasis. This section shows a Strongyloides stercoralis larva (arrow) in the leptomeninges.

 Microinfarcts that result from capillary obstruction by the parasites.
Microinfarcts that result from capillary obstruction by the parasites.

 PATHOGENESIS OF STRONGYLOIDIASIS
PATHOGENESIS OF STRONGYLOIDIASIS




VISCERAL LARVA MIGRANS
 PATHOGENESIS OF VISCERAL LARVA MIGRANS
PATHOGENESIS OF VISCERAL LARVA MIGRANS
 This disorder results from accidental human infection with various nematodes that usually infect non-human hosts. The disease is relatively common in the southern United States, where it is generally due to the dog ascarid, Toxocara canis. Less commonly, the cat ascarid, Toxocara cati, or the raccoon ascarid, Baylisascaris procyonis, is responsible. The adult nematode lives in the animal’s intestines and eggs are excreted in the feces.
This disorder results from accidental human infection with various nematodes that usually infect non-human hosts. The disease is relatively common in the southern United States, where it is generally due to the dog ascarid, Toxocara canis. Less commonly, the cat ascarid, Toxocara cati, or the raccoon ascarid, Baylisascaris procyonis, is responsible. The adult nematode lives in the animal’s intestines and eggs are excreted in the feces.
 Children acquire the infection by ingesting ova from contaminated soil.
Children acquire the infection by ingesting ova from contaminated soil.

TREMATODES
 SCHISTOSOMIASIS AND THE NERVOUS SYSTEM
SCHISTOSOMIASIS AND THE NERVOUS SYSTEM


MACROSCOPIC APPEARANCES
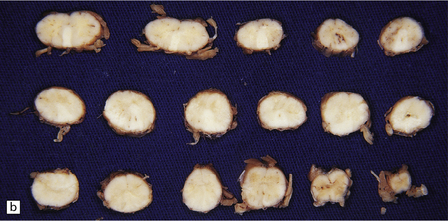
Miliary granulomas may be seen scattered throughout the brain, but are most numerous in the cerebral and cerebellar cortex and the deep gray matter. Sometimes the lesions are solitary and mimic a tumor. Spinal schistosomiasis can cause marked swelling of the affected cord segments (Fig. 18.32). Granulomas may be studded over the surface of the brain or cord, or agglomerated to form nodules, which protrude from the surface and occasionally invade the dura

MICROSCOPIC APPEARANCES
The histologic reaction to the ova varies from nothing to florid granulomatous inflammation with multinucleated giant cells, marked fibrosis, and adjacent gliosis (Fig. 18.33). Eosinophils are usually present. Some patients develop focal arteritis.
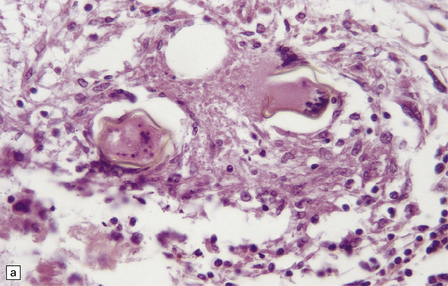
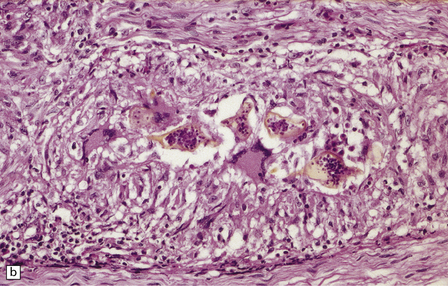
18.33 Schistosomiasis. (a) Moderate inflammatory reaction to Schistosoma ova. (b) Schistosoma ova with surrounding granulomatous inflammation in a nerve root.
 PATHOGENESIS OF SCHISTOSOMIASIS
PATHOGENESIS OF SCHISTOSOMIASIS
 Several species, including Schistosoma japonicum (endemic in China, Philippines, Japan, Thailand, and Laos), S. mansoni (in South America, Puerto Rico, the West Indies, the Middle East, and Africa) and S. haematobium (in Africa and the Middle East) are important causes of the disease.
Several species, including Schistosoma japonicum (endemic in China, Philippines, Japan, Thailand, and Laos), S. mansoni (in South America, Puerto Rico, the West Indies, the Middle East, and Africa) and S. haematobium (in Africa and the Middle East) are important causes of the disease.


 Once in water, miracidia hatch from the eggs and penetrate a snail. Fork-tailed cercariae then develop in this intermediate host, emerge into the water, and swim in search of their definitive host, entering through the skin. The cercariae invade blood vessels and eventually lodge in the mesenteric or vesical veins to mature into adult male or female trematodes (Fig. 18.31).
Once in water, miracidia hatch from the eggs and penetrate a snail. Fork-tailed cercariae then develop in this intermediate host, emerge into the water, and swim in search of their definitive host, entering through the skin. The cercariae invade blood vessels and eventually lodge in the mesenteric or vesical veins to mature into adult male or female trematodes (Fig. 18.31).
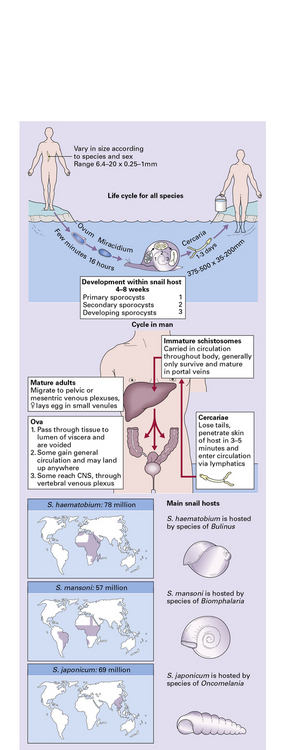
 Through pulmonary arteriovenous shunts, or portopulmonary anastomosis via the azygous vein, into the pulmonary veins, and thence the systemic arterial circulation and CNS.
Through pulmonary arteriovenous shunts, or portopulmonary anastomosis via the azygous vein, into the pulmonary veins, and thence the systemic arterial circulation and CNS.

 Possibly by anomalous migration of the adult worms, followed by egg deposition in situ in the CNS.
Possibly by anomalous migration of the adult worms, followed by egg deposition in situ in the CNS.
The distribution of the embolized ova is related to their size and shape (see box Schistosomiasis and the nervous system).
REFERENCES
Chacko, G. Parasitic diseases of the central nervous system. Semin Diagn Pathol.. 2010;27:167–185.
Feasey, N., Wansbrough-Jones, M., Mabey, D.C., et al. Neglected tropical diseases. Br Med Bull.. 2010;93:179–200.
Jansen, M., Corcoran, D., Bermingham, N., et al. The role of biopsy in the diagnosis of infections of the central nervous system. Ir Med J.. 2010;103:6–8.
Katchanov, J., Nawa, Y. Helminthic invasion of the central nervous system: many roads lead to Rome. Parasitol Int.. 2010;59:491–496.
Kristensson, K., Mhlanga, J.D., Bentivoglio, M. Parasites and the brain: neuroinvasion, immunopathogenesis and neuronal dysfunctions. Curr Top Microbiol Immunol.. 2002;265:227–257.
Lawn, S.D. Immune reconstitution disease associated with parasitic infections following initiation of antiretroviral therapy. Curr Opin Infect Dis.. 2007;20:482–488.
Lucas, S., Bell, J., Chimelli, L., Parasitic and fungal infections. 8th ed. Greenfields’ neuropathology vol. 1. London: Hodder Arnold; 2008. [1447–1512].
Lv, S., Zhang, Y., Steinmann, P., et al. Helminth infections of the central nervous system occurring in Southeast Asia and the Far East. Adv Parasitol.. 2010;72:351–408.
Deol, I., Robledo, L., Meza, A., et al. Encephalitis due to a free-living amoeba (Balamuthia mandrillaris): case report with literature review. Surg Neurol.. 2000;53:611–616.
Khan, N.A. Acanthamoeba and the blood-brain barrier: the breakthrough. J Med Microbiol.. 2008;57:1051–1057.
Marciano-Cabral, F., Cabral, G. Acanthamoeba spp as agents of disease in humans. Clin Microbiol Rev.. 2003;16:273–307.
Martinez, A.J., Visvesvara, G.S. Free-living, amphizoic and opportunistic amebas. Brain Pathol.. 1997;7:583–598.
Idro, R., Jenkins, N.E., Newton, C.R. Pathogenesis, clinical features, and neurological outcome of cerebral malaria. Lancet Neurol.. 2005;4:827–840.
Idro, R., Marsh, K., John, C.C., et al. Cerebral malaria: mechanisms of brain injury and strategies for improved neurocognitive outcome. Pediatr Res.. 2010;68:267–274.
Lou, J., Lucas, R., Grau, G.E. Pathogenesis of cerebral malaria: recent experimental data and possible applications for humans. Clin Microbiol Rev.. 2001;14:810–820.
Hill, D., Dubey, J.P. Toxoplasma gondii: transmission, diagnosis and prevention. Clin Microbiol Infect Dis.. 2002;8:634–640.
Martin, S. Congenital toxoplasmosis. Neonatal Netw.. 2001;20:23–30.
Montoya, J.G., Kovacs, J.A., Remington, J.S. Toxoplasma gondii. In: Mandell G.L., Bennet J.E., Dolin R., eds. Mandell, Douglas, and Bennett’s Principles and practice of infectious disease. 6th ed. Philadelphia: Elsevier Churchill Livingstone; 2005:3170–3198.
Almeida, E.A., Lima, J.N., Lages-Silva, E., et al. Chagas’ disease and HIV co-infection in patients without effective antiretroviral therapy: prevalence, clinical presentation and natural history. Trans R Soc Trop Med Hyg.. 2010;104:447–452.
Corti, M. AIDS and Chagas’ disease. AIDS Patient Care Stds.. 2000;14:581–588.
Enanga, B., Burchmore, R.J., Stewart, M.L., et al. Sleeping sickness and the brain. Cell Mol Life Sci.. 2002;59:845–858.
Kirchhoff, L.V. Agents of African trypanosomiasis (sleeping sickness). In: Mandell G.L., Bennet J.E., Dolin R., eds. Mandell, Douglas, and Bennett’s Principles and practice of infectious disease. 6th ed. Philadelphia: Elsevier Churchill Livingstone; 2005:3165–3170.
Kirchhoff, L.V. Trypanosoma species (American trypanosomiasis, Chagas’ disease): Biology of trypanosomes. In: Mandell G.L., Bennet J.E., Dolin R., eds. Mandell, Douglas, and Bennett’s Principles and practice of infectious disease. 6th ed. Philadelphia: Elsevier Churchill Livingstone; 2005:3157–3164.
Pittella, J.E. Central nervous system involvement in Chagas disease: a hundred-year-old history. Trans R Soc Trop Med Hyg.. 2009;103:973–978.
Rodgers, J. Trypanosomiasis and the brain. Parasitology.. 2010;137:1995–2006.
Silva, N., O’Bryan, L., Medeiros, E., et al. Trypanosoma cruzi meningoencephalitis in HIV-infected patients. J Acquir Immune Defic Syndr Hum Retrovirol.. 1999;20:342–349.
Weiss, L.M. Microsporidiosis. In: Mandell G.L., Bennet J.E., Dolin R., eds. Mandell, Douglas, and Bennett’s Principles and practice of infectious disease. 6th ed. Philadelphia: Elsevier Churchill Livingstone; 2005:3237–3254.
Fleury, A., Escobar, A., Fragoso, G., et al. Clinical heterogeneity of human neurocysticercosis results from complex interactions among parasite, host and environmental factors. Trans R Soc Trop Med Hyg.. 2010;104:243–250.
Kraft, R. Cysticercosis: an emerging parasitic disease. Am Fam Physician.. 2007;76:91–96.
Nourbakhsh, A., Vannemreddy, P., Minagar, A., et al. Hydatid disease of the central nervous system: a review of literature with an emphasis on Latin American countries. Neurol Res.. 2010;32:245–251.
Pamir, M.N., Ozduman, K., Elmaci, I. Spinal hydatid disease. Spinal Cord.. 2002;40:153–160.
Pittella, J.E. Neurocysticercosis. Brain Pathol.. 1997;7:681–693.
Rengarajan, S., Nanjegowda, N., Bhat, D., et al. Cerebral sparganosis: a diagnostic challenge. Br J Neurosurg.. 2008;22:784–786.
Sinha, S., Sharma, B.S. Neurocysticercosis: a review of current status and management. J Clin Neurosci.. 2009;16:867–876.
Taratuto, A.L., Venturiello, S.M. Echinococcosis. Brain Pathol.. 1997;7:673–679.
Alto, W. Human infections with Angiostrongylus cantonensis. Pac Health Dialog.. 2001;8:176–182.
Clausen, M.R., Meyer, C.N., Krantz, T., et al. Trichinella infection and clinical disease. QJM.. 1996;89:631–636.
Pawlowski, Z. Toxocariasis in humans: clinical expression and treatment dilemma. J Helminthol.. 2001;75:299–305.
Pien, F.D., Pien, B.C. Angiostrongylus cantonensis eosinophilic meningitis. Int J Infect Dis.. 1999;3:161–163.
Taratuto, A.L., Venturiello, S.M. Trichinosis. Brain Pathol.. 1997;7:663–672.
Tsai, H.C., Liu, Y.C., Kunin, C.M., et al. Eosinophilic meningitis caused by Angiostrongylus cantonensis: report of 17 cases. Am J Med.. 2001;111:109–114.
Carod-Artal, F.J. Neurological complications of Schistosoma infection. Trans R Soc Trop Med Hyg.. 2008;102:107–116.
Ferrari, T.C., Moreira, P.R., Cunha, A.S. Spinal-cord involvement in the hepato-splenic form of Schistosoma mansoni infection. Ann Trop Med Parasitol.. 2001;95:633–635.
Pittella, J.E. Neuroschistosomiasis. Brain Pathol.. 1997;7:649–662.