15.2 Neuraxis310
15.3 Forebrain312
15.4 Brain stem318
15.5 Cranial nerve nuclei322
15.6 Vessels of the brain323
15.7 Motor systems and tracts325
15.8 Sensory systems and tracts328
15.9 Special sensation330
15.10 Clinical miscellany332
15.11 And finally…333
15.1. Brain vesicles, general terms, ventricles, CSF
Introduction
Introductory coverage of the brain considers its components from below upwards: hindbrain, midbrain, and forebrain. However, for looking at the brain in slightly more detail, beginning at section 15.2, it seems better to work down from the cerebral hemispheres to the medulla. Embryological terms are used both descriptively and clinically, so we begin with a brief developmental survey.
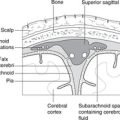
Hindbrain, midbrain, forebrain (Fig. 15.1)
Working up from the embryonic spinal cord, the three brain vesicles are the hindbrain, the midbrain and the forebrain.
 |
| Fig. 15.1 |
Hindbrain
This develops into the medulla, pons, and cerebellum. It contains the vital centres (cardiac, respiratory, etc.) and the nuclei for cranial nerves V–XII (facial sensation, and movements of the upper end of the gut tube for ingestion and phonation). The cerebellum is concerned with balance and position in space, receiving the majority of fibres from the vestibular apparatus, and with the coordination of complex patterns of skilled movement.
Midbrain
This part of the brain changes least during growth and maturation. It contains the nuclei for cranial nerves III and IV and the centres for eye reflexes (e.g. pupillary light reflex) and auditory reflexes.
Forebrain
This gives rise to the diencephalon (thalamic structures), and the right and left telencephalic derivatives – the cerebral hemispheres. The hemispheres are the only paired elements, all other components of the brain being unpaired. The cerebral hemispheres become very large and grow at first posteriorly, then downwards and forwards so that they are shaped like the horns of a ram, the inferior portions coming to lie on either side of the thalamus and midbrain, which are buried between them. The parts of the hemisphere are named according to the cranial bones to which they are related: the frontal, parietal, occipital and temporal lobes. These are not lobes in the sense of being clearly demarcated anatomical entities, but are simply areas.
• Diencephalic derivatives are concerned with motor and sensory coordination, regulation and control, and include the optic vesicles, which give rise to the optic nerves and retinas.
• Cerebral hemispheres are concerned with so-called ‘higher’ functions: intellectual, reasoning, long-term memory, conscious perception, and voluntary movement. In the cerebral hemispheres, unlike the spinal cord, grey matter is on the external surface, often called the cerebral cortex, or simply the cortex.
Brain stem
This term is used for the medulla, pons and midbrain together.
Terms derived from Greek:
• Hindbrain = rhombencephalon, rarely used.
• Midbrain = mesencephalon, widely used.
• Forebrain = prosencephalon, rarely used, of which:
– thalamic structures = diencephalon: widely used, and
– cerebral hemisphere = telencephalon: sometimes used.
15.2. Neuraxis (axis of the brain): motor and sensory areas
• In the medulla, pons and midbrain the ventral aspect of the neural tube is anterior, and the dorsal aspect is posterior, much as in the spinal cord. Above the midbrain, the axis bends forwards so that in the diencephalon, the ventral aspect is inferior and the dorsal surface superior.
• In the spinal cord, the general rule is that dorsal components tend to be concerned with afferent (sensory) functions, and ventral with efferent (motor) functions. In the medulla it is as if the two dorsal (sensory) components have been dragged laterally so that sensory components are no longer dorsal but lateral, and motor components now not so much anterior as medial. Higher in the brain stem the pattern becomes less and less obvious, and it has all but disappeared in the diencephalon.
Nucleus
A nucleus is a collection in the central nervous system of neuronal cell bodies serving similar functions. A collection of cell bodies in the peripheral nervous system is a ganglion – except that some groups of cell bodies in the brain are known as the basal ganglia, a term sanctified by long-standing use.
Ventricular system, cerebrospinal fluid
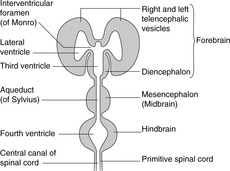
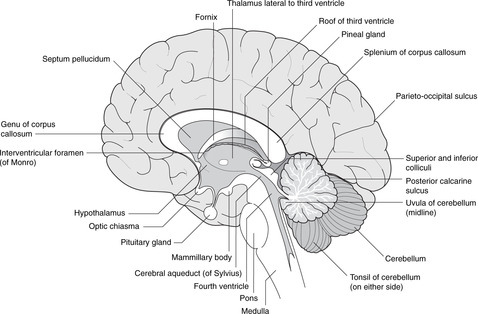
Ventricles (Figs 15.1, 15.2)
Dilatations of the central canal of the neural tube form the cerebral ventricles.
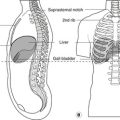
• Hindbrain. The central canal forms the fourth ventricle, continuous below with the central canal of the spinal cord. The ventral surface of the ventricle is the diamond-shaped floor (diamond-shaped brain: rhombencephalon). The thin dorsal surface is the roof and has three openings into the subarachnoid space: one median aperture, the foramen of Magendie, and two lateral apertures, the foramina of Luschka.
• Midbrain. The central canal forms the cerebral aqueduct of Sylvius.
• Diencephalon. The central cavity of the diencephalon forms the third ventricle. This is narrow from side to side (1 or 2 mm) but 10–15 mm in height. Its floor is the hypothalamus and its lateral walls the thalamus of each side, which may touch each other in the midline giving the interthalamic adhesion. You can regard the diencephalon as the structures surrounding the third ventricle or attached to its walls, thus including the optic chiasma, nerve and retina, and the posterior pituitary.
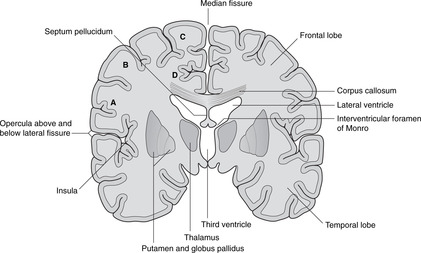
• Telencephalon. The cavities of the cerebral hemispheres are the right and left lateral ventricles. They are adjacent to the midline and are separated from each other by the septum pellucidum, a thin sheet of tissue of no neurological significance. Each lateral ventricle communicates with the third ventricle through an interventricular foramen of Monro, but the two lateral ventricles do not communicate with each other. The lateral ventricle assumes the shape of the hemisphere in which it is situated: there is an anterior (frontal) horn, a posterior (occipital) horn, and an inferior (temporal) horn.
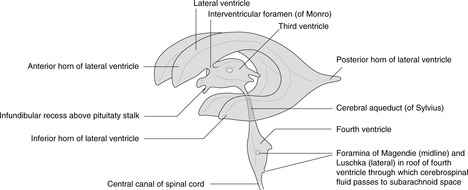
Cerebrospinal fluid
Cerebrospinal fluid is produced by the choroid plexus, a vascular plexus present in all ventricles formed where vessels of the pia mater (external) come into contact with the ependymal lining of the central canal (i.e. there is no intervening neural tissue).
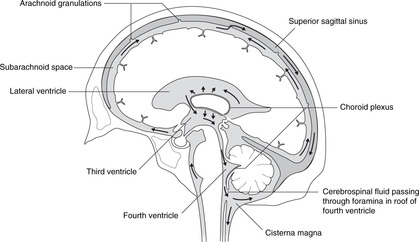
Circulation of cerebrospinal fluid (Fig. 15.3)
From the lateral ventricles, cerebrospinal fluid flows through the foramina of Monro to the third ventricle. It continues through the cerebral aqueduct to the fourth ventricle where some may enter the central canal of the spinal cord, but most passes to the subarachnoid space through foramina in the roof. It flows around the brain and spinal cord and is absorbed back into the venous system at the arachnoid granulations (see section 14.12, p. 297, Fig. 14.37).
 |
| Fig. 15.3 |
Hydrocephalus may be caused by overproduction of cerebrospinal fluid, or reduced absorption (e.g. thrombosis of the superior sagittal sinus). It may be generalised or local, for example blockage of the cerebral aqueduct would cause hydrocephalus in the lateral and third ventricles, but not in the fourth or in the subarachnoid space.
Tracts in the central nervous system, internal capsule
The brain stem contains tracts: bundles of myelinated axons (i.e. white matter) all serving similar functions. They can be divided into:
• motor or descending tracts which convey impulses away from the cerebral hemispheres; and
• sensory or ascending tracts which convey impulses towards the hemispheres.
Each of these may be further subdivided into named tracts connecting specific areas, the name of the tract generally being derived from the origin and the destination. Thus, the corticospinal tract conducts impulses from the cortex to the spinal cord – a motor tract; the spinothalamic tract conducts impulses from the spinal cord to the thalamus – a sensory tract.
Internal capsule
In the region where telencephalic and diencephalic regions meet, all the tracts in and out of the cerebral hemispheres are within a fairly small area on each side forming a sheet of white matter. This is the internal capsule, and it is covered in more detail later.
15.3. Forebrain
The cerebral cortex on each side is shaped like a ram’s horn. Its parts are named according to the bones they are related to. The principal functional areas are motor, sensory, auditory, and visual. The limbic system is the area responsible for emotions, short-term memory and instinctual behaviour. The central part of the brain, on either side of the third ventricle, is the diencephalon consisting mainly of the thalamus and hypothalamus. All tracts conveying impulses between cerebral cortex and brain stem pass in the internal capsule, which lies lateral to the thalamus, between it and the cerebral cortex. Lesions here are usually catastrophic because so many axons are packed into such a small area. The basal ganglia are a group of nuclei adjacent to the diencephalon, and in the midbrain.
You should:
• list the principal parts of the cortex
• identify the site of the motor, sensory, auditory and visual areas
• list the components of the limbic system and their functions, in as far as they are known
• list the components of the basal ganglia and their functions, in as far as they are known
• describe the anatomy of the internal capsule, and the term ‘stroke’.
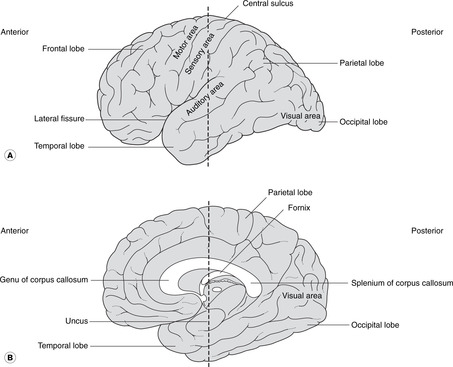
Cortex
The cerebral cortex is shown in Figures 15.4 and 15.5. The numerous small fissures are sulci (singular: sulcus) and the raised areas between them are gyri (singular: gyrus).
• The large fissure between the parietal and temporal lobes is the lateral fissure and its edges are the opercula (singular: operculum). If you separate the opercula you will see a buried area of cortex deep in the lateral fissure: this is the insula.
• Commissures. Numerous bundles of fibres (white matter) connect the two sides: these are the commissures, and the largest, deep in the sagittal fissure, is the corpus callosum. The anterior portion of the corpus callosum is the genu, the central part the body, and the posterior part the splenium. There are other commissures: the anterior commissure is part of the anterior wall of the third ventricle, and the posterior and habenular commissures are in the posterior wall of the third ventricle.
 |
| Fig. 15.4 |
Functional areas of cortex (Figs 15.4, 15.5)
Motor, sensory
On the lateral aspect of the cortex is the central sulcus (it is neither central nor easily identifiable), the approximate surface marking of its inferior limit being the pterion. The cortex anterior to the central sulcus, the precentral gyrus, is the main motor area concerned with the initiation of voluntary movement. The cortex posterior to it, the postcentral gyrus, is the main sensory area.
• For both the motor and sensory cortex the inferior (lateral) portion is concerned with the head. The rest of the body is represented sequentially passing superiorly and medially over the cortex so that the cortical areas for the lower limbs are on the medial aspect of each cerebral hemisphere with the foot area being adjacent to the corpus callosum.
• It is a fact, although one without convincing explanation, that the right cortex serves the left side of the body and vice versa.
• Broca’s area is adjacent to the head area of the dominant motor cortex (usually the left in a right-handed person). It is in the frontal lobe, immediately above the lateral fissure, and is concerned with the motor mechanisms of speech. Damage to it (for example occlusion of a branch of the middle cerebral artery) leads to motor speech aphasia (aphasia: wordless): it is not that the patient does not know the word, it is that he can not muster the motor forces to enunciate it.
Visual, auditory, olfactory
• The visual cortex is in the occipital lobe (posterior)
• The auditory cortex is on the superior surface of the temporal lobe.
• The olfactory cortex is on the under surface of the temporal lobe, principally the uncus.
Limbic system (Fig. 15.6)
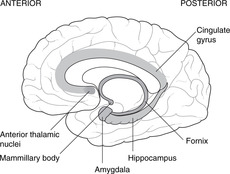
The Latin word limbus means margin or border. The limbic system develops from the border of the telencephalon adjacent to the diencephalon. Like many other telencephalic derivatives, this assumes the ram’s-horn shape. Components of the limbic system include the fornix, the hippocampus (medial to the inferior horn of the lateral ventricle), the amygdala, the uncus and the cingulate gyrus (medially, just above the corpus callosum). The limbic system has connections with olfactory and taste fibres, and with certain diencephalic structures, e.g. the mammillary bodies and anterior thalamic nuclei. Limbic functions include elements of emotion, recent memory, olfaction, taste, sexual urges, and basic instinctive drives: smells and tastes can evoke memories, emotions, mood changes and sexual responses.
Destruction or degeneration of the mammillary bodies
This occurs in alcoholics and as a result of vitamin B1 (thiamine) deficiency. Short-term memory is lost, so patients tend to cover up this defect by inventing fiction: confabulation (Korsakov’s syndrome). It has even been known to occur in operations for pituitary tumours that have gone wrong.
Congenital absence of the amygdala
Patients without an amygdala, and such people are rarely found, neither feel rage, nor recognise it in others – a blissful state, one would think. They may also exhibit hypersexuality. It is difficult to think of the amygdala as anything other than the organ of guilt and repression.
Dementias, Alzheimer’s disease
Progressive dementias may arise from disorders of the hippocampus and its connections, associated with a loss of cholinergic neurons in Meynert’s nucleus in the anterior limbic system.
 |
| Fig. 15.6 |
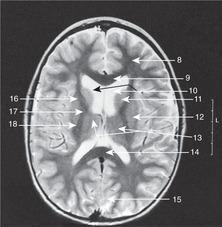
Diencephalon (Figs 15.7, 15.8, 15.10)
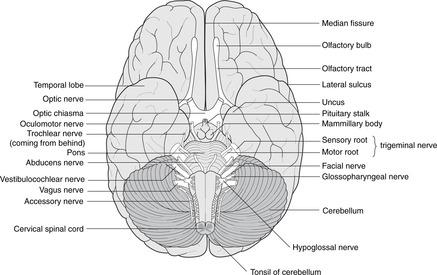
• The hypothalamus forms the floor of the third ventricle. Its most obvious features when inspecting the brain from below are the pituitary stalk and the mammillary bodies (mammilla = little mamma or breast). The hypothalamus contains the centres regulating appetite and much autonomic activity (e.g. temperature regulation) as well as controlling the activity of the pituitary gland.
– The posterior pituitary is a component part of the hypothalamus with direct neural connections from the supraoptic and paraventricular nuclei of the hypothalamus. Hormones released by the posterior pituitary (oxytocin, vasopressin) are manufactured in the cell bodies of neurons in these nuclei, and travel down axons to the posterior pituitary where they are released into the bloodstream.
– The anterior pituitary is a separate gland that migrates from the roof of the primitive pharynx. The hypothalamus controls it by releasing hormones into the hypophyseal portal system of veins, through which they are transported to the anterior pituitary to act upon its component cells.
• The roof of the third ventricle contains no neural tissue except for commissural fibres which arise anteriorly and grow backwards, over the roof of the third ventricle, to form the corpus callosum.
• The epithalamus is the posterior region of the third ventricle: it contains the pineal gland.
• The metathalamus, the most lateral part of the diencephalon, consists of the geniculate bodies (lateral: part of the visual pathway; medial: part of the auditory pathway).
Pituitary tumours
Immediately anterior to the pituitary stalk is the optic chiasma, which may be damaged by enlarging pituitary tumours causing a bitemporal hemianopia (see section 14.11, pp. 293, 294).
Pineal
The pineal gland often calcifies with advancing age. The advantage of this is that it provides a landmark for the assessment of whether or not there is any displacement of the brain: the pineal gland is a midline structure.
Internal capsule, basal ganglia (Figs 15.5, Fig. 15.10, Fig. 15.11, Fig. 15.12 and Fig. 15.13)
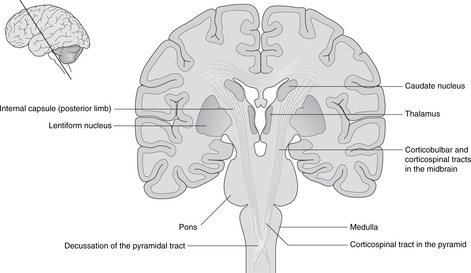
Internal capsule
The relations of the internal capsule are:
• medial to the posterior limb: the thalamus
• medial to the anterior limb: the caudate nucleus, another of the structures stretched into the shape of a ram’s horn by the growth of the brain
• lateral to both limbs: the lentiform nucleus, itself composed of a medial globus pallidus and a lateral putamen.
There are numerous fibres crossing the internal capsule linking the thalamus, caudate and lentiform nuclei, and when a fresh or fixed brain is cut, these are evident as striations. For this reason, this area was originally called the corpus striatum, and this term, although of doubtful validity, is still used.
Basal ganglia
The caudate nucleus, putamen and globus pallidus, together with the substantia nigra in the midbrain (see below) and other nuclei, constitute the basal ganglia. Their function is the control of movement, and disease of them leads to unwanted involuntary movements, for example tremors, writhing (athetosis), fidgeting (chorea) and uncontrolled explosive movements (hemiballismus). Parkinson’s disease results from disorders of the substantia nigra, and chorea and athetosis from disorders of the caudate and lentiform nuclei.
Lateral to the basal ganglia
Lateral to the basal ganglia are, in order, the external capsule, the claustrum (grey matter), the extreme capsule and the insular cortex. None of these areas is very important clinically.
Stroke
When disease (such as haemorrhage or arterial occlusion) affects the internal capsule, the result is a profound and often catastrophic loss of function because so many fibres, both motor and sensory, are concentrated into such a small area. This is, in common parlance, a stroke and is discussed in more detail later.
15.4. Brain stem
The brain stem consists of grey matter made up of cell bodies grouped as nuclei, and white matter: the myelinated axons of fibre tracts. Nuclei of the cranial nerves are prominent in the brain stem. Those of III and IV are in the midbrain, those of V–XII in the pons and medulla. Some of the tracts are short local connections which run in all directions between nuclei, and others are long motor and sensory tracts conveying information between cerebral hemispheres and spinal cord. The midbrain houses nuclei concerned with visual and auditory reflexes. The cerebellum is posterior to the pons and receives most of the information from the vestibular system. Cerebellar lesions are ipsilateral, unlike those of the rest of the brain which are usually contralateral.
Summary of brain stem nuclei
• Motor nuclei of cranial nerves III–XII.
• Parasympathetic (visceral motor) nuclei: Edinger–Westphal, superior salivatory, inferior salivatory, dorsal motor nucleus of the vagus (Fig. 14.14, p. 258).
• Sensory nuclei: trigeminal sensory nuclei for cutaneous and oral sensation, nucleus of the solitary tract for visceral sensation (e.g. taste).
• Other named nuclei for visual and auditory reflexes, as outlined in the text below.
• Nuclei of the reticular formation concerned with vital centres and arousal. These are hard to pinpoint accurately and for clinical purposes it is unnecessary to do so. One definition of the reticular formation is that it is what is left when you have removed everything else with a specific name.
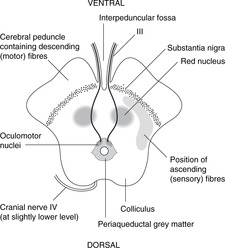
Midbrain (Fig. 15.14)
The midbrain passes through the tentorial notch of the tentorium cerebelli. It has a very distinctive shape in cross-section and is easy to draw. This is worth the effort since the midbrain is an important area from the point of view of pupillary and accommodation reflexes, descending motor tracts and other clinical conditions. Its parts are as follows.
• Tectum. This is the area dorsal to the aqueduct. There are four dorsal prominences, two superior colliculi and two inferior colliculi (also known together as the corpora quadrigemina). The superior colliculi are associated with visual reflexes and the inferior colliculi with auditory reflexes. Cranial nerve IV is attached between them.
• Periaqueductal (around the aqueduct) grey matter. This contains the nuclei of cranial nerves III and IV and the medial longitudinal fasciculus, which integrates the nuclei of III, IV and VI with vestibular information, amongst other things. In this area, fibres involved in the pupillary light reflex pass from the optic tracts to the Edinger–Westphal nucleus of III.
• The tegmentum is the area ventral to the aqueduct. One of its components is the red nucleus, to all intents and purposes an outpost of the cerebellum (see below).
• Substantia nigra (Latin: black substance). This is a dark area in the ventral tegmentum. The pigment is melanin, a reminder of the amine biochemistry commonly found in derivatives of neuroectoderm. Depleted levels of dopamine in this area lead to symptoms of Parkinson’s disease (e.g. resting tremor): functionally, this is one of the basal ganglia.
• Anterior to the substantia nigra are two large projections facing slightly laterally. These contain the main descending motor fibres called, variously, the cerebral peduncles, basis pedunculi, or cerebral crura, each being the crus cerebri. This text uses the first term.
• The fossa between the peduncles is the interpeduncular fossa in which cranial nerve III is attached.
• The long sensory tracts conveying information from the spinal cord and the lower brain stem to the thalamus are just behind the substantia nigra, extending posteriorly on to the lateral aspect of the midbrain. The long motor tracts are in the cerebral peduncles. (Dorsal: sensory; ventral: motor.)
Pons (Figs 15.7, 15.8)
The pons is that part of the brain stem to which the cerebellum and cranial nerve V are attached. It contains scattered nuclei concerned with cerebellar connections, and nuclei of cranial nerves V, VI and VII, although the course of fibres from VI and VII means that these nerves are attached lower down the brain stem at the pontomedullary junction. For reasons which are not clear, fibres of VII pass at first dorsally before looping round the nucleus of VI and turning anteriorly.
The locus coeruleus (Latin: blue place) is a pigmented area (amines again) in the dorsal pons. It is concerned with the limbic system and autonomic functions and is of interest to neuroscientists as a focus for drug action.
The long motor and sensory tracts are scattered in numerous bundles as they pass through the pontine substance.
Cerebellum (Figs 15.7, 15.8)
The cerebellum is an outgrowth of the dorsal pons, separated from it by the fourth ventricle. On each side, the white matter connecting the pons and cerebellum forms the superior, middle and inferior cerebellar peduncles. Despite all the work of neuroscientists, the cerebellum remains somewhat mysterious: it is concerned with the coordination of motor activity, the regulation of muscle tone and the maintenance of equilibrium.
Cerebellar cortex
The cerebellar cortex has superficial grey matter, inside which is white matter and, most deeply of all, the four paired cerebellar nuclei. Cerebellar equivalents of the cerebral gyri are folia (thinner and straighter than gyri). The midline portion is the vermis, the most inferior part of which is the uvula and the most superior part (on the roof of the fourth ventricle) the lingula. Laterally, the most inferior parts of the cerebellar hemispheres are the tonsils, so, as at the back of the mouth, the uvula is between the tonsils.
Vestibulocerebellum
The cerebellum, more obviously than other parts of the nervous system, displays its evolution. The earliest part to arise was that concerned with vestibular impulses, the vestibulocerebellum, and it is largely separate from the rest of the cerebellum, although this is not evident to the naked eye. It is the flocculonodular node, the floccules being on either side joined in the midline at the nodule. The deep cerebellar nucleus of the vestibulocerebellum is the fastigial nucleus, which has connections with:
• cranial nerve VIII: most fibres from the vestibular apparatus pass to the cerebellum through the inferior cerebellar peduncle
• the medial longitudinal fasciculus for the coordination of vestibular impulses and eye movements (nuclei of III, IV, VI).
Other parts of the cerebellum. Subsequently (in evolutionary terms), parts of the cerebellum acquired connections with the spinal cord (for movements of the limbs and trunk) and most recently with the cerebral cortex. The deep cerebellar nuclei for spinal connections are the globose and emboliform, and for cerebral connections the dentate nucleus.
• Impulses from the spinal cord arrive in spinocerebellar tracts.
• Impulses to and from the cerebral cortex pass from the dentate nucleus through the superior cerebellar peduncle to the red nucleus (midbrain) and on to the cerebral cortex (dentato-rubro-thalamic tract). Connections from the red nucleus pass to and from the spinal cord (thus rubrospinal, spinorubral) for further integrative activity.
Cerebellar disease
Disease of the cerebellum causes ipsilateral symptoms, not contralateral, manifested principally as a lack of coordination called ataxia. This gives rise to an intention tremor; that is to say there is no tremor present at rest, but as soon as any movement is attempted, lack of coordination becomes apparent. This contrasts with the tremor of basal ganglia disease, which is present at rest, but which disappears when purposeful movement is attempted. The resultant speech patterns are called scanning speech (it is as if the person is scanning his vocabulary for the right word, so laboured is the progress); the lack of coordination of eye movement is nystagmus. Cerebellar lesions are perfectly mimicked by drunkenness, which is nothing more or less than cerebellar poisoning by alcohol, and it is not unknown for a person with cerebellar disease to be arrested on suspicion of being intoxicated.
Pontomedullary junction (Fig. 15.15)
The pontomedullary junction is marked anteriorly by a transverse sulcus where VI is attached medially, and VII and VIII laterally in the cerebellopontine angle.
 |
| Fig. 15.14 |
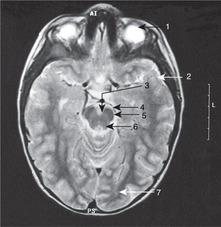
Medulla (Fig. 15.15)
Anterior aspect. Four swellings are visible, two on either side of the midline.
• The medial swelling on each side is the pyramid containing the descending motor corticospinal tracts to the opposite side of the body. The rootlets of XII are attached laterally to these swellings.
• Lateral to the pyramid is the olive containing the inferior olivary nucleus, a convoluted band of grey matter which, like the red nucleus in the midbrain, is an outpost of the cerebellum. Lateral to the olive are the rootlets of, from above down, IX, X and XI.
Posterior aspect.
Two tubercles are visible on each side, the gracile medially and the cuneate laterally. These are concerned with sensation from the spinal cord (see below).
Nuclei
The hypoglossal nucleus is close to the midline and the floor of the fourth ventricle. The motor nuclei of the vagus nerve are lateral to this (dorsal motor and nucleus ambiguus), and the ‘spinal’ part of the trigeminal sensory nucleus more lateral still.
Medulla/spinal cord junction
This is not at the foramen magnum, but a few millimetres below. It means that C1 and C2 nerves pass slightly upwards from their spinal cord origin to emerge from the vertebral column.
Some features of the fourth ventricle
• In the floor, the facial colliculus is a small swelling lateral to the central region of the ventral median sulcus. It is caused by the internal genu of the fibres of VII looping around the nucleus of VI. Lower down is the hypoglossal trigone, immediately over the nucleus of XII.
• In the roof, the foramina of Luschka (right and left) and Magendie (median) allow cerebrospinal fluid to pass from the ventricular system to the subarachnoid space.
15.5. Cranial nerve nuclei
Cranial nerve nuclei can be grouped by functional type: somatic motor, branchiomotor, visceral motor (parasympathetic), somatic sensory, visceral sensory and special sensory. These groups are disposed vertically in the brain stem, somatic motor being the most medial, with sensory nuclei more laterally sited. Not all nuclei contribute to all cranial nerves, some nerves containing only somatic motor fibres, and some only sensory fibres. Mixed nerves are those that supply the upper end of the gut tube – in embryological terms, these are the nerves of the branchial arches.
Brain stem nuclei
Lesions of the brain stem give rise to symptoms according to which cranial nerves arise near the lesion. Motor impulses in cranial nerves arise from brain stem motor nuclei, and sensory impulses in cranial nerves pass to brain stem nuclei. Table 15.1, Table 15.2 and Table 15.3 set out these nuclei in detail. As a simplification, it is enough for you to realise that:
• midbrain lesions cause disturbances of eye movements
• pontine lesions cause disturbances of mandibular movement
• medullary lesions cause disturbances of phonation and swallowing.
| Nucleus | Position | Nerve | Function |
|---|---|---|---|
| Oculomotor | Upper midbrain | Oculomotor III | Eyeball movements |
| Trochlear | Lower midbrain | Trochlear IV | Eyeball movements |
| Trigeminal | Pons | Mandibular Vc | Mastication |
| Abducens | Pons | Abducens VI | Eyeball movements |
| Facial | Pons | Facial VII | Facial expression, etc., stapedius |
| Nucleus ambiguus | Medulla | Glossopharyngeal IX | Swallowing, phonation |
| Vagus X | |||
| Accessory | See text | Accessory XI | Sternocleidomastoid, trapezius |
| Hypoglossal | Medulla | Hypoglossal XII | Tongue |
| Nucleus | Position | Nerve(s) | Function |
|---|---|---|---|
| Edinger–Westphal | Midbrain | Oculomotor III | Pupilloconstriction, accommodation |
| Superior salivatory | Pons | Facial VII, Vb | Secretomotor: nasal, palatine, lacrimal glands |
| Superior salivatory | Pons | Facial VII,Vc | Secretomotor: submandibular, sublingual glands |
| Inferior salivatory | Upper medulla | Glossopharyngeal IX | Secretomotor: parotid gland |
| Dorsal motor of vagus | Medulla | Vagus X | Heart, foregut, midgut |
| Source of stimulus | Ganglion | Nucleus and/or connections | |
|---|---|---|---|
| Cutaneous and oral sensation | |||
| V | Oral and nasal cavities, teeth, TMJ, skin of anterior scalp, face, most of external ear | Trigeminal | Trigeminal: Spinal (pain, temperature) Principal (tactile) |
| X | Some skin of external ear | Sensory ganglia of X | Mesencephalic (proprioception, see text) |
| Visceral sensation | |||
| VII | Taste buds, anterior, tongue, etc. | Geniculate VII | |
| IX | Mucosa of oropharynx Taste buds, posterior tongue Carotid body |
Sensory ganglia of IX | |
| X | Mucosa of hypopharynx, larynx Taste buds, epiglottic region Aortic body Thoracoabdominal viscera |
Sensory ganglia of X | Nucleus of solitary tract |
| Special sensation | |||
| I | Olfactory epithelium | Olfactory bipolar cells | Olfactory bulb, olfactory cortex, etc. |
| II | Rods and cones | Retinal bipolar cells | Lateral geniculate body, visual cortex |
| VIII | Organ of Corti | Cochlear (spiral) | Cochlear nuclei, auditory cortex |
| VIII | Saccule, utricle, semicircular ducts | Vestibular | Vestibular nuclei, cerebellum |
15.6. Vessels of the brain
You should:
• describe the territories of the vertebrobasilar system and the anterior, middle and posterior cerebral arteries
• describe the likely effects of blockage of these vessels
• describe the vessels that supply the internal capsule and the likely effect of their blockage
• revise the anatomy of the venous sinuses.
Arteries
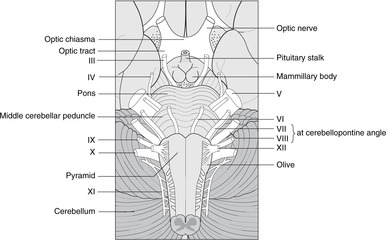
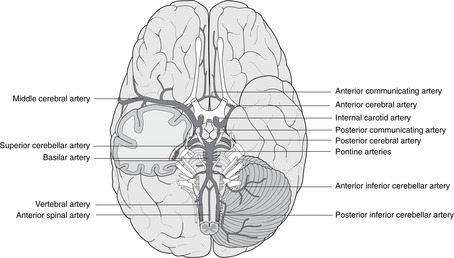
Vertebrobasilar system, posterior cerebral artery (Fig. 15.16)
The vertebrobasilar system supplies the brain stem and visual cortex.
• After the right and left vertebral arteries have pierced the dura in the region of the foramen magnum they give branches which include:
– anterior and posterior spinal arteries, which pass down to supply the spinal cord
– posterior inferior cerebellar artery (PICA), which supplies the lateral medulla and the lower cerebellum.
• The vertebrals unite to form the basilar artery, which ascends on the ventral aspect of the pons giving:
– pontine branches
– the labyrinthine artery, which enters the internal acoustic meatus and supplies the inner ear
– anterior inferior and superior cerebellar arteries.
• The basilar artery terminates by contributing to the arterial circle of Willis, giving:
Lateral medullary syndrome (Wallenberg’s or PICA syndrome)
This is caused by thrombosis of the posterior inferior cerebellar artery. Since this supplies the upper lateral medulla, some of the symptoms are explicable on the basis of the involvement of the spinal part of the trigeminal sensory nucleus and the nucleus ambiguus: disturbances of facial temperature sensation and paralysis of speech and swallowing. There may also be cerebellar involvement and balance disorders (because of possible involvement of cochlear and vestibular nuclei). The spinothalamic tract conveying pain and temperature sensation from the contralateral trunk and limbs may also be involved.
Medial medullary syndrome
The medial medulla is supplied by the anterior spinal artery. This involves the hypoglossal nucleus, corticospinal tract and medial lemniscus.
 |
| Fig. 15.15 |
Internal carotid artery (Fig. 15.16)
The intracranial course of the internal carotid artery was described in Chapter 14 (section 14.12, p. 296). It terminates by giving the anterior and middle cerebral arteries as part of the arterial circle of Willis.
Arterial circle of Willis (Fig. 15.16)
The circle is formed by: the posterior cerebral arteries, the posterior communicating arteries, the internal carotid artery (briefly), the anterior cerebral artery and the (single) anterior communicating artery. There is much variation and asymmetry. The anastomotic possibilities of the circle are not normally brought into action unless there is some requirement for increased blood flow as a result of disease or blockage elsewhere.
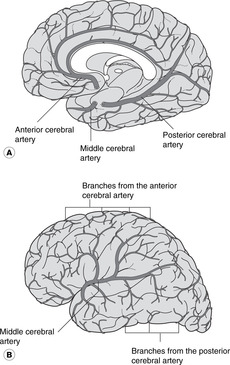
Anterior cerebral artery (Fig. 15.17)
• This passes forwards and medially under the genu of the corpus callosum, then upwards and posteriorly on the superior surface of the corpus callosum between the cerebral hemisphere and the falx cerebri. It runs very close to its opposite number (and would be better called the medial cerebral artery).
• It supplies the lower limb areas of both motor and sensory cortex.
Middle cerebral artery (Fig. 15.17)
From its origin, the middle cerebral artery passes laterally to the insula and on to the lateral aspect of the cerebral hemisphere through the lateral fissure.
• It supplies most of the parietal lobe, all the temporal lobe and part of the occipital lobe. This includes most of the sensory, motor and auditory cortical areas.
• The middle cerebral, and the termination of the internal carotid, give off a group of arteries, sometimes known as central arteries, supplying the internal capsule and basal ganglia:
– lenticulo(thalamo)striate arteries, which supply the basal ganglia and internal capsule, blockage or haemorrhage of which will cause a catastrophic stroke
– choroidal branches, which supply the choroid plexus and diencephalon.
• As the middle cerebral artery emerges on to the lateral aspect of the hemisphere, it provides a rich arterial supply for Broca’s motor speech area and the head and neck areas of the sensory and motor cortex.
Veins
These were described in Chapter 14 (section 14.12, pp. 296, 297).
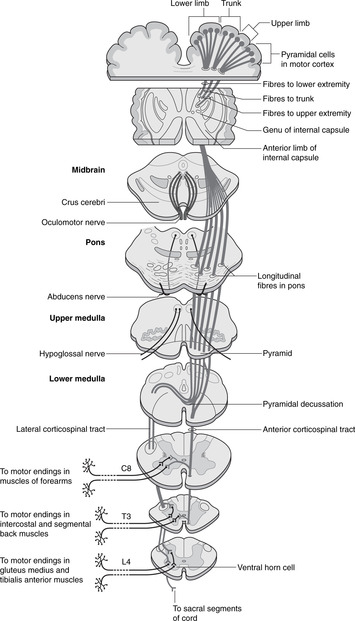
15.7. Motor systems and tracts (Fig. 15.18)
Voluntary motor systems pass from the motor cortex to cell bodies of lower motor neurons. Corticobulbar fibres pass to cranial nerve motor nuclei, most decussating towards the end of their journey. Corticospinal fibres pass to spinal cord ventral horn cells. Most decussate in the medulla at the decussation of the pyramids, but some do so just before terminating on the ventral horn cells. Interruption of these pathways gives pyramidal signs and symptoms. Lower motor neuron cell bodies also receive impulses form other brain centres (basal ganglia, reticular formation, etc.) and lesions of these are referred to as extrapyramidal syndromes. Both pyramidal and extrapyramidal lesions are examples of upper motor neuron lesions: such lesions interrupt motor impulses before they reach the cell body of the lower motor neuron.
You should:
• describe the layout of the corticobulbar and corticospinal pathways
• describe the position of the decussation and the synapse in these pathways
• explain the effects of interrupting these pathways
• describe in simple terms the extrapyramidal system
• explain what constitutes an upper motor neuron lesion.
Motor systems
Impulses leave the cortex in corticofugal pathways. Between the motor cortex and skeletal (striated) muscles, the pathways are arranged in two neuronal groups: upper motor neurons and lower motor neurons.
• Lower motor neurons have cell bodies in the central nervous system: for spinal nerves these are in the ventral horn of grey matter of the spinal cord, and for cranial nerves, in the motor nucleus of the nerve concerned (III, IV, Vc, VI, VII, nucleus ambiguus, XII). The axons of lower motor neurons pass out in peripheral nerves to the muscle fibres without further synapse.
Corticospinal, corticobulbar, corticonuclear, pyramidal, extrapyramidal pathways: what do these terms mean?
• The corticobulbar or corticonuclear pathway conveys impulses from cortex to cranial nerve motor nuclei. The terms are synonymous even though not all the cranial nerve motor nuclei are in the bulb, an archaic name for the medulla.
• The corticospinal pathway conveys impulses from cortex to ventral horn cells of the spinal cord. Since the corticospinal pathways pass deep to the pyramids of the medulla, they are known as the pyramidal pathways – a widely used clinical term.
• Extrapyramidal pathway. Upper motor neurons include fibres from other brain centres (e.g. basal ganglia, cerebellum, reticular formation) that connect with the lower motor neurons, thus influencing their activity. These do not pass through the pyramids – they are not components of the corticospinal pathways – and so are grouped together as extrapyramidal pathways. It is important to remember these when considering what may go wrong (see later).
Upper motor neurons for cranial nerves: corticonuclear, corticobulbar
For cranial nerves, cell bodies of upper motor neurons are in the head area of the motor cortex. Axons descend through the genu of the internal capsule, decussating just before synapsing with cell bodies of lower motor neurons that make up the motor nucleus of that cranial nerve. Functionally, these pathways are similar to the corticospinal pathway, but of course the term corticospinal should not be used, since the impulses never descend as far as the spinal cord!
Upper motor neurons for spinal nerves: corticospinal
For spinal nerves, cell bodies of upper motor neurons are in the appropriate area of the motor cortex. Axons descend through the posterior limb of the internal capsule, the cerebral peduncles of the midbrain, the pons and the pyramids of the medulla. Immediately below the pyramids, just above the beginning of the spinal cord, most of them (about 85%) decussate to form the lateral corticospinal tract of the spinal cord. They descend in this tract to the appropriate level in the spinal cord and synapse on the ventral horn cells – the cell bodies of the lower motor neurons. The remainder descend without decussating as the anterior corticospinal tract, but even these decussate at the last minute, just before synapsing on ventral horn cells.
Pyramidal decussation
The decussation is just below the pyramids. There is no synapse here: the axon merely changes sides.
Lower motor neuron lesion – flaccidity, hyporeflexia, wasting, ipsilateral
If all lower motor neurons passing to a muscle are severed, the muscle will be completely paralysed. It will be flaccid (atonic, hypotonic), it will not respond to reflexes (arreflexic, hyporeflexic) since no impulses reach it, and it will fairly quickly atrophy as a result of denervation. The injury and the paralysis are on the same side: they are ipsilateral with respect to each other.
Upper motor neuron lesion – spasticity, hyperreflexia, contralateral (usually)
If upper motor neurons to a muscle are severed, the ability to control and initiate movement in the muscle may be lost. However, lower motor neurons are intact, and since some of the fibres to lower motor neurons from elsewhere are inhibitory, other centres which influence lower motor neurons, e.g. basal ganglia, may cause an increase in muscle tone (hypertonic, spastic). Also, reflexes are disinhibited (hyperreflexic, exaggerated). The muscle will not become atrophied except through disuse. Most upper motor neuron lesions are in the internal capsule (strokes) and in this case, since upper motor neurons have not yet decussated, the functional loss will be on the side opposite to the site of the lesion: they are contralateral with respect to each other.
These characteristics of upper motor neuron lesions (UMNL) and lower motor neuron lesions (LMNL) are clinically very important.
Bulbar palsy: ipsilateral lower motor neuron lesion
This is caused by a lesion of the medulla that involves the nucleus ambiguus and the hypoglossal nucleus. It may arise from blockage of the posterior inferior cerebellar artery (Fig. 15.16). The cell bodies of lower motor neurons are affected, with resultant ipsilateral motor disorders of chewing (to a variable extent), swallowing and speaking. Because the nuclei concerned are in the bulb (medulla), this is a bulbar palsy.
Pseudobulbar palsy: contralateral upper motor neuron lesion
This is caused by interruption of the upper motor neuron pathways, usually in the internal capsule, cerebral peduncles or pons. The contralateral muscles of the tongue and pharynx would be affected. Since this presents as a disorder of the muscles of speech and swallowing, it is at first sight like a bulbar palsy, but being an upper motor neuron lesion, it is caused by a lesion on the opposite side. It is known as a pseudobulbar palsy.
Bilateral upper motor neuron control of cranial nerves III, IV, VI and part of VII
The pattern in the head and neck, as in the rest of the body, is that the motor cortex innervates contralateral motor nuclei. However, muscles which move the eyes, and the eyelids and forehead in association with eye movements, receive bilateral cortical innervation. The nuclei concerned are those of III, IV and VI, and that portion of the facial (VII) motor nucleus that innervates orbicularis oculi and frontalis. This must have evolved in association with, and for the protection of, the sense of sight, by which means we seek sustenance and mates, and avoid danger.
There may be limited bilateral control of the other cranial nerve motor nuclei as is evidenced by partial recovery of function in patients after a stroke.
Upper motor neuron lesions of VII
Because of the bilateral cortical control of the upper facial muscles, in upper motor neuron lesions of VII, the forehead and orbicularis oculi muscles are largely spared. For the lower facial muscles this is not so: the normal pattern prevails with only contralateral control. So: the upper part of the face is spared in upper motor neuron lesions of VII.
Another interesting feature of upper motor neuron lesions of VII is that although patients may be unable, at will, to move facial muscles around the mouth, they may smile at something they consider amusing. This is a reminder of the fact that the lower motor neurons are intact and are able to respond to impulses from other than the corticobulbar pathway (in this case presumably from the jocularity centre – if it exists).
Facial palsy (lower motor neuron lesion)
A lower motor neuron lesion of VII, a facial palsy, may result from a lesion of the facial motor nucleus, or the nerve itself, and the muscle fibre(s) supplied by the affected neuron would be paralysed. There is no question of partial preservation of function in this case: if the lower motor neuron is damaged, nothing can reach the muscle. Facial wrinkles disappear if the muscles are paralysed, so it is usually easy to spot. Bell’s palsy is a facial palsy of unknown aetiology. See Chapter 14 (section 14.6, p. 267).
VI and VII
The relationship between the nucleus of VI and the axons of VII means that a brain stem lesion may cause facial weakness in association with a paralysis of the ipsilateral lateral rectus muscle of the eye.
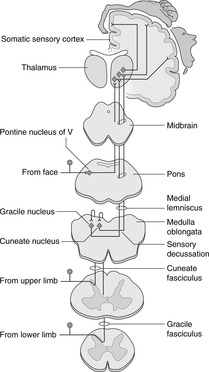
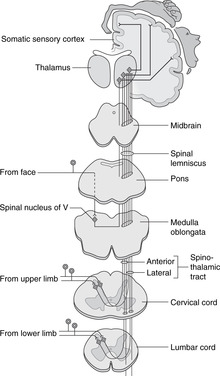
15.8. Sensory systems and tracts (Figs 15.19, 15.20)
Sensory impulses are delivered to the central nervous system in neurons with cell bodies in peripheral ganglia. Pathways decussate after the first synapse in the CNS and pass to the contralateral thalamus before being relayed to various parts of the brain. Within the spinal cord, some modalities (e.g. fine touch) are carried by tracts of neurons before decussating, so are ipsilateral with respect to the source of the impulse (dorsal column pathways). Other modalities (e.g. pain, temperature) are carried by fibre tracts of neurons after synapsing, so are contralateral with respect to the source of the impulse (spinothalamic pathways). Sensory information from cranial nerves is handled by brain stem nuclei of V before going on to the thalamus. Visceral sensation, such as taste, or sensation from internal organs, passes to the nucleus of the solitary tract in the brain stem.
 |
| Fig. 15.18 |
 |
| Fig. 15.19 |
 |
| Fig. 15.20 |
Basic plan of sensory systems
There are no synapses outside the brain and spinal cord: the first synapse is in the central nervous system between primary and secondary sensory neurons.
From spinal nerves
There are two systems, spinothalamic (or anterolateral) and the dorsal columns. For confirmation of this, consider what happens when you stub your toe. You are aware of something having touched your toe about half a second or so before feeling the pain. The tactile information (dorsal columns) speeds up the spinal cord much more quickly than the pain information (spinothalamic), speed in this case being a result of larger diameter, more heavily myelinated fibres.
Dorsal columns: touch, vibration sense, limb position sense (Fig. 15.19)
• From the dorsal rootlets of spinal nerves, fibres pass into the adjacent ipsilateral dorsal column of white matter and ascend to the top of the spinal cord. Fibres from the sacral region lie most medially, the higher the subsequent level of entry, the more laterally placed the fibre. Fibres in the dorsal white column form two bundles, the medial being thin (gracile) and the lateral larger and wedge-shaped (cuneate).
• At the top of the spinal cord these fibres synapse on cell bodies of secondary sensory neurons in the gracile and cuneate nuclei, and axons cross the midline to ascend to the contralateral thalamus through the medulla, pons and midbrain in a tract called the medial lemniscus.
• The spinal cord tract in this system is made up of primary sensory neurons and is ipsilateral with respect to the source of the impulse.
Spinothalamic (anterolateral): pain, temperature (Fig. 15.20)
• From the dorsal rootlets of spinal nerves, fibres enter the adjacent dorsal horn of grey matter and synapse in this region within a segment or two.
• Axons of secondary sensory neurons cross the midline and ascend in the spinothalamic tracts in the lateral white column to the ventral nucleus of the thalamus.
• The spinal cord tract in this system is made up of secondary sensory neurons and is contralateral with respect to the source of the impulse.
From cranial nerves
Most somatic sensory fibres are carried in V and a small number enter the brain stem in X, but within the central nervous system all somatosensory fibres pass to the trigeminal sensory nuclei.
Trigeminal sensory nuclei
The trigeminal sensory nucleus has three parts, each for different modalities.
• Principal or chief nucleus: tactile sensation. This is in the pons and receives the central processes of primary sensory neurons transmitting tactile sensation. From the nucleus, axons decussate and ascend in the trigeminal lemniscus to the contralateral thalamus. A small proportion of fibres may also pass to the ipsilateral thalamus.
• The spinal nucleus: pain and temperature sensation. This is so called because it extends down through the medulla into the cervical spinal cord and is continuous with the dorsal horn of spinal grey matter (which also receives pain and temperature fibres). From the nucleus, axons decussate and ascend in the trigeminal lemniscus to the contralateral thalamus.
• The mesencephalic nucleus: proprioceptive sensation, etc. This is in the upper pons and lower midbrain and receives proprioceptive information from masticatory muscles, and deep pressure sensation from the teeth and gums (see section 14.7, pp. 273, 274). The mesencephalic nucleus is unique since it houses primary sensory nerve cell bodies which, for all other sensory fibres, would be in a peripheral ganglion. Details of its connections are not clear, but some pass to the motor nucleus of V, the salivatory nuclei, and the nucleus ambiguus, for chewing and swallowing.
Visceral sensation (e.g. taste, internal organs)
Much less is known about visceral sensation pathways. All that you need to know is that it is all handled by the nucleus of the solitary tract in the lateral medulla, no matter how it gets there (see Fig. 14.15, p. 259). Axons from the nucleus of the solitary tract pass to the salivatory nuclei (for salivation reflexes) and also rostrally by multisynaptic pathways, to the thalamus, and probably the insula and uncus for connections with olfactory centres. As pointed out earlier (p. 257), none of this matters clinically.
15.9. Special sensation
Olfactory impulses pass to components of the limbic system and the temporal lobe. Visual impulses pass to the occipital cortex for vision and to the midbrain for visual reflexes. Auditory pathways pass to the temporal lobe. Vestibular impulses pass mainly to the cerebellum. There are numerous other connections allowing for integration between systems, both within the brain and from brain to spine cord. A good example of such a pathway is the medial longitudinal fasciculus that coordinates cranial nerve nuclei of III, IV and VI with vestibular and cerebellar function.
Olfactory pathways in the brain
The olfactory epithelium and nasal cavity were considered in Chapter 14 (section 14.9, p. 279).
The connections of the olfactory bulb are not fully established. Olfactory impulses pass posteriorly from the olfactory tract to the limbic system, particularly the uncus and amygdala of the temporal lobe, thus providing connections with memory circuitry and much else.
Olfaction and taste
The pleasures of eating and drinking lie as much with smell as with taste: it is a common experience that an upper respiratory tract infection which interferes with the sense of smell impairs the enjoyment of food. Olfaction and taste are clearly closely linked and it is thought that from the nucleus of the solitary tract, to which taste fibres pass, axons project to the uncus to connect with olfactory centres.
Temporal lobe tumours
Diseases such as epilepsy in the areas to which the olfactory impulses project (e.g. the temporal lobe) may cause olfactory hallucinations. The smells are usually unpleasant and are often accompanied by pseudo-purposeful movements associated with tasting, such as licking the lips. Temporal lobe tumours may also present as mood or memory disturbances – more evidence of limbic functions.
Visual pathways in the brain
The visual pathways as far back as the lateral geniculate bodies were considered in Chapter 14 (section 14.11, p. 288).
Lateral geniculate body, optic radiation, visual cortex
In the lateral geniculate body, axons of retinal ganglion cells synapse with cell bodies of neurons forming the optic radiation. This passes backwards, skirting the posterior limb of the internal capsule and the lentiform nucleus (thus they are described as retrolenticular), and around the inferior horn of the lateral ventricle to end in the visual cortex in the occipital lobe.
• Lesions of this part of the visual pathway depend upon precisely which part of the lateral geniculate body or optic radiation is affected since there is a precise retinotopic arrangement in the optic radiation and visual cortex.
• Revise the connections of the visual pathway with the midbrain (section 14.11, p. 294) and Fig. 14.39 (p. 295) for the pathway of the pupillary light and other reflexes.
Blood supply of visual cortex (Fig. 15.17)
Although the posterior cerebral artery supplies most of the visual cortex, both middle and posterior arteries generally supply the cortex to which the macula of the retina projects. This is one of the explanations given for the phenomenon of macular sparing in which vision at the macula may be preserved even though the surrounding areas of the visual cortex are no longer functional. As with the bilateral cortical innervation of muscles involved in eye movements, there may be an evolutionary basis for this.
Auditory pathways in the brain
The auditory pathways as far back as the cochlear nuclei were considered in Chapter 14 (section 14.10, p. 287).
Cochlear nuclei, medial geniculate body, auditory cortex
From the cochlear nuclei, neurons pass bilaterally to the inferior colliculi (midbrain, for auditory reflexes) and the medial geniculate bodies (diencephalon). Some neurons pass to other centres (e.g. medial longitudinal fasciculus, reticular formation, spinal cord) for integration with other systems. Axons from medial geniculate bodies project through the internal capsule to the auditory cortex in the upper part of the temporal lobe on the inferior operculum, in the territory of middle cerebral artery.
Note. Visual system: lateral geniculate bodies, superior colliculi; auditory system: medial geniculate bodies, inferior colliculi.
Vestibular pathways in the brain
The vestibular pathways and the cerebellum have been considered above (section 15.3, p. 312) and in Chapter 14 (section 14.10, p. 287).
15.10. Clinical miscellany
Your attention has already been drawn to clinical conditions illustrating individual points of the anatomy of the head and neck and cranial nerves. The anatomy of the brain is so complex, however, that a single lesion can have profound effects on a combination of different neurological systems and some of these are considered here.
Stroke, cerebrovascular accident (CVA)
• A lesion in the genu of the internal capsule would produce sensory and motor loss in the contralateral side of the head. This may not be complete since there is bilateral cortical innervation of some cranial nerve nuclei. The motor symptoms could produce a pseudobulbar palsy (see p. 328).
• A lesion in the posterior limb of the internal capsule would produce motor and sensory signs in the contralateral side of the body.
• Monoplegia is weakness or paralysis of one limb, possibly arising as a result of a lesion of a specific limb area of one motor cortex.
• Hemiplegia is weakness or paralysis of one side of the body, possibly arising as a result of an internal capsule lesion, or a lesion elsewhere on the corticospinal pathway.
• Quadriplegia is weakness or paralysis of all four limbs, probably arising as a result of injury to the cervical spine which completely sections the spinal cord below the origin of the phrenic nerve (so diaphragmatic breathing is preserved). In these cases, the spinal cord below the lesion is completely cut off from the brain and each spinal cord segment functions only as a component of a segmental reflex. The sphincters innervated by S2–4 would revert to automatic emptying as required as a result of reflex sensory stimulation.
Bilateral upper motor neuron lesions of the lower limbs
Your detective powers may be tried by this until you realise that because of the anatomy of the motor cortex, and the manner in which the body is represented on it, the two motor areas for the lower limbs are actually very close to one another on the medial sides of the two cerebral hemispheres: they are separated only by the falx cerebri. In theory, these symptoms could be produced by occlusion of both anterior cerebral arteries which supply these areas, but, on the ground that one explanation is better than two, the symptoms could be produced by an enlarging tumour of the falx itself which presses on the medial surfaces of both hemispheres. This is a sagittal meningioma.
Pyramidal, extrapyramidal
Paralysis with hyperreflexia and hypertonia probably means a lesion of the corticospinal (pyramidal) tract. These are often called pyramidal symptoms.
Uncontrolled involuntary movements probably indicate a lesion of the basal ganglia, or tracts descending from them which do not pass deep to the pyramids. These are often called extrapyramidal symptoms.
Medial longitudinal fasciculus
Multiple sclerosis
Multiple (disseminated) sclerosis is a disease of myelin produced by oligodendrocytes. It can strike anywhere in the central nervous system causing symptoms as a result of defective neurotransmission.
Spinal cord lesions and sensory symptoms
From receptors in one part of the body, some sensory impulses ascend in the contralateral side of the spinal cord, and some in the ipsilateral. To any muscle, most impulses come from the contralateral cortex but descend in the ipsilateral side of the cord. The interpretation of symptoms of spinal cord lesions is therefore not straightforward. For example, if the right half of the spinal cord is damaged at spinal cord segment T6:
• all ipsilateral sensory fibres entering at that level will be affected
• all ipsilateral motor fibres leaving at that level will be affected
• contralateral pain and temperature fibres from the body at all levels below the injury will be interrupted
• ipsilateral dorsal column sensation from the body at all levels below the injury will be interrupted.
It is somewhat less complex with motor symptoms because most motor fibres in the spinal cord pass to the same side of the body.
These detective games are often made more difficult by the fact that there may be other features complicating the clinical picture.
15.11. And finally…
When you reflect on this brief consideration of the brain, you may be able to perceive a caudal (bottom) to rostral (top) progression in functional terms. Unconscious mechanical functions, such as would be possessed by a primitive vertebrate, seem to be housed in the hindbrain. More sophisticated functions like visual and auditory reflexes, so essential in detecting approaching predators or prey, are prominent in the midbrain. Bodily homeostasis is a function of the diencephalon. The limbic system in the caudal telencephalon deals with urges and emotions and, most rostrally of all, what passes for intellectual function is based in the cortex. You might like to consider whether or not some psychiatric disease is caused by the intellectual activity of the cerebral cortex suppressing the basic urges of the limbic system. If you feel the need to throw this book at someone, and do so, you might be preserving your mental health but you must be prepared to take the consequences.
Self-assessment: questions, Chapter 15
This section also tests some head and neck material from Chapter 14. There are more questions here than is, perhaps, warranted by the length of Chapter 15: For some reason, I find it easier to think of questions on head and neck and CNS than on other topics, and every physician I have spoken to thinks that students never know enough about cranial nerves.
True/false questions
Each lettered option is either true or false.
1. The following cranial nerves arise as stated:
a. XII: medulla, lateral to olive.
b. X: medulla, medial to olive.
c. VI, VII: cerebellopontine angle.
d. V: pons.
e. III: interpeduncular fossa of midbrain.
2. The following are midline structures:
a. the foramen of Magendie.
b. the foramen of Monro.
c. the pineal.
d. the thalamus.
e. the pyramid.
3. The following are derivatives of the diencephalon:
a. the third ventricle.
b. the substantia nigra.
c. the retina.
d. the thalamus.
e. the neurohypophysis.
4. A voluntary motor impulse from the motor cortex to the right biceps brachii muscle:
a. passes through the posterior limb of the left internal capsule.
b. passes through the left substantia nigra of the midbrain.
c. decussates above the pyramids of the medulla.
d. descends in the spinal cord in the right lateral corticospinal tract.
e. synapses on ventral horn cells in spinal cord segments C5 and C6.
5. Fine touch and vibration sense impulses from the skin over the left patella:
a. synapse in the left dorsal root ganglion of L3.
b. ascend in the left dorsal column of white matter of the spinal cord.
c. decussate in the lower medulla.
d. ascend through the brain stem as the right medial lemniscus.
e. pass in the midbrain ventral to the substantia nigra.
6. A left pupil that does not respond to the shining of a light into the right eye might result from:
a. an opaque right cornea and/or lens.
b. a severed right optic nerve.
c. damage to the right lateral geniculate body.
d. damage to the right Edinger–Westphal nucleus.
e. damage to the left oculomotor nerve.
7. The lateral medullary (Wallenberg’s or PICA) syndrome:
a. may be caused by blockage of the posterior inferior cerebellar artery.
b. may cause disturbances of facial temperature sensation.
c. would impair swallowing and speaking.
d. may involve the nucleus ambiguus.
e. would involve the pyramids.
8. All the following structures are found in the midbrain:
a. red nucleus.
b. substantia nigra.
c. superior colliculus.
d. facial colliculus.
e. aqueduct of Sylvius.
9. The head and neck area of the motor cortex:
a. lies in the postcentral gyrus.
b. extends on to the medial surface of the hemisphere.
c. is supplied by the anterior cerebral artery.
d. is part of the occipital lobe.
e. is related to the pterion.
11. Lower motor neuron lesions are characterised by:
a. flaccidity.
b. hyporeflexia.
c. wasting.
d. sensory loss.
e. contralateral symptoms with respect to site of lesion.
12. The spinothalamic tracts serve:
a. two-point discrimination.
b. pain.
c. conscious proprioception.
d. heat.
e. vision.
13. A loss of short-term memory is likely to be caused by a lesion of the:
a. caudate nucleus.
b. fornix.
c. mammillary bodies.
d. putamen.
e. thalamus.
14. The following structures develop from the hindbrain:
a. cerebellum.
b. lamina terminalis.
c. corpora quadrigemina.
d. medulla oblongata.
e. hypothalamus.
15. The third ventricle:
a. is part of the midbrain.
b. lies between the two thalamuses.
c. communicates with the fourth ventricle by the cerebral aqueduct.
d. is related to the pineal gland.
e. does not produce cerebrospinal fluid.
16. The following arteries form the circle of Willis:
a. middle meningeal
b. anterior cerebral.
c. anterior communicating.
d. posterior communicating.
e. vertebral.
17. The posterior limb of the internal capsule:
a. is medial to the thalamus.
b. is lateral to the caudate nucleus.
c. contains corticobulbar fibres.
d. contains corticospinal fibres.
e. contains only motor fibres.
18. The pituitary:
a. is between the cavernous sinuses.
b. is directly superior to the maxillary sinus.
c. is part of the thalamus.
d. pituitary tumour may produce bitemporal hemianopia.
e. the anterior pituitary is developed from the roof of the buccal cavity.
19. Concerning the retina:
a. it has two layers.
b. the optic disc lies lateral to the macula lutea.
c. the optic disc has only rods as photoreceptors.
d. rods and cones lie adjacent to the vitreous humour.
e. its blood supply comes from the central artery of the retina.
20. A tumour in the cerebellopontine angle may cause:
a. conductive deafness.
b. ipsilateral facial paralysis.
c. sensorineural deafness.
d. sensory loss over the contralateral forehead.
e. visual field defects.
Single best answer questions
For each numbered question, only ONE of the options lettered a–d is correct.
21. A voluntary motor impulse from the cerebral cortex to the right gastrocnemius:
a. decussates in the pons.
b. originates in the left motor cortex adjacent to the falx cerebri.
d. passes through the tectum of the midbrain.
22. A voluntary motor impulse from the cerebral cortex to the right masseter:
a. decussates below the pyramids of the medulla.
b. passes through the anterior limb of the left internal capsule.
c. synapses in the left substantia nigra of the midbrain.
d. synapses on lower motor neuron cell bodies in the trigeminal motor nucleus.
23. Fine touch and vibration sense impulses from the skin over the left patella:
a. ascend in the right dorsal column of white matter of the spinal cord.
b. decussate in the lower medulla.
c. pass in the midbrain ventral to the substantia nigra.
d. synapse in the left dorsal root ganglion of L3.
24. Complete destruction of the fibres anterior to the substantia nigra in the rightcerebral peduncle of the midbrain would result in:
a. left sided hyperreflexia.
b. loss of conscious proprioception on the right side of the body.
c. loss of pain sensation on the left side of the body.
d. right sided lack of coordination.
25. The head and neck area of the motor cortex:
a. extends on to the medial surface of the hemisphere.
b. is part of the occipital lobe.
c. is related to the pterion externally.
d. is supplied by the anterior cerebral artery.
26. In the internal capsule:
a. blood supply is from branches of the middle cerebral artery.
b. the anterior limb contains the main motor and sensory tracts.
c. the caudate nucleus is lateral to the anterior limb.
d. the optic radiation occupies the anterior limb.
27. The spinothalamic tracts serve:
a. conscious proprioception.
b. hearing.
c. pain.
d. two-point discrimination.
28. Which of the following is a midline structure:
a. aqueduct of Sylvius.
b. foramen of Luschka.
c. lentiform nucleus
d. olive.
29. A lesion of which of the following may cause an intention tremor:
a. the caudate nucleus.
b. the cerebellum.
c. the pyramid.
d. the substantia nigra.
30. Concerning the midbrain:
a. it is closely related to the free border of the falx cerebri.
b. the cerebral peduncles lie posteriorly.
c. the fourth cranial nerve takes origin from the upper part.
d. the superior colliculi are connected with auditory reflexes.
31. The lateral ventricle:
a. does not produce cerebrospinal fluid.
b. extends into the temporal lobe.
c. is the cavity of the midbrain.
d. opens into the third ventricle by the cerebral aqueduct.
32. Concerning the fourth ventricle:
a. it communicates with the subarachnoid space by a single aperture.
b. it does not produce cerebrospinal fluid.
c. the abducens nerve is in its roof.
d. the posterior aspect of the pons and medulla form the anterior wall.
33. The cranial subdural space:
a. contain choroid plexus.
b. contains cerebrospinal fluid.
c. contains meningeal vessels.
d. is traversed by cerebral veins before they drain into the dural venous sinuses.
35. Within the skull the following are associated:
a. aneurysm of the anterior cerebral artery…paralysis of medial rectus muscle.
b. extradural haemorrhage…shift of the calcified pineal gland on radiograph.
c. extradural haemorrhage…venous bleed.
d. occlusion of the middle cerebral artery…a lesion of the thalamus.
Matching item questions
Questions 36–45: blood vessels
Match the numbered item to the lettered response. Each lettered response may be used once, more than once, or not at all.
a. anterior cerebral artery or branches.
b. middle cerebral artery or branches.
c. posterior cerebral artery or branches.
d. vertebrobasilar system.
e. middle meningeal artery or branches.
36. auditory cortex.
37. cranial dura.
38. diploë.
39. frontal cortex.
40. internal capsule.
41. medulla.
42. most of visual cortex.
43. motor cortex for upper limb.
44. motor nucleus for phonation and swallowing.
45. sensory and motor cortex for lower limb.
Questions 46–54: brain stem nuclei and tracts
Match the numbered item to the lettered response. Each lettered response may be used once, more than once, or not at all.
a. facial motor nucleus.
b. medial longitudinal fasciculus.
c. nucleus ambiguus.
d. solitary tract and nucleus (NTS).
e. spinal nucleus of V.
46. chemoreceptor sensation.
47. earache.
48. facial pain.
49. facial paralysis.
50. hyperacusis.
51. muscles of swallowing and phonation.
52. nystagmus.
53. taste sensation.
54. toothache.
Questions 55–65: cranial nerves
Match the numbered item to the lettered response. Each lettered response may be used once, more than once, or not at all.
a. cranial nerve III
b. cranial nerve IV
c. cranial nerve Va
d. cranial nerve Vb
e. cranial nerve Vc
f. cranial nerve VI
g. cranial nerve VII
h. cranial nerve X
i. cranial nerve XII
55. abduction of eyeball.
56. blinking.
57. facial sensation, between eye and mouth.
58. forehead sensation.
59. nerve arising from dorsal midbrain.
60. nerve at risk in middle ear disease.
61. nerve at risk in parotid tumours.
62. nerve involved in pupillary light reflex.
63. smiling.
64. swallowing, phonation.
65. tongue movements.
Questions 66–72: blood vessels
Match the numbered item to the lettered response. Each lettered response may be used once, more than once, or not at all.
b. branch of external carotid artery.
c. branch of internal carotid artery.
d. branch of anterior cerebral artery.
e. branch of middle cerebral artery.
f. branch of posterior cerebral artery.
g. vertebrobasilar system.
66. facial artery.
67. maxillary artery.
68. passes close to corpus callosum.
69. supplies auditory cortex.
70. supplies brain stem.
71. supplies head and neck area of motor cortex.
72. supplies lower limb area of motor cortex.
Questions 73–80: brain lesions
Match the numbered item to the lettered response. Each lettered response may be used once, more than once, or not at all.
a. lesion of caudate/lentiform complex (corpus striatum)
b. lesion of cerebellum, flocculonodular node
c. lesion of cerebellar hemispheres
d. lesion of mammillary bodies
e. lesion of occipital lobe
f. lesion of substantia nigra
g. lesion of temporal lobe
73. hearing voices.
74. intention tremor.
75. involuntary writhing/jerky/sinuous movements.
76. nystagmus.
77. Parkinson’s disease.
78. scanning speech.
79. short term memory loss.
80. visual problems.
Questions 81–84: visual field defects
Match the numbered item to the lettered response. Each lettered response may be used once, more than once, or not at all.
a. binasal hemianopia
b. bitemporal hemianopia
c. right eye, complete blindness
d. right homonymous hemianopia
81. bilateral internal carotid artery aneurysms.
82. pituitary tumour.
83. section of right optic nerve.
84. right optic tract lesion.
Questions requiring short answers
85. Trace the course of a motor impulse from the motor cortex to the left masseter muscle. Give details of cell bodies, synapse(s), decussations (if any), and give the blood supply of that part of the pathway within the forebrain.
86. Where is CSF produced, how does it circulate and where is it resorbed? What would happen if its passage through the midbrain were blocked?
87. What and where is the substantia nigra? What is known of its function and its connections?
88. Trace the course of a pain impulse from the left upper incisors. Give details of cell bodies, synapse(s), decussation (if any), and give the blood supply of those parts of the brain to which these impulses are relayed.
89. Describe the connections of the vestibular system. Explain why vestibular disease may give rise to nystagmus.
90. A patient imagines he can detect odours that other people can not. When out walking he is almost knocked down by a car because he did not hear it approaching. After some weeks he begins to notice mood disturbance and personality changes. Can you put this story together with one cause?
91. Following a cerebrovascular accident (stroke), a patient is unable to swallow normally. Trace the course of a motor nerve impulse from the motor cortex to the left pharyngeal musculature. Include details of central pathways, synapses (if any), decussation (if any), and peripheral course. Explain where the lesion might be and add any other notes you wish.
92. Compare and contrast the trigeminal and pterygopalatine ganglia.
Self-assessment on Chapter 15: answers
True/false answers
1.
b. False. It arises lateral to the olive, with IX and XI.
c. False. VI does not, VII does.
d. True.
e. True.
2.
a. True.
b. False. Each lateral ventricle communicates with the third ventricle through a foramen of Monro, so there are right and left.
c. True.
d. False. There is a right thalamus and a left thalamus.
e. False. There are right and left pyramids.
3.
a. True.
b. False. The substantia nigra is in the midbrain (mesencephalon).
c. True.
d. True.
e. True.
4.
a. True.
b. False. It passes anterior to the substantia nigra, not through it.
c. False. It decussates below the pyramids.
d. True.
e. True.
5.
a. False. Nothing ever synapses in sensory ganglia.
b. True.
c. True.
d. True.
e. False. Sensory tracts are dorsal to the substantia nigra.
6.
a. True.
b. True.
c. False. Geniculate bodies have nothing to do with the pupillary light reflex.
d. False. The left Edinger–Westphal nucleus would be relevant, but not the right.
e. True.
7.
a. True.
b. True. The spinal trigeminal nucleus may be affected.
c. True.
d. True.
e. False. The pyramids are medial.
8.
a. True.
b. True.
c. True.
d. False. It is in the medulla.
e. True.
9.
a. False. It is in the precentral gyrus.
b. False. The lower limb portion is medial.
c. False. It is supplied by the middle cerebral artery.
d. False. It is part of the frontal lobe.
e. True.
10.
a. True. It is medial to both.
b. False. The optic radiation is not really part of the internal capsule at all.
c. False. These are in the posterior limb.
d. False. The caudate nucleus is medial to the anterior limb.
e. True.
11.
a. True.
b. True.
c. True.
d. False. This is a motor lesion.
e. False. Symptoms are ipsilateral with respect to the site of the lesion.
12.
a. False. This is served by the dorsal columns.
b. True.
c. False. This is served by the dorsal columns.
d. True.
e. False. This is nothing to do with the spinothalamic tracts.
13 A loss of short term memory is likely to be caused by a lesion of the:
a. False. This is one of the basal ganglia.
c. True. This is part of the limbic system, imvolved in short term memory function.
d. False. This is one of the basal ganglia.
e. False.
14.
a. True.
b. False. This is a forebrain (diencephalic) structure.
c. False. This is another name for the colliculi of the midbrain.
d. True.
e. False. This is a forebrain (diencephalic) structure.
15.
a. False. It is part of the diencephalon.
b. True.
c. True.
d. True.
e. False. Choroid plexus is present in all ventricles.
16.
a. False.
b. True.
c. True.
d. True.
e. False.
17. The posterior limb of the internal capsule:
a. False. It is lateral to the thalamus.
b. True.
c. False. These fibres are in the genu.
d. True.
e. False. Both motor and sensory.
18.
a. True.
b. False. It is superior to the sphenoid sinus.
c. False. It is attached to the hypothalamus.
d. True.
e. True. Tumours of this embryonic tissue are called craniopharyngiomas.
19.
a. True.
b. False. It’s medial.
c. False.
d. False.
e. True.
20.
a. False. See below.
b. True.
c. True.
d. False. This is supplied by Va, which arises in the pons.
e. False. II is nowhere near.
Single best answers
21. b. It originates in the lower limb area on the medial surface of the motor cortex.
22. d. It passes through the genu of the internal capsule, the cerebral peduncle of the midbrain anterior to the substantia nigra.
23. b.
24. a. These are motor fibres, so an upper motor neuron lesion would result.
25. c.
26. a.
27. c. And temperature.
28. a.
29. b.
30. a. This may be relevant when the brain stem is displaced by a space-occupying lesion: midbrain damage can result.
31. b.
32. d.
33. d.
34. d.
35. b.
Matching item answers
36. b.
37. e.
38. e. The middle meningeal system is important in supplying cranial bones.
39. a.
40. b.
41. d.
42. c.
43. b.
44. d. This is the nucleus ambiguus in the medulla.
45. a.
46. d.
48. e. All cranial nerve pain fibres synapse here.
49. a.
50. a. Stapedius.
51. c.
52. b.
53. d.
54. e. All cranial nerve pain fibres synapse here.
55. f.
56. g.
57. d.
58. c.
59. b.
60. g.
61. g.
62. a.
63. g.
64. h.
65. i.
66. b.
67. b.
68. d.
69. e.
70. g.
71. e.
72. d.
73. g. Auditory hallucinations are likely to mean a lesion of the auditory cortex in the temporal lobe.
74. c.
75. a.
76. b. Also caused by lesion of medial longitudinal fasciculus and/or vestibular system.
77. f.
78. c.
79. d.
80. e.
81. a. Pressing on lateral aspect of chiasma.
82. b. Pressing on crossing fibres of chiasma.
83. c.
84. d.
Short answers
85. The impulse begins in the right motor cortex deep to pterion (supplied by the middle cerebral artery), and the corticobulbar axon passes through the genu of the internal capsule (supplied by the middle cerebral artery), and the right cerebral peduncle of the midbrain ventral to the substantia nigra. Fibres decussate just before reaching the left trigeminal motor nucleus in the pons, where they synapse on the lower motor neuron cell bodies in the trigeminal nucleus. From there, axons pass in the left trigeminal nerve and branches of the mandibular nerve to the masseter muscle.
86. CSF is produced in the choroid plexus in all ventricles and reabsorbed into the venous system at the arachnoid granulations. Its circulation around the brain and spinal cord is shown in Figure 15.3 (p. 312). If the cerebral aqueduct were blocked, CSF pressure in the lateral and third ventricles would rise.
87. The substantia nigra is an area of pigmented (melanin) grey matter in midbrain, separating the cerebral peduncles from the rest of the midbrain. Depleted dopamine here leads to Parkinson’s disease. It is one of the basal ganglia and has connections with the other members of this group (caudate nucleus, putamen, globus pallidus: nigrostriatal) and with the spinal cord (nigrospinal). Unwanted involuntary movements are a feature of basal ganglia (extrapyramidal) disease.
88. Pain impulses pass in palatine branches of the left maxillary nerve (Vb) and then gain the maxillary nerve itself. They then run in Vb through the foramen rotundum to the trigeminal ganglion (cell bodies, no synapse). The trigeminal nerve enters the pons (supplied by the vertebrobasilar system), where pain fibres synapse in the spinal nucleus of V (supplied by the vertebrobasilar system) and axons decussate and ascend to the right thalamus (supplied by the right middle cerebral artery), and on to higher centres including the right sensory cortex (supplied by the right middle cerebral artery).
Nystagmus results from overstimulation of the medial longitudinal fasciculus as a result of vestibular disease.
90. A temporal lobe tumour is the probable cause. These may present with:
• olfactory hallucinations: uncus
• failure to localise the direction of sound: auditory cortex
• mood changes: components of the limbic system (amygdala, hippocampus).
91. False. From the right motor cortex, the impulse passes through the genu of the right internal capsule, the right cerebral peduncle, and the right pons, crossing to the left nucleus ambiguus in the medulla. Here it synapses and impulses pass in the left vagus and through pharyngeal branches to pharyngeal muscles. The lesion could be in the left nucleus ambiguus (bulbar palsy), or the corticobulbar tract above it (e.g. right internal capsule).
92. Similarities: both are swellings on nerves, both contain cell bodies. Differences: the trigeminal ganglion is sensory, so has no synapses; the pterygopalatine is parasympathetic, so contains synapses. The trigeminal ganglion is susceptible to anaesthetics; the pterygopalatine is susceptible to neurotransmitter blocking agents. This is a bit of a ‘so-what?’ question, somewhat nitpicking. But if you understand these things, you have grasped a great deal.