[level-membership-for-rheumatology-category]Chapter 10
Axial Skeleton
Muscle and Joint Interactions
INNERVATION OF THE MUSCLES AND JOINTS WITHIN THE TRUNK AND CRANIOCERVICAL REGIONS
TRUNK AND CRANIOCERVICAL REGIONS
Special Considerations for the Study of Muscle Actions within the Axial Skeleton
Muscles of the Trunk: Anatomy and Their Individual Actions
Set 1: Muscles of the Posterior Trunk (“Back” Muscles)
Set 2: Muscles of the Anterior-Lateral Trunk (“Abdominal” Muscles)
Set 3: Additional Muscles (Iliopsoas and Quadratus Lumborum)
Muscles of the Trunk: Functional Interactions among Muscles
Muscles of the Craniocervical Region: Anatomy and Their Individual Actions
Muscles of the Craniocervical Region: Functional Interactions among Muscles That Cross the Craniocervical Region
SELECTED BIOMECHANICAL ISSUES OF LIFTING: A FOCUS ON REDUCING BACK INJURY
Muscular Mechanics of Extension of the Low Back during Lifting
Osteologic and arthrologic components of the axial skeleton are presented in Chapter 9. Chapter 10 focuses on the many muscle and joint interactions occurring within the axial skeleton. The muscles control posture and stabilize the axial skeleton, protect the spinal cord and internal organs, produce torques required for movement of the body as a whole, and, lastly, furnish fine mobility to the head and neck for optimal placement of the eyes, ears, and nose. Muscles associated with ventilation and mastication (chewing) are presented in Chapter 11.
INNERVATION OF THE MUSCLES AND JOINTS WITHIN THE TRUNK AND CRANIOCERVICAL REGIONS
An understanding of the organization of the innervation of the craniocervical and trunk muscles begins with an appreciation of the formation of a typical spinal nerve root (Figure 10-1). Each spinal nerve root is formed by the union of ventral and dorsal nerve roots: the ventral nerve roots contain primarily “outgoing” (efferent) axons that supply motor commands to muscles and other effector organs associated with the autonomic system. The dorsal nerve roots contain primarily “incoming” (afferent) dendrites, with the cell body of the neuron located in an adjacent dorsal root ganglion. Sensory neurons transmit information to the spinal cord from the muscles, joints, skin, and other organs associated with the autonomic nervous system.
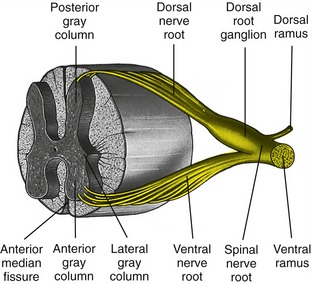
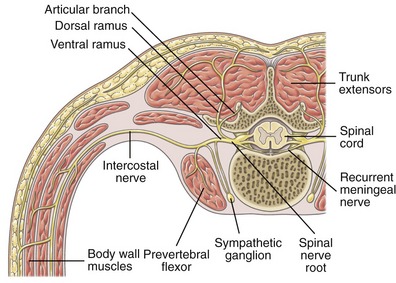
Once a spinal nerve root exits its intervertebral foramen, it immediately divides into a ventral and dorsal ramus (the Latin word ramus means “path”) (see Figure 10-1). Depending on location, the ventral ramus forms nerves that innervate, in general, the muscles, joints, and skin of the anterior-lateral trunk and neck, and the extremities. The dorsal ramus, in contrast, forms nerves that innervate, in general, the muscles, joints, and skin of the posterior trunk and neck. This anatomic organization is depicted generically by the illustration in Figure 10-2.
Ventral Ramus Innervation
INDIVIDUAL NAMED NERVES
Many of the ventral rami within the trunk and craniocervical regions do not join a plexus; rather, they remain as individual named nerves. Each of these nerves typically innervates only a part or a segment of a muscle or connective tissue. This is why, for instance, many muscles that extend across a large part of the axial skeleton possess multiple levels of segmental innervation. The two most recognized sets of individual segmental nerves derived from the ventral rami are the intercostal (thoracic) and the recurrent meningeal nerves (see Figure 10-2).
Intercostal Nerves (T1 to T12): Each of the 12 ventral rami of the thoracic spinal nerve roots forms an intercostal nerve, innervating an intercostal dermatome and the set of intercostal muscles that share the same intercostal space. (Refer to dermatome chart in Appendix III, Part B, Figure III-2.) The T1 ventral ramus forms the first intercostal nerve and part of the lower trunk of the brachial plexus. The ventral rami of T7-T12 also innervate the muscles of the anterior-lateral trunk (i.e., the “abdominal” muscles). The T12 ventral ramus forms the last intercostal (subcostal) nerve and part of the L1 ventral ramus of the lumbar plexus.
Recurrent Meningeal Nerves: A single recurrent meningeal (sinuvertebral) nerve branches off the extreme proximal aspect of each ventral ramus. After its bifurcation, the recurrent meningeal nerve courses back into the intervertebral foramen (hence the name “recurrent” [see Figure 10-2]). As a set, these often very small nerves provide sensory and sympathetic nerve supply to the meninges that surround the spinal cord, and to connective tissues associated with the interbody joints.23 Most notably, the recurrent meningeal nerve supplies sensation to the posterior longitudinal ligament and adjacent areas of the superficial part of the annulus fibrosus. Sensory nerves innervating the anterior longitudinal ligament reach the spinal cord via small branches from nearby ventral rami and adjacent sympathetic connections.23
Dorsal Ramus Innervation
A dorsal ramus branches from every spinal nerve root, innervating structures in the posterior trunk usually in a highly segmental fashion. With the exception of the C1 and C2 dorsal rami, which are discussed separately, all dorsal rami are smaller than their ventral rami counterparts (see Figure 10-2). In general, dorsal rami course a relatively short distance posteriorly (dorsally) before innervating selected adjacent muscles and connective tissues on the back of the trunk (Box 10-1).
TRUNK AND CRANIOCERVICAL REGIONS
The muscles of the axial skeleton are organized into two broad and partially overlapping areas: the trunk and the craniocervical region (Table 10-1). The muscles within each area are further organized into sets, based more specifically on their location.
TABLE 10-1.
Anatomic Organization of the Muscles of the Axial Skeleton*
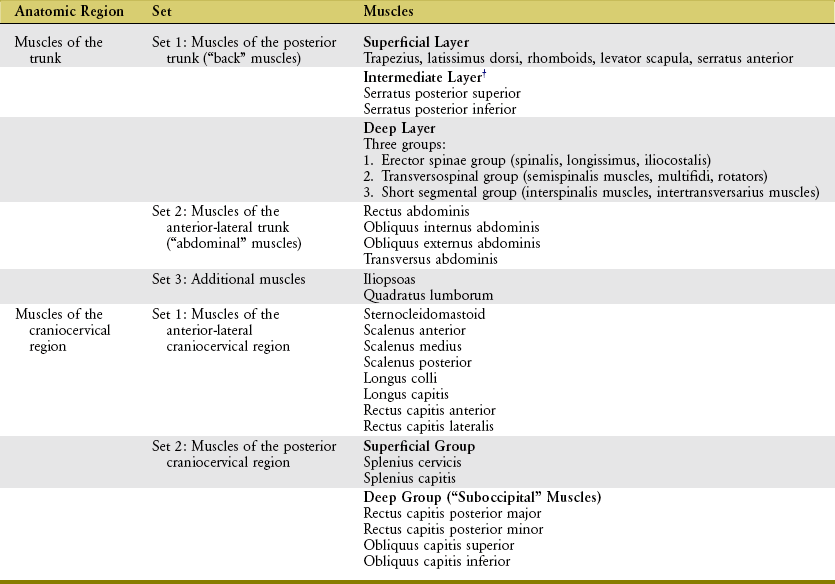
*A muscle is classified as belonging to the “trunk” or “craniocervical region” based on the location of most of its attachments.
The muscles within each area of the body are presented in two sections, the first covering anatomy and individual muscle actions, and the second covering examples of the functional interactions among related muscles. Throughout this chapter, the reader is encouraged to consult Chapter 9 for a review of the pertinent osteology related to the attachments of muscles. Appendix III, Part C should be consulted for a summary of more detailed muscular anatomy and innervation of the muscles of the axial skeleton.
Production of Internal Torque
By convention, the “strength” of a muscle action within the axial skeleton is expressed as an internal torque, defined for the sagittal, frontal, and horizontal planes. Within each plane, the maximal internal torque potential is equal to the product of (1) the muscle force generated parallel to a given plane, and (2) the length of the internal moment arm available to the muscle (Figure 10-3).
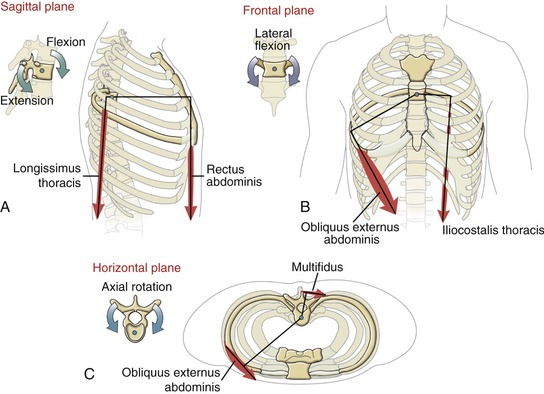
The spatial orientation of a muscle’s line of force determines its effectiveness for producing a torque for a particular action. Consider, for example, the obliquus externus abdominis muscle producing a force across the lateral thorax, with a line of force oriented about 30 degrees from the vertical (Figure 10-4). The muscle’s resultant force vector can be trigonometrically partitioned into unequal vertical and horizontal force components. The vertical force component—about 86% of the muscle’s maximal force—is available for producing lateral flexion or flexion torques. The horizontal force component—about 50% of the muscle’s maximal force—is available for producing an axial rotation torque. (This estimation is based on the cosine and sine of 30 degrees, respectively.) For any muscle of the axial skeleton to contribute all its force potential toward axial rotation, its overall line of force must be directed solely in the horizontal plane. For a muscle to contribute all its force potential toward either lateral flexion or flexion-extension, its overall line of force must be directed vertically. (Realize, though, that a vertically oriented muscle cannot produce any axial rotation because it lacks the moment arm required to produce a torque in the horizontal plane. As described in Chapter 1, a muscle force is incapable of producing a torque within a given plane if it either parallels or pierces the associated axis of rotation.)
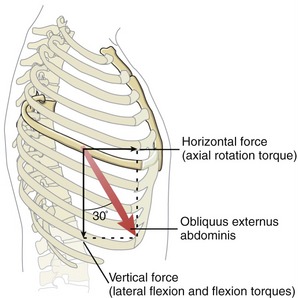
The lines of force of muscles that control movement of the axial skeleton have a spatial orientation that varies over a wide spatial spectrum, from nearly vertical to nearly horizontal. This fact is important with regard to a muscle or muscle group’s torque potential for a given action. For instance, because more of the total muscle mass of the trunk is biased vertically than horizontally, maximal efforts usually produce greater frontal and sagittal plane torques than horizontal plane torques.145
Muscles of the Trunk: Anatomy and Their Individual Actions
The following section describes the relationships between the anatomy and the actions of the muscles of the trunk. Musculature is divided into three sets: (1) muscles of the posterior trunk, (2) muscles of the anterior-lateral trunk, and (3) additional muscles (see Table 10-1).
SET 1: MUSCLES OF THE POSTERIOR TRUNK (“BACK” MUSCLES)
The muscles of the posterior trunk are organized into three layers: superficial, intermediate, and deep (see Table 10-1).
Muscles in the Superficial and Intermediate Layers of the Back: The muscles in the superficial layer of the back are presented in the study of the shoulder (see Chapter 5). They include the trapezius, latissimus dorsi, rhomboids, levator scapula, and serratus anterior. The trapezius and latissimus dorsi are most superficial, followed by the deeper rhomboids and levator scapula. The serratus anterior muscle is located more laterally on the thorax.
The muscles included in the intermediate layer of the back are the serratus posterior superior and the serratus posterior inferior. They are located just deep to the rhomboids and latissimus dorsi. The serratus posterior superior and inferior are thin muscles that contribute little to the movement or stability of the trunk. Their function is more likely related to the mechanics of ventilation and therefore is described in Chapter 11.
SPECIAL FOCUS 10-1  Muscles of the Superficial Layer of the Back: an Example of Muscles “Sharing” Actions between the Axial and Appendicular Skeletons
Muscles of the Superficial Layer of the Back: an Example of Muscles “Sharing” Actions between the Axial and Appendicular Skeletons
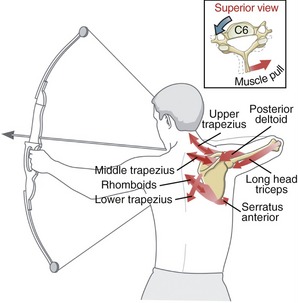
Chapter 5 describes the actions of the muscles of the superficial layer of the back, based on their ability to rotate the appendicular skeleton (i.e., humerus, scapula, or clavicle) toward a fixed axial skeleton (i.e., head, sternum, vertebral column, or ribs). The same muscles, however, are equally capable of performing the “reverse” action (i.e., rotating segments of the axial skeleton toward the fixed appendicular skeleton). This muscular action is demonstrated by highlighting the functions of the trapezius and rhomboids during use of a bow and arrow. As indicated in Figure 10-5, several muscles produce a force needed to stabilize the position of the scapula and abducted arm. Forces produced in the upper trapezius, middle trapezius, and rhomboids simultaneously rotate the cervical and upper thoracic spine to the left, indicated by the bidirectional arrows.23,82 This “contralateral” axial rotation effect is shown for C6 in the inset within Figure 10-5. As the muscle pulls the spinous process of C6 to the right, the anterior side of the vertebra is rotated to the left. The trapezius and rhomboids also stabilize the scapula against the pull of the posterior deltoid, long head of the triceps, and serratus anterior. The shared actions of these muscles demonstrate the inherent efficiency of the musculoskeletal system. In this example, a few muscles accomplish multiple actions across both the axial and the appendicular skeletons.
Muscles in the Deep Layer of the Back: Muscles in the deep layer of the back are the (1) erector spinae group, (2) transversospinal group, and (3) short segmental group (Table 10-2). The anatomic organization of the erector spinae and transversospinal groups is illustrated in Figure 10-6.
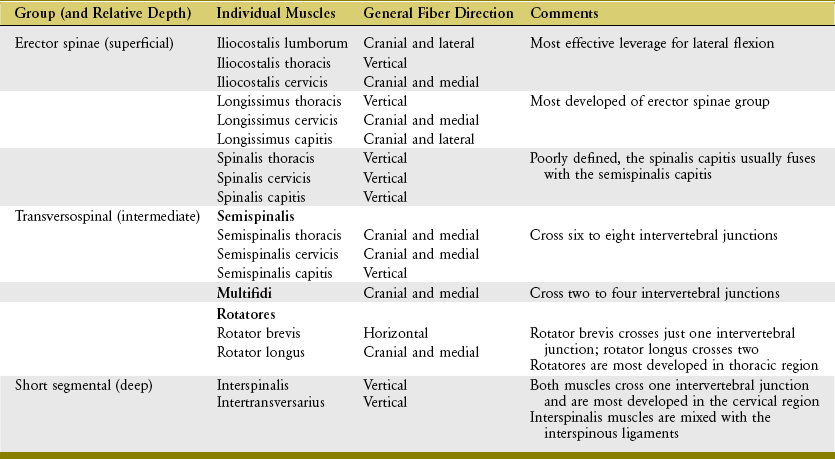
Although a few exceptions prevail, muscles in the deep layer of the back are innervated segmentally through the dorsal rami of spinal nerves.132 A particularly long muscle within the erector spinae group, for instance, is innervated by multiple dorsal rami throughout the spinal cord. A shorter muscle such as one multifidus, however, is innervated by a single dorsal ramus.100
Erector Spinae Group: The erector spinae are an extensive and rather poorly defined group of muscles that run on both sides of the vertebral column, roughly within one hand’s width from the spinous processes (Figure 10-7). Most are located deep to the posterior layer of thoracolumbar fascia (see Chapter 9) and the muscles in the intermediate and superficial layers of the back. The erector spinae consist of the spinalis, longissimus, and iliocostalis muscles. Each muscle is further subdivided topographically into three regions, producing a total of nine named muscles (see Table 10-2). Individual muscles overlap and vary greatly in size and length.23
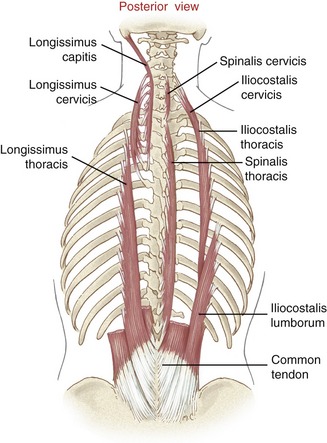
The bulk of the erector spinae muscles have a common attachment on a broad and thick common tendon, located in the region of the sacrum (see Figure 10-7). This common tendon anchors the erector spinae to many locations (Box 10-2). From this common tendon arise three poorly organized vertical columns of muscle: the spinalis, longissimus, and iliocostalis.132 The general muscle attachments are described in the following sections; more specific attachments can be found in Appendix III, Part C.
Spinalis Muscles: Spinalis muscles include the spinalis thoracis, spinalis cervicis, and spinalis capitis. In general, this small and often indistinct (or missing) column of muscle arises from the upper part of the common tendon. The muscle ascends by attaching to adjacent spinous processes of most thoracic vertebrae or, in the cervical region, the ligamentum nuchae. The spinalis capitis, if present, often blends with the semispinalis capitis.132
Longissimus Muscles: The longissimus muscles include the longissimus thoracis, longissimus cervicis, and longissimus capitis. As a set, these muscles form the largest and most developed column of the erector spinae group. The fibers of the longissimus thoracic muscles fan cranially from the common tendon, attaching primarily to the posterior end of most ribs. In the neck, the longissimus cervicis angles slightly medially before attaching to the posterior tubercle of the transverse processes of the cervical vertebrae (see Figure 10-7). The longissimus capitis, in contrast, courses slightly laterally and attaches to the posterior margin of the mastoid process of the temporal bone. The slightly more oblique angulation of the superior portion of the longissimus capitis and cervicis suggests that these muscles assist with ipsilateral axial rotation of the craniocervical region.
Iliocostalis Muscles: The iliocostalis muscles include the iliocostalis lumborum, iliocostalis thoracis, and iliocostalis cervicis. This group occupies the most lateral column of the erector spinae. The iliocostalis lumborum arises from the common tendon and courses upward and slightly outward to attach lateral to the angle of the lower ribs. The iliocostalis thoracis continues vertically to attach just lateral to the angle of the middle and upper ribs. From this point, the iliocostalis cervicis continues cranially and slightly medially to attach to posterior tubercles of the transverse processes of the midcervical vertebrae, along with the longissimus cervicis.
Summary: The erector spinae muscles cross a considerable distance throughout the axial skeleton. This anatomic feature suggests a design more suited for control of gross movements across a large part of axial skeleton (such as extending the trunk while rising from a low chair) rather than finer movements at selected intervertebral junctions.15 Bilateral contraction of the erector spinae as a group extends the trunk, neck, or head (Figure 10-8).47 The muscles’ relatively large cross-sectional areas enable them to generate large extension torque across the axial spine, such as for lifting or carrying heavy objects.37
By attaching to the sacrum and to the pelvis, the erector spinae can anteriorly tilt the pelvis, thereby accentuating the lumbar lordosis. (Pelvic tilt describes a sagittal plane rotation of the pelvis around the hip joints. The direction of the tilt is indicated by the rotation direction of the iliac crests.) As depicted in Figure 10-8, A, the anterior pelvic tilt is accentuated by the increased tension in stretched hip flexor muscles, such as the iliacus.
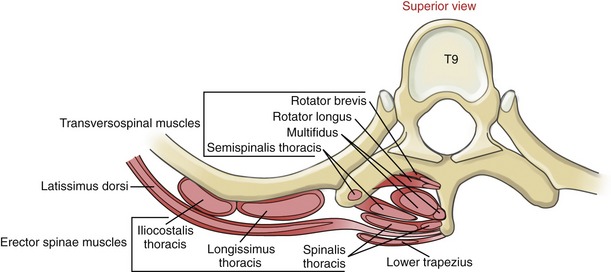
Transversospinal Muscles: Located immediately deep to the erector spinae muscles is the transversospinal muscle group: the semispinalis, multifidi, and rotatores (Figures 10-9 and 10-10). Semispinalis muscles are located superficially; the multifidi, intermediately; and the rotatores, deeply.
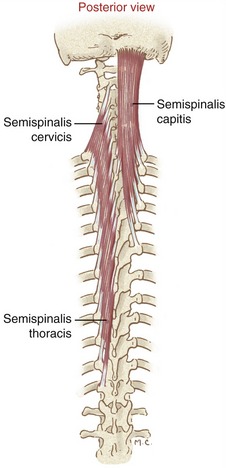
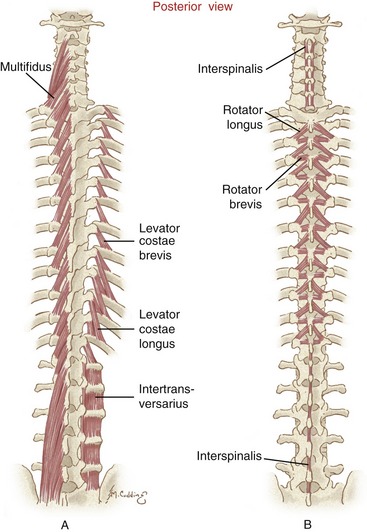
FIGURE 10-10. A posterior view shows the deeper muscles within the transversospinal group (multifidi on entire left side of A; rotatores bilaterally in B). The muscles within the short segmental group (intertransversarius and interspinalis) are depicted in A and B, respectively. Note that intertransversarius muscles are shown for the right side of the lumbar region only. The levator costarum muscles are involved with ventilation and are discussed in Chapter 11. (Modified from Luttgens K, Hamilton N: Kinesiology: scientific basis of human motion, ed 9, Madison, Wis, 1997, Brown and Benchmark.)
The name transversospinal refers to the general attachments of most of the muscles (i.e., from the transverse processes of one vertebra to the spinous processes of a more superiorly located vertebra). With a few exceptions, these attachments align most muscle fibers in a cranial-and-medial direction. Many of the muscles within the transversospinal group are morphologically similar, varying primarily in length and in the number of intervertebral junctions that each muscle crosses (Figure 10-11). Although somewhat oversimplified, this concept can greatly assist in learning the overall anatomy and actions of these muscles.
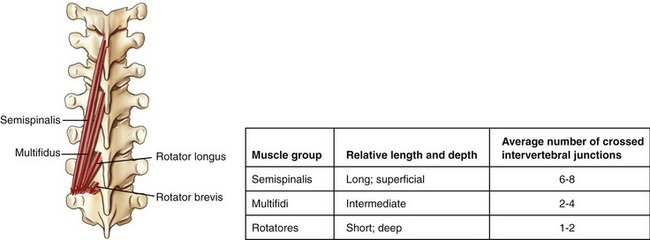
Semispinalis Muscles: The semispinalis muscles consist of the semispinalis thoracis, semispinalis cervicis, and semispinalis capitis (see Figure 10-9). In general, each muscle, or main set of fibers within each muscle, crosses six to eight intervertebral junctions. The semispinalis thoracis consists of many thin muscle fasciculi, interconnected by long tendons. Muscle fibers attach from transverse processes of T6 to T10 to spinous processes of C6 to T4. The semispinalis cervicis, much thicker and more developed than the semispinalis thoracis, attaches from upper thoracic transverse processes to spinous processes of C2 to C5. Muscle fibers that attach to the prominent spinous process of the axis (C2) are particularly well developed, serving as important stabilizers for the suboccipital muscles (described ahead).
The semispinalis capitis lies deep to the splenius and trapezius muscles. The muscle arises primarily from upper thoracic transverse processes. The muscle thickens superiorly as it attaches to a relatively large region on the occipital bone, filling much of the area between the superior and inferior nuchal lines (see Figure 9-3).
The semispinalis cervicis and capitis are the largest muscles that cross the posterior side of the neck. Their large size and near-vertical fiber direction account for the fact that these muscles provide 35% to 40% of the total extension torque of the craniocervical region.145 Right and left semispinalis capitis muscles are readily palpable as thick and round cords on either side of the midline of the upper neck, especially evident in infants and in thin, muscular adults (Figure 10-12).

Multifidi: Multifidi are situated just deep to the semispinalis muscles. The plural “multifidi” indicates a collection of multiple fibers, rather than a set of individual muscles. All multifidi share a similar fiber direction and length, extending between the posterior sacrum and the axis (C2).6,23,132 In general, the multifidi originate from the transverse process of one vertebra and insert on the spinous process of a vertebra located two to four intervertebral junctions above (see Figure 10-10, A).
Multifidi are thickest and most developed in the lumbosacral region (see multiple attachments listed in Box 10-3).100 The overlapping fibers of the multifidi fill much of the concave space formed between the spinous and transverse processes. The multifidi provide an excellent source of extension torque and associated stability to the base of the spine. Excessive force in the lumbar multifidi—from either active contraction or protective spasm—may be expressed clinically as an exaggerated lordosis.
Rotatores: The rotatores are the deepest of the transversospinal group of muscles. Like the multifidi, the rotatores consist of a large set of individual muscle fibers. Although the rotatores exist throughout the entire vertebral column, they are best developed in the thoracic region (see Figure 10-10, B).132 Each fiber attaches between the transverse process of one vertebra and the lamina and base of the spinous process of a vertebra located one or two intervertebral junctions above. By definition, the rotator brevis spans one intervertebral junction, and the rotator longus spans two intervertebral junctions.
Summary: On average, the transversospinal muscles cross fewer intervertebral junctions than the erector spinae group. This feature suggests that, in general, the transversospinal muscles favor a design for producing relatively fine controlled movements and stabilizing forces across the axial skeleton.15,22
Contracting bilaterally, the transversospinal muscles extend the axial skeleton (see Figure 10-8, B). Increased extension torque exaggerates the cervical and lumbar lordosis and decreases the thoracic kyphosis. The size and thickness of the transversospinal muscles are greatest at either end of the axial skeleton. Cranially, the semispinalis cervicis and capitis are very well-developed extensors of the craniocervical region; caudally, the multifidi are very well-developed extensors of the lower lumbar region, accounting for two thirds of the muscular-based stability in this region.154
Contracting unilaterally, the transversospinal muscles laterally flex the spine; however, their leverage for this action is limited because of their close proximity to the vertebral column. The more obliquely oriented transversospinal muscles assist with contralateral axial rotation. From a relatively fixed transverse process, contraction of a single left multifidus or rotator longus, for example, can rotate a superiorly located spinous process toward the left and, as a result, rotate the anterior side of the vertebra to the right. Compared with all the trunk muscles, however, the transversospinal muscles are secondary axial rotators. The leverage for this rotation is relatively poor because of the muscle’s proximity to the vertebral column. (Compare the multifidi with the obliquus abdominis externus, for example, in Figure 10-3, C). Furthermore, the prevailing line of force of most transversospinal muscle fibers is directed more vertically than horizontally, thereby providing a greater force potential for extension than for axial rotation.
Short Segmental Group of Muscles: The short segmental group of muscles consists of the interspinalis and the intertransversarius muscles (see Figure 10-10). (The plural “interspinales and intertransversarii” is often used to describe all the members within the entire set of these muscles.) They lie deep to the transversospinal group of muscles. The name “short segmental” refers to the extremely short length and highly segmented organization of the muscles. Each individual interspinalis or intertransversarius muscle crosses just one intervertebral junction. These muscles are most developed in the cervical region, where fine control of the head and neck is so critical.132
Each right and left pair of intertransversarius muscles is located between adjacent transverse processes. The anatomy of the intertransversarii as a group is more complex than that of the interspinales.132 In the cervical region, for example, each intertransversarius muscle is divided into small anterior and posterior muscles, between which pass the ventral rami of spinal nerves.
Summary: The highly segmented nature of the interspinalis and intertransversarius muscles is ideal for fine motor control of the axial skeleton. Because these unisegmental muscles possess a relatively high density of muscle spindles, they likely provide the nervous system (and therefore other muscles) a rich source of sensory feedback, especially in the craniocervical region.23
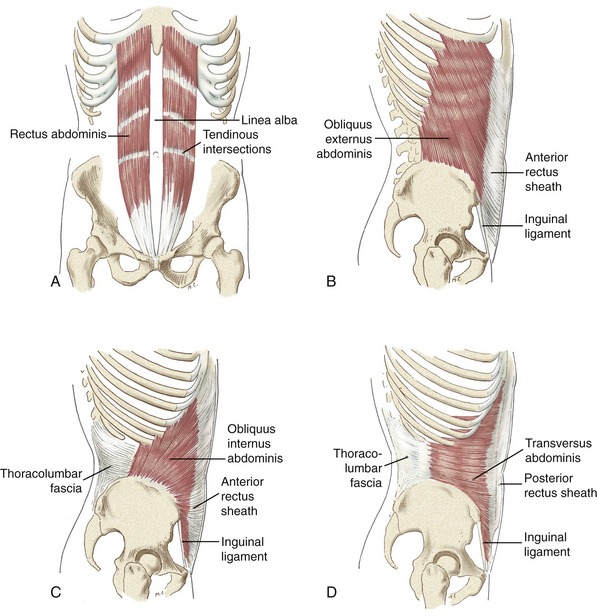
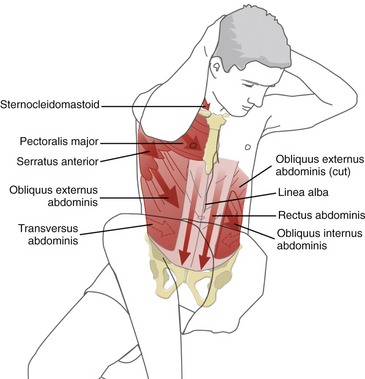
SET 2: MUSCLES OF THE ANTERIOR-LATERAL TRUNK (“ABDOMINAL” MUSCLES)
The muscles of the anterior-lateral trunk include the rectus abdominis, obliquus externus abdominis, obliquus internus abdominis, and transversus abdominis (Figure 10-13). As a group, these muscles are often collectively referred to as the “abdominal” muscles. The rectus abdominis is a long straplike muscle located on both sides of the midline of the body. The obliquus externus abdominis, obliquus internus abdominis, and transversus abdominis—the lateral abdominals—are wide and flat, layered superficial to deep, across the anterior-lateral aspects of the abdomen.
The abdominal muscles have several important physiologic functions, including supporting and protecting abdominal viscera and increasing intrathoracic and intra-abdominal pressures. As will be further described in Chapter 11, increasing the pressures in these cavities assists with functions such as forced expiration of air from the lungs, coughing, defecation, and child birth. This chapter focuses more on the kinesiologic functions of the abdominal muscles.
Formation of the Rectus Sheaths and Linea Alba: The obliquus externus abdominis, obliquus internus abdominis, and transversus abdominis muscles from the right and left sides of the body fuse at the midline of the abdomen through a blending of connective tissues. Each muscle contributes a thin bilaminar sheet of connective tissue that ultimately forms the anterior and posterior rectus sheaths. As depicted in Figure 10-14, the anterior rectus sheath is formed from connective tissues from the obliquus externus abdominis and the obliquus internus abdominis muscles. The posterior rectus sheath is formed from connective tissues from the obliquus internus abdominis and transversus abdominis. Both sheaths surround the vertically oriented rectus abdominis muscle and continue medially to fuse with identical connective tissues from the other side of the abdomen. The connective tissues thicken and crisscross as they traverse the midline, forming the linea alba (the Latin word linea means “line,” and albus, “white”). The linea alba runs longitudinally between the xiphoid process and pubic symphysis and pubic crest.
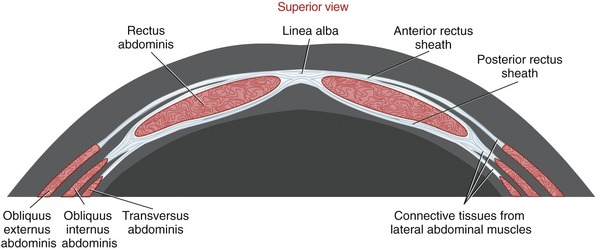
Anatomy of the Abdominal Muscles: The rectus abdominis muscle consists of right and left halves, separated by the linea alba. Each half of the muscle runs longitudinally, widening as it ascends within an open sleeve formed between the anterior and posterior rectus sheaths. The muscle is intersected and reinforced by three fibrous bands, known as tendinous intersections. These bands blend with the anterior rectus sheath. The rectus abdominis arises from the region on and surrounding the crest of the pubis, and it attaches superiorly on the xiphoid process and cartilages of the fifth through seventh ribs.
The anatomic organization of the obliquus externus abdominis, obliquus internus abdominis, and transversus abdominis muscles is different from that of the rectus abdominis. As a group, the more laterally placed muscles originate laterally or posterior-laterally on the trunk and run in a different direction toward the midline, eventually blending with the linea alba and contralateral rectus sheaths (Table 10-3).
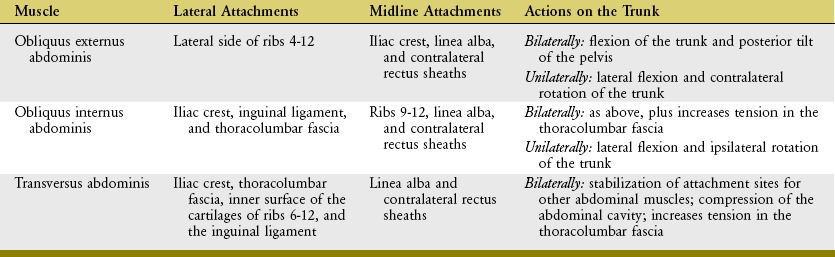
The obliquus externus abdominis (informally referred to as the “external oblique”) is the largest and most superficial of the lateral abdominal muscles. The external oblique muscle travels in an inferior-and-medial direction, similar to the direction of the hands placed diagonally in front pockets of pants. The obliquus internus abdominis (or less formally the “internal oblique”) is located immediately deep to the external oblique muscle, forming the second layer of the lateral abdominals. Fibers originate from the iliac crest and, to a varying degree, blend with the adjacent thoracolumbar fascia. From this lateral attachment point, the fibers course in a cranial-and-medial direction toward the linea alba and lower ribs. As evident in Figure 10-13, C, the inferior attachments of the internal oblique muscle extend to the inguinal ligament. The average fiber direction of the internal oblique muscle is nearly perpendicular to the average fiber direction of the overlying external oblique muscle.
The transversus abdominis is the deepest of the abdominal muscles. The muscle is also known as the “corset muscle,” reflecting its role in compressing the abdomen as well as stabilizing the lower back through attachments into the thoracolumbar fascia.132 Of all the abdominal muscles, the transversus abdominis has the most extensive and consistent attachments into the thoracolumbar fascia,136 followed closely by the internal oblique muscle.
Actions of the Abdominal Muscles: Bilateral action of the rectus abdominis and oblique abdominal muscles reduces the distance between the xiphoid process and the pubic symphysis. Depending on which body segment is the most stable, bilateral contraction of these abdominal muscles flexes the thorax and upper lumbar spine, posteriorly tilts the pelvis, or both. Figure 10-15 depicts a diagonally performed sit-up maneuver that places a relatively large demand on the oblique abdominal muscles. During a standard sagittal plane sit-up, however, the opposing axial rotation and lateral flexion tendencies of the various abdominal muscles are neutralized by opposing right and left muscles.
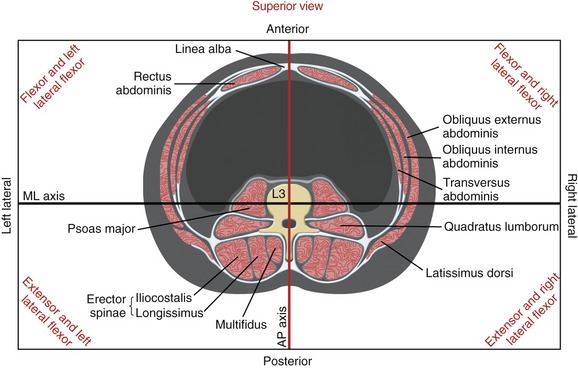
As described in Chapter 9, the axes of rotation for all motions of the vertebral column are located in the region of the interbody joints. The relative posterior placement of the axes relative to the trunk equips the abdominal muscles, most notably the rectus abdominis, with very favorable leverage for generating trunk flexion torque (Figure 10-16). Note in Figure 10-16 that, with the exception of the psoas major, all muscles have a moment arm to produce torques in both sagittal and frontal planes.
Contracting unilaterally, the abdominal muscles laterally flex the trunk. The external and internal obliques are particularly effective in this action owing to their relatively favorable leverage (i.e., long moment arms) (see Figure 10-16) and, as a pair, relatively large cross-sectional area. The combined cross-sectional area of the external and internal obliques at the level of the L4-L5 junction is almost twice that of the rectus abdominis muscle.106
Lateral flexion of the trunk often involves activation of both trunk flexor and extensor muscles. For example, lateral flexion against resistance to the right demands a contraction from the right external and internal oblique, right erector spinae, and right transversospinal muscles. Coactivation amplifies the total frontal torque while simultaneously stabilizing the trunk within the sagittal plane.12
By far, the internal and external oblique muscles are the most effective axial rotators of the trunk.7,12,73,141 The external oblique muscle is a contralateral rotator, and the internal oblique muscle is an ipsilateral rotator. The strong axial rotation potential of these muscles reflects their relatively large cross-sectional area and favorable leverage (see Figure 10-3, C for long moment arm length of the obliquus externus abdominis). During active axial rotation in a particular direction, the external oblique muscle on one side functions synergistically with the internal oblique on the other side.141 This functional synergy produces a diagonal line of force that crosses the midline through the muscles’ mutual attachment into the linea alba (see Figure 10-15). Contraction of the two muscles therefore reduces the distance between one shoulder and the contralateral iliac crest.
Several electromyographic (EMG) studies using intramuscular (fine-wire) electrodes demonstrate some degree of bilateral activation of the transversus abdominis during axial rotation.38,82,141 It has been shown that during axial rotation the middle and lower fibers of the transversus abdominis coactivate at slightly different times than the upper fibers.141 Although the exact role of the transversus abdominis during axial rotation is uncertain, the muscle appears to function more as a stabilizer for the oblique abdominal muscles than a torque generator of axial rotation. Bilateral activation of the transversus abdominis can stabilize the ribs, linea alba, and thoracolumbar fascia—areas that serve as attachments for the internal or external oblique muscles.
The torque demands placed on the axial rotators of the trunk vary considerably based on the nature of an activity and position of the body.12 Torque demands are relatively large during high-power axial rotations, such as sprinting, wrestling, and throwing a discus or javelin. The demands may be very low, however, during activities that involve slow twisting of the trunk while in an upright position, such as during walking over level surfaces.
SPECIAL FOCUS 10-2  Role of Trunk Extensors as “Rotational Synergists” to the Oblique Abdominal Muscles
Role of Trunk Extensors as “Rotational Synergists” to the Oblique Abdominal Muscles
The external and internal oblique muscles are the primary axial rotators of the trunk. Secondary axial rotators include the ipsilateral latissimus dorsi, the more oblique components of the ipsilateral iliocostalis lumborum, and the contralateral transversospinal muscles. These secondary axial rotators are also effective extensors of the trunk. During a strong axial rotation movement, these extensor muscles are able to offset or neutralize the potent trunk flexion potential of the oblique abdominal muscles.98,154 Without this neutralizing action, a strenuous action of axial rotation would automatically be combined with flexion of the trunk. The aforementioned extensor muscles resist the flexion tendency of the oblique abdominal muscles, but also contribute slightly to the axial rotation torque.
The multifidi muscles provide a particularly important element of extension stability to the lumbar region during axial rotation.128,156 Pathology involving the apophyseal joints or discs in the lumbar region may be associated with weakness, fatigue, or reflexive inhibition of these muscles. Without adequate activation from the multifidi during axial rotation, the partially unopposed oblique muscles would, in theory, create a subtle and undesirable flexion bias to the base of the spine.
Comparing Trunk Flexor versus Trunk Extensor Peak Torque: In the healthy adult, on average, the magnitude of a maximal-effort trunk flexion torque is typically less than maximal-effort trunk extension torque. Although data vary based on gender, age, history of back pain, and angular velocity of the testing device, the flexor-to-extensor torque ratios determined isometrically for the trunk and craniocervical regions are between 0.45 and 0.77.20,80,114,145 Although the trunk flexor muscles normally possess greater leverage for sagittal plane torque (see Figure 10-16), the trunk extensor muscles possess greater mass and, equally important, greater overall vertical orientation of muscle fibers.95,106 The typically greater torque potential of the trunk extensor muscles reflects the muscles’ predominant role in counteracting gravity, either for the maintenance of upright posture or for carrying loads in front of the body.
SET 3: ADDITIONAL MUSCLES (ILIOPSOAS AND QUADRATUS LUMBORUM)
Iliopsoas: The iliopsoas is a large muscle consisting of two parts: the iliacus and the psoas major (see Figure 12-27). As are most hip flexors, the iliopsoas is innervated by the femoral nerve, a large branch from the lumbar plexus. The iliacus has a proximal attachment on the iliac fossa and lateral sacrum, just anterior and superior to the sacroiliac joint. The psoas major attaches proximally to the transverse processes of T12 to L5, including the intervertebral discs. The two muscles fuse distal to the inguinal ligament and typically attach as a single tendon to the lesser trochanter of the femur.
The iliopsoas is a long muscle, exerting a potent kinetic influence across the trunk, lumbar spine, lumbosacral junction, and hip joints. Crossing anterior to the hip, it is a dominant flexor, drawing the femur toward the pelvis or the pelvis toward the femur. In the latter movement, the iliopsoas can anteriorly tilt the pelvis, a motion that increases the lordosis of the lumbar region (review in Figure 9-63, A). With muscular assistance from the abdominal muscles, a strong bilateral contraction of the iliopsoas can also rotate the pelvis and superimposed trunk over fixed femurs. Based on this ability, the iliopsoas is as much a respected trunk flexor as a hip flexor. This discussion resumes later in the chapter.
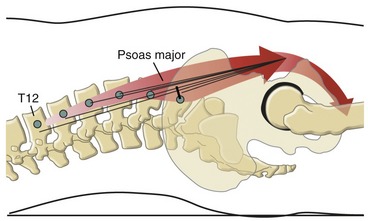
Function of the Psoas Major at the Lumbosacral Region: In the anatomic position the psoas major demonstrates leverage for lateral flexion of the lumbar spine (see Figure 10-16).95 Little, if any, leverage exists for axial rotation.
The flexor and extensor capacity of the psoas major differs throughout the lumbosacral region. Across the L5-S1 junction, the psoas major has an approximate 2-cm moment arm for flexion (Figure 10-17).107 The psoas major is therefore an effective flexor of the lower end of the lumbar spine relative to the sacrum. Progressing superiorly toward L1, however, the line of force of the psoas major gradually shifts slightly posterior, falling either through or just posterior to the multiple medial-lateral axes of rotation (see cross-section at L3 in Figure 10-16). The muscle’s location reduces or eliminates its flexor or extensor capacity. The psoas major therefore is neither a dominant flexor nor extensor of the lumbar region, but rather a dominant vertical stabilizer of the region.57,127 (The term “vertical stabilizer” describes a muscular function of stabilizing a region of the axial skeleton in a near-vertical position while maintaining its natural physiologic curve.) Because of the lack of effective leverage in the lumbar region, the psoas major has a minimal role in directly influencing the degree of lordosis.127 The iliopsoas, however (as all hip flexor muscles), can indirectly increase the lordotic posture of the lumbar spine by tilting the pelvis anteriorly across the hip joints.
Quadratus Lumborum: Anatomically, the quadratus lumborum is considered a muscle of the posterior abdominal wall. The muscle attaches inferiorly to the iliolumbar ligament and iliac crest and superiorly to the twelfth rib and the tips of the stout transverse processes of L1 to L4 (Figure 10-18). The relative thickness of the muscle is evident by viewing Figure 10-16. The quadratus lumborum is innervated by the ventral rami of spinal nerves T12-L3.
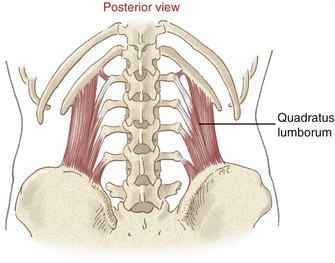
Contracting bilaterally, the quadratus lumborum is an extensor of the lumbar region. Its action is based on the line of force passing about 3.5 cm posterior to the medial-lateral axis of rotation at L3.107
Contracting unilaterally, the quadratus lumborum has very favorable leverage as a lateral flexor of the lumbar region.57 The axial rotation potential of the quadratus lumborum, however, is minimal.
The psoas major and the quadratus lumborum run nearly vertically on both sides of the lumbar vertebrae (see Figure 10-16). A strong bilateral contraction of these muscles affords excellent vertical stability throughout the lumbar spine, including the L5-S1 junction. Theoretically, exercises that increase the volitional control and conditioning of these muscles may benefit a person with pain related to instability of the lumbar region.
Muscles of the Trunk: Functional Interactions among Muscles
Thus far in this chapter, the discussion of the muscles of the trunk has focused primarily on their anatomy and, for the most part, individual actions (Table 10-4). The upcoming discussion pays more attention to the functional interactions among the muscles or muscle groups. Two themes are explored: (1) muscular-based stability of the trunk, and (2) muscular kinesiology of performing a standard sit-up movement. The second interaction exemplifies a classic kinesiologic relationship between the trunk and hip muscles.
TABLE 10-4.
Actions of Most Muscles of the Trunk
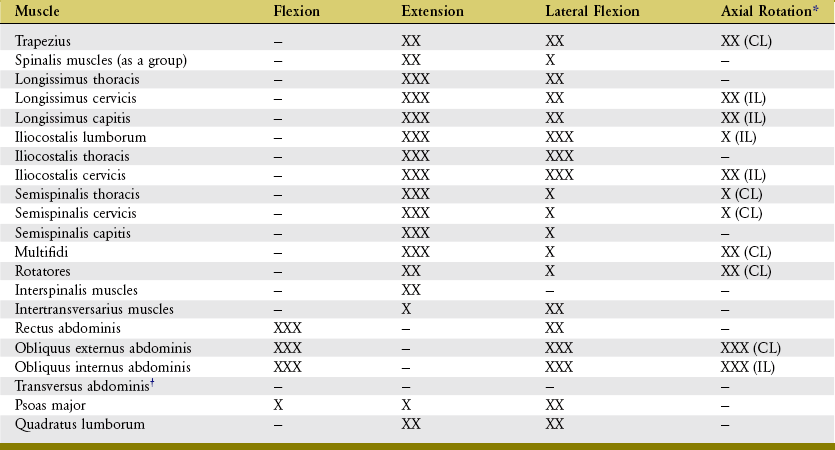
*CL, contralateral rotation; IL, ipsilateral rotation.
†Acts primarily to increase intra-abdominal pressure and, via attachments to the thoracolumbar fascia, to stabilize the lumbar region. Also stabilizes the attachment sites for the other lateral abdominal muscles.
MUSCULAR-BASED STABILITY OF THE TRUNK
Active muscle force provides the primary mechanism for stabilizing the axial skeleton, including the trunk.12,28,84,148 Although ligaments and other connective tissues provide a secondary source of this stability, only muscles can adjust both the magnitude and timing of their forces.
Muscular-based stability of the trunk is often referred to as “core stability.” Such stability ensures a near-static posture of the trunk even under the influence of destabilizing external forces.16 Consider, for example, the wave of muscular activation experienced throughout the trunk when one attempts to stand or sit upright in an accelerating bus or train. Normally, trunk muscles are able to subconsciously stabilize the position of the trunk relative to the surrounding environment and, equally important, to stabilize the individual spinal segments within the axial skeleton. Ideally, a stable trunk optimizes postural alignment and limits excessive, and potentially stressful, micromotions between intervertebral junctions. Finally, stability of the trunk also establishes a firm base for muscles to move the limbs.
Intrinsic Muscular Stabilizers of the Trunk: The intrinsic muscular stabilizers of the trunk include the transversospinal and short segmental groups of muscles. These deep and relatively short muscles are depicted in a highly diagrammatic fashion in Figure 10-19, A. In general, these muscles stabilize the spine by controlling the precise alignment and stiffness among a relatively few intervertebral junctions at a time. The relative high density of muscle spindles residing in many of these segmental muscles enhances their fine-tuning ability.113
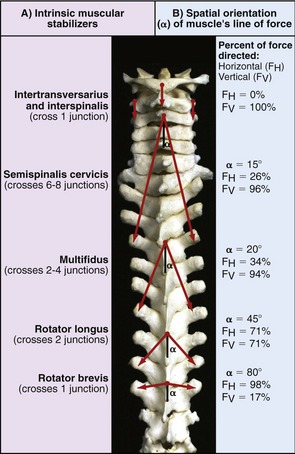
As indicated in Figure 10-19, B, the spatial orientation of each muscle’s line of force (depicted by α) produces a unique stabilization effect on the vertebral column. Vertically running interspinalis and intertransversarius muscles produce 100% of their force in the vertical direction (FV). In contrast, the near-horizontally oriented rotator brevis muscle produces close to 100% of its force in the horizontal direction (FH). All of the remaining muscles produce forces that are directed diagonally, at some angle between 0 and 90 degrees. The muscles act as an array of bilaterally matched guy wires, specifically aligned to compress as well as control the shear between intervertebral junctions. In addition to effectively securing both vertical and horizontal stability, collectively these muscles exert extension, lateral flexion, and axial rotation torques across the entire vertebral column. Without such fine muscle control, the multi-segmented vertebral column becomes very vulnerable to exaggerated spinal curvature, excessive interspinal mobility, and, in some cases, painful instability.
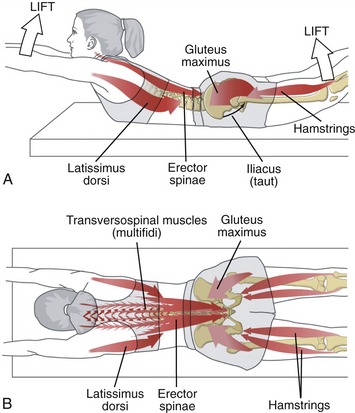
Extrinsic Muscular Stabilizers of the Trunk: The primary extrinsic muscular stabilizers of the trunk include the abdominal muscles, erector spinae, quadratus lumborum, psoas major, and the hip muscles (by connecting the lumbopelvic region with the lower extremities). These relatively long and often thick muscles stabilize the trunk by creating a strong and semirigid link between the cranium, vertebral column, pelvis, and lower extremities.28 Because many of these muscles cross a broad region of the body or trunk, they likely provide relatively coarse control over trunk stability. In addition, because many of these muscles possess a sizable cross-sectional area and leverage, they are, as a group, also important torque generators for the trunk and adjacent hip joints.22
To further illustrate the potential role of the extrinsic stabilizers, Figure 10-20 shows a person activating his external muscular stabilizers in response to an impending external perturbation. Note the concentration of muscular activity in the lower region of the trunk. Activation of the psoas major, quadratus lumborum, erector spinae, and abdominal muscles provides substantial stability to the lumbopelvic regions, in all three planes. Strong activation of abdominal muscles also helps to increase intra-abdominal pressure—a mechanism believed to exert a stabilizing effect throughout the lumbar region.67 The horizontally disposed transversus abdominis, in particular, creates a circumferential splinting effect across the entire low back region, including the sacroiliac joints.
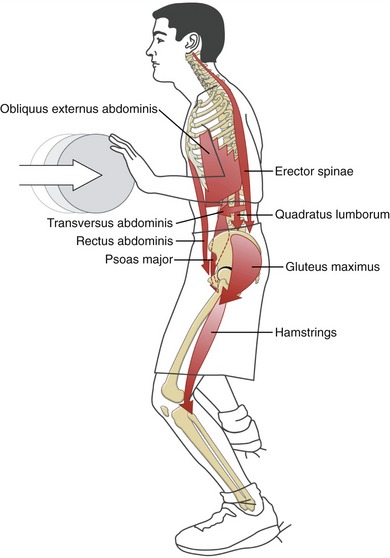
In closing, it should be pointed out that although the external and internal muscular stabilizers have been presented separately, in reality there is a large overlap and redundancy in their functions. This may be appreciated by mentally superimposing the muscular arrows depicted in both Figures 10-19 and 10-20. In ideal health, all muscles of the trunk contribute to the stabilization of the trunk, in both static and dynamic conditions.28,36,135,147,148 The specific strategy used by any single muscle differs, however, based on factors such as its depth, morphology, spatial orientation, and skeletal or connective tissue attachments.
CLINICAL CONNECTION 10-2  Therapeutic Exercise as a Way to Improve Lumbopelvic Stability: a Brief Overview
Therapeutic Exercise as a Way to Improve Lumbopelvic Stability: a Brief Overview
A significant percentage of the stress-related musculoskeletal pathology of the trunk occurs at the lumbopelvic region. This region includes the lumbar spine, lumbosacral junction, and the sacroiliac joints. The term lumbopelvic instability has evolved to describe a painful, usually nonspecific, condition that is associated with hypermobility at one or more of the articulated segments.156 The amount of hypermobility may be slight and difficult to quantify through routine clinical assessment. This condition, nevertheless, is believed capable of generating excessive stress on spinal-related structures, including the interbody, apophyseal joint and sacroiliac joints, spinal ligaments, and neural tissues. Persons often seek medical attention when pain occurs with movement in the region. The clinical picture of this condition is often complicated by the uncertainty regarding whether lumbopelvic instability is the cause or the effect of other impairments in the low back, such as degenerative disc disease.*
Weakness, fatigue, or the inability to specifically control the timing or magnitude of forces produced by the trunk muscles has long been suspected as a potential cause, or at least an associated factor, in the pathogenesis of lumbopelvic instability. For this reason, specific muscular-based exercises are often considered an essential component of conservative treatment for this condition. It is beyond the scope of this chapter to describe the details or varying effectiveness of the many types of exercises designed to improve muscular-based lumbopelvic stability; this information can be found in other sources.† The following box, however, lists four themes that tend to be emphasized with this therapeutic approach. It is important to note that exercises of any kind may not be appropriate in cases of specific structural instability of the lumbopelvic region (such as acute or significant spondylolisthesis), acute disc herniation, or any conditions involving marked pain or deteriorating neurologic symptoms.
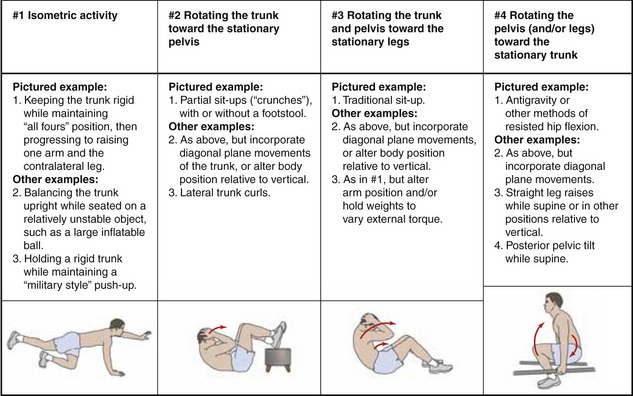
PERFORMING A STANDARD SIT-UP MOVEMENT
In addition to being a very important functional activity, the full sit-up is often performed as a way to strengthen the abdominal muscles. The common goal of the resistive exercise is to increase the strength and control of these muscles, often as a way to improve overall stability of the trunk. In a very broad sense, the strategies used to strengthen abdominal muscles usually fall into one of four categories (Figure 10-21). In column 1 of Figure 10-21, the abdominal muscles contract to produce an isometric force to maintain a near-constant distance between the xiphoid process and the anterior pelvis. In columns 2 to 4, the abdominal muscles contract to reduce the distance between the xiphoid process and the anterior pelvis. (By acting eccentrically, the same muscles could also be challenged to slowly resist an increase in distance between these two regions of the body.) Of the examples illustrated in Figure 10-21, perhaps the most traditional exercise, at least historically, is the standard sit-up, depicted in column 3.48
A full sit-up performed in a bent-knee position can be divided into two phases. The trunk flexion phase terminates when both scapulae are raised off the mat (Figure 10-22, A). The later hip flexion phase involves an additional 70 to 90 degrees of combined lumbar flexion and pelvic-on-femoral (hip) flexion (see Figure 10-22, B).
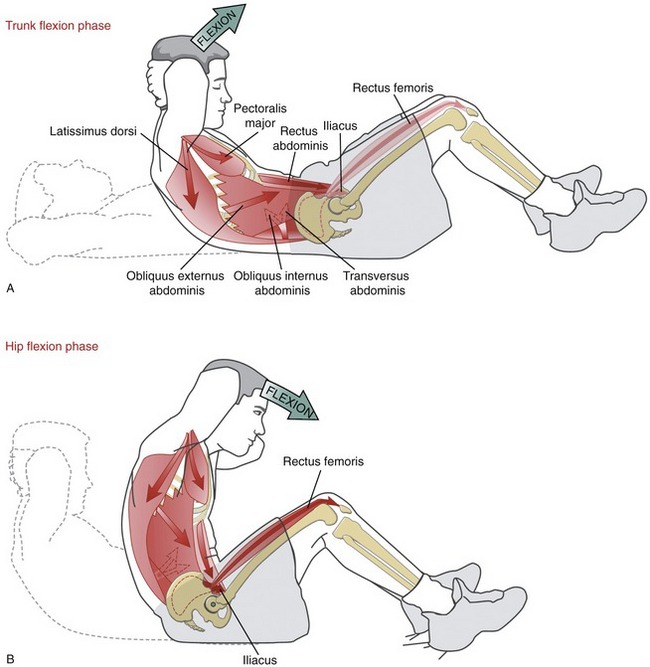
As depicted in Figure 10-22, A, the trunk flexion phase is driven primarily by contraction of the abdominal muscles, most notably the rectus abdominis.8,48 Contraction of these muscles flexes the thoracolumbar spine and tilts (rotates) the pelvis posteriorly, thereby flattening the lumbar spine. The EMG level of the hip flexor muscles is relatively low during the trunk flexion phase, regardless of the position of the hips and knees.8,48 Partially flexing the hips before performing the exercise releases passive tension in the hip flexor muscles while simultaneously increasing the passive tension in the gluteus maximus. These combined effects may assist the abdominal muscles in maintaining a posteriorly tilted pelvis.
Finally, as illustrated in Figure 10-22, A, the latissimus dorsi, by passing anterior to the upper thoracic spine, may assist in flexing this region of the thorax; the sternal head of the pectoralis major may assist in advancing the upper extremities toward the pelvis.
During the hip flexion phase of the sit-up, the pelvis and trunk rotate toward the femurs. The hip flexion phase is marked by stronger active contraction of the hip flexor muscles.48 Although any hip flexor muscle can assist with this action, Figure 10-22, B shows the iliacus and rectus femoris as the active participants. Relative levels of EMG from the iliacus, sartorius, and rectus femoris are significantly greater when the legs are actively held fixed to the supporting surface.8 The axis of rotation during the hip flexion phase of the full sit-up shifts toward the hip joints. Depending on technique, the abdominal muscles may continue to contract strongly or remain isometrically active. Their activation, however, does not contribute to hip (pelvic-on-femoral) flexion; rather, these muscles hold the flexed thoracolumbar region firmly against the rotating pelvis.
Persons with moderately weakened abdominal muscles typically display a characteristic posture when attempting to perform a full sit-up. Throughout the attempt, the hip flexor muscles dominate the activity. As a result there is minimal thoracolumbar flexion and excessive and “early” pelvic-on-femoral (hip) flexion. The dominating contraction of the hip flexor muscles exaggerates the lumbar lordosis, especially during the initiation of the maneuver.86
SPECIAL FOCUS 10-3  Comparing the Abdominal “Crunch” Exercise with the Standard Full Sit-up
Comparing the Abdominal “Crunch” Exercise with the Standard Full Sit-up
The early, trunk flexion phase of the full sit-up (depicted in Figure 10-22, A) is similar in many respects to the popular, and often recommended, “crunch” exercise for strengthening the abdominal muscles. Both the crunch and the full sit-up place significant and clinically challenging demands on the abdominal muscles as a whole.56 Differences exist, however, as the crunch places relatively greater demands on the rectus abdominis, whereas the full sit-up places relatively greater demands on the oblique musculature. Furthermore, when compared with a full (bent-knee) sit-up, the crunch exercise (as depicted in Figure 10-22, A) places only marginal demands on the hip flexor muscles. Perhaps the most clinically significant difference in the two exercises is the fact that the crunch exercise involves only marginal amounts of flexion in the lumbar spine, reported to be only 3 degrees.120 This is strikingly less than the lumbar flexion that accompanies a full (bent-knee) sit-up. The flexion of the lumbar spine during the full-sit up can create greater pressure on the discs (see Chapter 9). The crunch exercise therefore may be more appropriate than the full sit-up in persons with a history of disc pathology. This precaution appears prudent, especially considering that the crunch exercise still places significant demands on the abdominal muscles.
Muscles of the Craniocervical Region: Anatomy and Their Individual Actions
The following sections describe the anatomy and individual actions of the muscles that act exclusively within the craniocervical region. Musculature is divided into two sets: (1) muscles of the anterior-lateral craniocervical region and (2) muscles of the posterior craniocervical region (review Table 10-1).
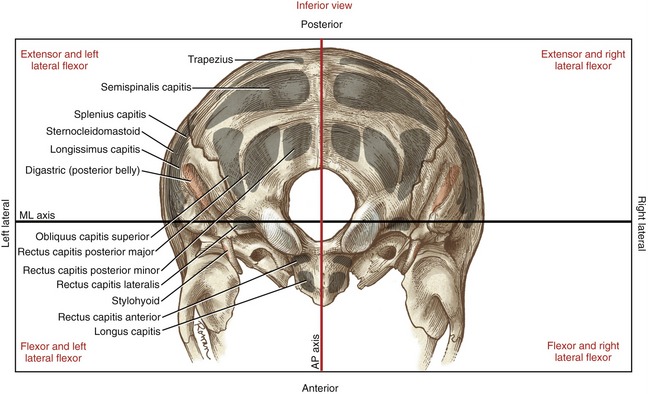
Figure 10-23 serves as an introduction to the potential actions of many muscles in the craniocervical region. The illustration depicts selected muscles as flexors or extensors, or right or left lateral flexors, depending on their attachment relative to the axes of rotation through the atlanto-occipital joints. Although Figure 10-23 describes the muscle actions at the atlanto-occipital joint only, the relative position of the muscles provides a useful guide for an understanding of the actions at other joints within the craniocervical region. This figure is referenced throughout the upcoming sections.
SET 1: MUSCLES OF THE ANTERIOR-LATERAL CRANIOCERVICAL REGION
The muscles of the anterior-lateral craniocervical region are listed in Box 10-4. With the exception of the sternocleidomastoid, which is innervated primarily by the spinal accessory nerve (cranial nerve XI), the muscles in this region are innervated by small unnamed nerves that branch from the ventral rami of the cervical plexus.
Sternocleidomastoid: The sternocleidomastoid is typically a prominent muscle located superficially on the anterior aspect of the neck. Inferiorly the muscle attaches by two heads: the medial (sternal) and lateral (clavicular) (Figure 10-24). From this attachment, the muscle ascends obliquely across the neck to attach to the cranium, specifically between the mastoid process of the temporal bone and the lateral half of the superior nuchae line.
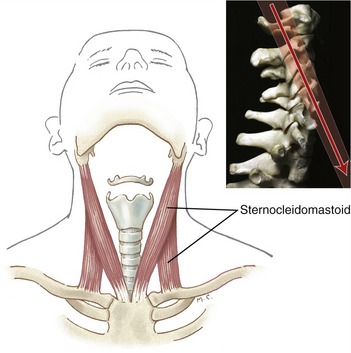
Acting unilaterally, the sternocleidomastoid is a lateral flexor and contralateral axial rotator of the craniocervical region. Contracting bilaterally, a pair of sternocleidomastoid muscles can flex or extend the craniocervical region depending on the specific area. Evident from a lateral view of a neutral cervical spine, the line of force of the right sternocleidomastoid is directed across the neck in an oblique fashion (see Figure 10-24, inset). Below approximately C3, the sternocleidomastoid crosses anterior to the medial-lateral axes of rotation; above C3, however, the sternocleidomastoid crosses just posterior to the medial-lateral axes of rotation.145 Acting together, the sternocleidomastoid muscles provide a strong flexion torque to the mid-to-lower cervical spine and a minimal extension torque to the upper cervical spine, including the atlanto-axial and atlanto-occipital joints.
Computer models predict that the sagittal plane torque potential of the different regions of the sternocleidomastoid is strongly affected by the initial posture of the craniocervical region.145 Primarily because of moment arm changes, the position of flexion of the mid-to-lower cervical spine, for example, nearly doubles the muscle’s flexion torque potential in this region. This becomes especially relevant in persons with an established marked forward head posture, referred to as protraction of craniocervical region in Chapter 9 (see Figure 9-47, A). Because this posture has greater flexion at the mid-to-lower cervical region, it may perpetuate the biomechanics that cause the forward head posture.
Scalenes: The scalene muscles attach between the tubercles of the transverse processes of the middle to lower cervical vertebrae and the first two ribs (Figure 10-25). (As a side note, the Latin or Greek root of the word scalene refers to a triangle with three unequal sides.) The specific attachments of these muscles are listed in Appendix III, Part C. The brachial plexus courses between the scalene anterior and scalene medius. Hypertrophy, spasm, or excessive stiffness of these muscles can compress the brachial plexus and cause motor and sensory disturbances in the upper extremity.
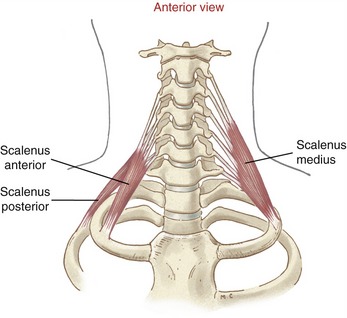
Contracting unilaterally, the scalene muscles laterally flex the cervical spine.29 Their axial rotation potential is likely limited because the muscle’s line of force nearly pierces the vertical axis of rotation. This topic remains controversial, however, with little scientific backing.39,65,83,86,122 The only rigorous study found on this subject concluded that the scalenes have a modest (5-degree) ipsilateral rotation function, at least when activated from the anatomic position.32 This conclusion, however, is difficult to confirm based on casual inspection of a human skeletal model, especially for the scalenus anterior. Further clarification is needed on the axial rotation function of the three scalene muscles. Their axial rotation function is likely highly dependent on the posture of the region and, even more important, on the starting position from which the muscles contract. It appears that an important function of the scalene muscles is their ability to return the craniocervical region to its near-neutral position from a fully rotated position. This more global and perhaps primary function may be overlooked when the neutral position is used as a starting point to analyze the muscles’ action.
Contracting bilaterally, the scalenus anterior and scalenus medius appear to have a limited moment arm to flex the cervical spine, particularly in the lower regions. The muscles’ bilateral activity is most likely related to ventilation (as described previously) and providing stability to the cervical region. The cervical attachments of all three scalene muscles split into several individual fasciculi (see Figure 10-25). Like a system of guy wires that stabilize a large antenna, the scalene muscles provide excellent bilateral and vertical stability to the middle and lower cervical spine. Fine control of the upper craniocervical region is more the responsibility of the shorter, more specialized muscles, such as the rectus capitis anterior and the suboccipital muscles (discussed ahead).
Longus Colli and Longus Capitis: The longus colli and longus capitis are located deep to the cervical viscera (trachea and esophagus), on both sides of the cervical column (Figure 10-26). These muscles function as a dynamic anterior longitudinal ligament, providing an important element of vertical stability to the region.49,87
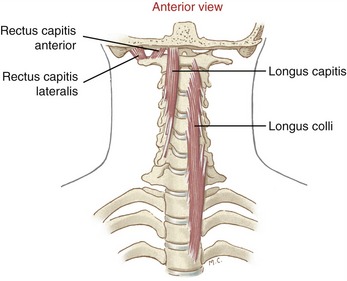
The longus capitis arises from the anterior tubercles of the transverse processes of the mid-to-lower cervical vertebrae and inserts into the basilar part of the occipital bone (see Figure 10-23). The primary action of the longus capitis is to flex and stabilize the upper craniocervical region. Lateral flexion is a secondary action.
Rectus Capitis Anterior and Rectus Capitis Lateralis: The rectus capitis anterior and rectus capitis lateralis are two short muscles that arise from the elongated transverse processes of the atlas (C1) and insert on the inferior surface of the occipital bone (see Figure 10-26). The rectus capitis lateralis attaches laterally to the occipital condyle; the rectus capitis anterior, the smaller of the recti, attaches immediately anterior to the occipital condyle (see Figure 10-23).
The actions of the rectus capitis anterior and lateralis muscles are limited to the atlanto-occipital joint; each muscle controls one of the joint’s two degrees of freedom (see Chapter 9). The rectus capitis anterior is a flexor, and the rectus capitis lateralis is a lateral flexor.
SPECIAL FOCUS 10-4  Soft-Tissue Whiplash Injury
Soft-Tissue Whiplash Injury
The soft tissues of the cervical spine are particularly vulnerable to injury from a whiplash event associated with an automobile accident. Whiplash associated with cervical hyperextension generally creates greater strain on soft tissues than does whiplash associated with cervical hyperflexion.133 Hyperextension occurs over a relatively large range of motion and therefore severely strains the craniocervical flexor muscles, cervical viscera, and other anteriorly located connective tissues, as well as excessively compressing the apophyseal joints and posterior elements of the cervical spine (Figure 10-27, A). In contrast, the maximum extent of flexion is partially blocked by the chin striking the chest (see Figure 10-27, B). Ideally, head restraints located within most automobiles help limit the extent of hyperextension and reduce injury from a collision.
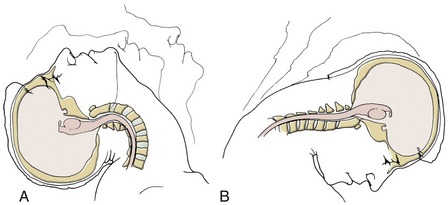
Hyperextension injuries tend to occur more often from rear-end impact automobile collisions. Careful measurements of human replicas and cadaver material show that immediately on contact the craniocervical region sharply retracts, followed by a more prolonged hyperextension.93,117 The brief retraction phase is usually completed before the cranium hits the head restraint. The anterior longitudinal ligament within the mid and lower cervical spine is particularly vulnerable to injury during this unprotected phase of the whiplash event.
The alar ligaments are particularly vulnerable to injury during the prolonged hyperextension phase of a rear-end collision, especially when the head is rotated at the time of the collision.44 Rotation of the head stretches the alar ligaments, which places them closer to their point of mechanical failure.
In addition, research has shown that the severe hyperextension associated with whiplash places excessive strain on flexor muscles, in particular the longus colli and longus capitis.108 In one study, a 56% strain (elongation) was measured in the longus colli—a level that can cause tissue damage. Often a person with a hyperextension injury shows a correlating pattern of marked tenderness and protective spasm in the region of the longus colli. Excessive strain in other muscles (such as the sternocleidomastoid and scalenus anterior) and the cervical viscera may also cause tenderness. Spasm in the longus colli tends to produce a relatively straight cervical spine, lacking the normal lordosis. Persons with a strained and painful longus colli often have difficulty shrugging their shoulders—an action produced primarily by the upper trapezius. When the longus colli and other flexors are too painful to fully contract, the upper trapezius muscle loses its stable cervical attachment and therefore becomes an ineffective elevator of the shoulder girdle. This clinical scenario is an excellent example of the interdependence of muscle function, in which one muscle’s action depends on the stabilization force of another.
SET 2: MUSCLES OF THE POSTERIOR CRANIOCERVICAL REGION
The muscles of the posterior craniocervical region are listed in Box 10-5. They are innervated by dorsi rami of cervical spinal nerves.
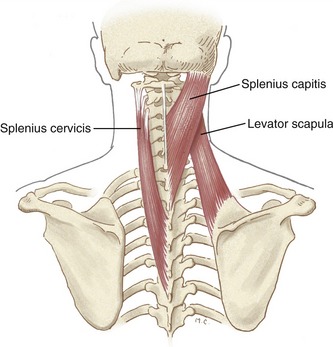
Splenius Cervicis and Capitis: The splenius cervicis and capitis muscles are a long and thin pair of muscles, named by their resemblance to a bandage (from the Greek splenion, bandage) (Figure 10-28). As a pair, the splenius muscles arise from the inferior half of the ligamentum nuchae and spinous processes of C7 to T6, just deep to the trapezius muscles. The splenius capitis attaches just posterior and deep to the sternocleidomastoid (see Figure 10-23). The splenius cervicis attaches to the posterior tubercles of the transverse processes of C1 to C3. Much of this cervical attachment is shared by the levator scapula muscle.
Suboccipital Muscles: The suboccipital muscles consist of four paired muscles located very deep in the neck, immediately superficial to the atlanto-occipital and atlanto-axial joints (Figure 10-29). These relatively short but thick muscles attach among the atlas, axis, and occipital bone. (Their specific muscular attachments are listed in Appendix III, Part C.)
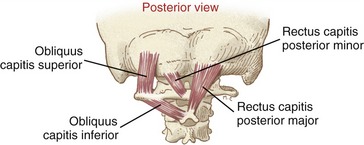
The suboccipital muscles are not easily palpable. They lie deep to the upper trapezius, splenius group, and semispinalis capitis muscles (see Figure 10-23). In conjunction with the rectus capitis anterior and lateralis, the suboccipital muscles are dedicated to providing precise control over the atlanto-occipital and atlanto-axial joints. This level of control is essential for optimal positioning of the eyes, ears, and nose. As indicated in Figure 10-30, each suboccipital muscle (plus each short rectus muscle) has a unique level of control and dominance over the joints of the upper craniocervical region.
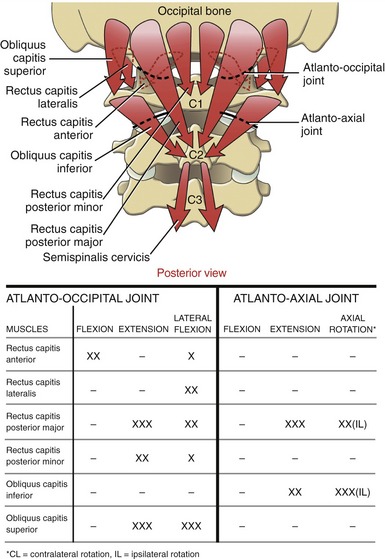
SPECIAL FOCUS 10-5  Specialized Muscles That Control the Atlanto-axial and Atlanto-occipital Joints: an Example of Fine-Tuning of Cervical Spinal Coupling
Specialized Muscles That Control the Atlanto-axial and Atlanto-occipital Joints: an Example of Fine-Tuning of Cervical Spinal Coupling
The specialized muscles listed in Figure 10-30 exert fine control over the movements of the upper craniocervical region. One benefit of this control is related to the spinal coupling pattern typically expressed within the cervical region. As described in Chapter 9, an ipsilateral spinal coupling pattern exists in the mid-and-lower cervical region between the motions of axial rotation and lateral flexion. Axial rotation, resulting primarily from the orientation of the facet surfaces within the apophyseal joints, is mechanically associated with slight ipsilateral lateral flexion, and vice versa. The expression of this coupling pattern can be obscured, however, by the action of the specialized muscles that control the atlanto-occipital and atlanto-axial joints. Consider, for example, right axial rotation of the craniocervical region. In order for a level horizontal visual gaze to be maintained throughout axial rotation, the left rectus capitis lateralis, for instance, produces a slight left lateral flexion torque to the head. This muscular action offsets the tendency of the head to laterally flex to the right with the rest of the cervical region during the right axial rotation. Similarly, right lateral flexion of the mid-to-lower cervical region (which is coupled with slight right axial rotation) may be accompanied by a slight offsetting left axial rotation torque applied to the head by the obliquus capitis inferior muscle. In both examples, the muscular actions allow the head and eyes to more precisely visually fix on an object.
Muscles of the Craniocervical Region: Functional Interactions among Muscles That Cross the Craniocervical Region
Nearly 30 pairs of muscles cross the craniocervical region. These include the muscles that act exclusively within the craniocervical region (Figure 10-30 and Table 10-5), plus those classified as muscles of the posterior trunk that cross the craniocervical region (e.g., trapezius and longissimus capitis).
TABLE 10-5.
Actions of Selected Muscles Located within the Craniocervical Region

CL, Contralateral rotation; IL, ipsilateral rotation.
*Upper parts of sternocleidomastoid extend the upper cervical region, atlanto-axial joint, and atlanto-occipital joint.
STABILIZING THE CRANIOCERVICAL REGION
The muscles that cross the craniocervical region comprise much of the bulk of the neck, especially in the regions lateral and posterior to the cervical vertebrae. When strongly activated, this mass of muscle serves to protect the cervical viscera and blood vessels, intervertebral discs, apophyseal joints, and neural tissues.87
Resistive or so-called “stabilization” exercises are often performed by athletes involved in contact sports as a means to hypertrophy this musculature. Hypertrophy alone, however, may not necessarily prevent neck injury. Data on the biomechanics of whiplash injury, for example, suggest that the time required to react to an impending injury and generate a substantial stabilizing force may exceed the time of the whiplash event.44 For this reason, athletes need to anticipate a potentially harmful situation and contract the neck musculature before impact. The timing of muscle contraction appears as important to protecting the neck as the magnitude of the muscle force.
In addition to protecting the neck, forces produced by muscles provide the primary source of vertical stability to the craniocervical region. The “critical load” of the cervical spine (i.e., maximum compressive load that the neck, unsupported by muscle, can sustain before buckling) is between 10.5 and 40 N (between 2.4 and 9 lb). Remarkably, this is less than the actual weight of the head.116,118 A coordinated interaction of craniocervical muscles generates forces that are, on average, directed nearly through the instantaneous axis of rotation at each intervertebral junction. By passing through or close to these multiple axes, the forces compress the vertebral segments together, thereby stabilizing them without buckling. The magnitude of these compression forces generated across the craniocervical region is quite high—nearly three times the weight of the head during the low-level muscle activation required to just balance the head during upright standing, and up to 23 times the weight of the head (or 1.7 times body weight) during maximal-effort muscle activation.109,118
Much of the muscular stabilization of the craniocervical region is accomplished by the relatively short, segmented muscles such as the multifidi, rotatores, longus colli and capitis, and interspinalis muscles. With relatively short fibers and multiple bony attachments, these muscles exert a fine, coordinated control of the stability in the region.25 This stability is augmented by other longer and typically thicker muscles, including the scalenes, sternocleidomastoid, levator scapula, semispinalis capitis and cervicis, and trapezius. When needed, these muscles form an extensive and strong guy-wire system that ensures vertical stability, most notably in frontal and sagittal planes. Figure 10-31, A highlights a sample of muscles that act as guy wires to maintain ideal anterior-posterior alignment throughout the craniocervical region. Ideally, the co-contraction of flexor and extensor muscles counterbalances, and as a consequence vertically stabilizes the region. Note that the muscles depicted in Figure 10-31, A are anchored inferiorly to several different structures: the sternum, clavicle, ribs, scapula, and vertebral column. These bony structures themselves must be stabilized by other muscles, such as the lower trapezius and subclavius, to secure the scapula and clavicle, respectively.
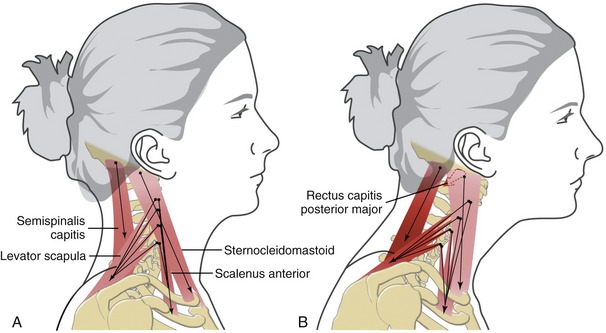
FIGURE 10-31. A, Four muscles are acting as guy wires to maintain ideal posture within the craniocervical region. B, Mechanics associated with a chronic forward head posture as discussed in Special Focus 10-6. The protracted position of the craniocervical region places greater stress on the levator scapula and semispinalis capitis muscles. The rectus capitis posterior major—one of the suboccipital muscles—is shown actively extending the upper craniocervical region. The highly active and stressed muscles are depicted in brighter red.
SPECIAL FOCUS 10-6  Muscular Imbalance Associated with Chronic Forward Head Posture
Muscular Imbalance Associated with Chronic Forward Head Posture
The ideal posture shown in Figure 10-31, A depicts an optimally balanced craniocervical “guy-wire” system. Excessive muscular tension in any of the muscles, however, can disrupt the vertical stability of the region. One such disruption is a chronic forward head posture, involving excessive protraction of the craniocervical region (see Figure 10-31, B). Habitual forward head posture can occur for at least two different reasons. First, severe hyperextension of the neck can injure anterior muscles, such as the sternocleidomastoid, longus colli, and scalenus anterior. As a result, chronic spasm in the excessively strained muscles translates the head forward, resulting in excessive flexion, especially at the cervicothoracic junction. A clinical sign often associated with forward head posturing is a realignment of the sternocleidomastoid within the sagittal plane. The cranial end of the muscle, normally aligned posterior to the sternoclavicular joint, shifts anteriorly with the head to a position directly above the sternoclavicular joint (compare Figure 10-31, A with B).
Regardless of the factors that predispose a person to a chronic forward head posture, the posture itself stresses extensor muscles, such as the levator scapula and semispinalis capitis (see Figure 10-31, B).108 A suboccipital muscle, such as the rectus capitis posterior major, may become fatigued as a result of its prolonged extension activity required to “level” the head and eyes. Over time, increased muscular stress throughout the entire craniocervical region can lead to localized and painful muscle spasms, or “trigger points,” common in the levator scapula and suboccipital muscles. This condition is often associated with headaches and radiating pain into the scalp and temporomandibular joints. The key to most treatment for chronic forward head posture is to restore optimal craniocervical posture,49 accomplished through improved postural awareness, ergonomic workplace design, therapeutic exercise, motor relearning, or specific manual therapy techniques.*
PRODUCING EXTENSIVE AND WELL-COORDINATED MOVEMENTS OF THE HEAD AND NECK: OPTIMIZING THE PLACEMENT OF THE EYES, EARS, AND NOSE
Figure 10-32 illustrates a total body movement that exhibits a sample of the muscular interactions used to maximize the extent of right axial rotation of the craniocervical region. Note that full axial rotation of the craniocervical region provides the eyes with well over 180 degrees of visual scanning. As depicted, rotation to the right is driven by simultaneous activation of the left sternocleidomastoid and left trapezius (see Figure 10-32, A); right splenius capitis and cervicis; right upper erector spinae, such as the longissimus capitis; and left transversospinal muscles, such as the multifidi (see Figure 10-32, B). Although not depicted, several suboccipital muscles (namely the right rectus capitis posterior major and right obliquus capitis inferior) are actively controlling atlanto-axial joint rotation.
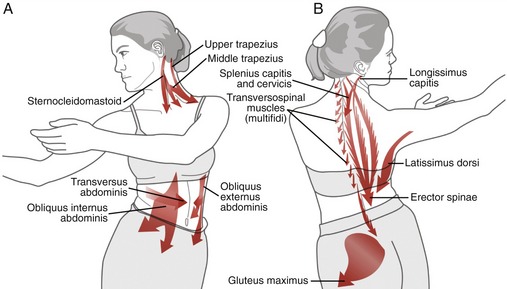
Full axial rotation of the craniocervical region requires muscular interactions that extend into the trunk and lower extremities. Consider, for example, the activation of the right and left oblique abdominal muscles (see Figure 10-32, A). They provide much of the torque needed to rotate the thoracic region, which serves as a structural foundation for the craniocervical region. Furthermore, as suggested by Figure 10-32, B, the erector spinae and transversospinal muscles are active throughout the posterior trunk to offset the potent trunk flexion tendency of the oblique abdominal muscles. The latissimus dorsi is an ipsilateral rotator of the trunk when the glenohumeral joint is well stabilized by other muscles.12 The left gluteus maximus is shown actively rotating the pelvis and attached lumbosacral region to the right, relative to the fixed left femur.
SELECTED BIOMECHANICAL ISSUES OF LIFTING: A FOCUS ON REDUCING BACK INJURY
Lifting heavy objects can generate large compression, tension, and shear forces throughout the body, most notably across the lumbopelvic regions. At some critical level, forces acting on a region may exceed the structural tolerance of the local muscles, ligaments and capsules, and apophyseal and interbody joints. Lifting is a leading risk factor associated with low-back pain in the United States and is especially related to occupation.50,52,74,85 Disability associated with low-back pain is a significant problem, in terms of both cost and suffering. An estimated 30% of the workforce in the United States regularly handles materials in a potentially harmful manner, including by lifting.112
Muscular Mechanics of Extension of the Low Back during Lifting
ESTIMATING THE MAGNITUDE OF FORCE IMPOSED ON THE LOW BACK DURING LIFTING
Considerable research has been undertaken to quantify the relative demands placed on the various structures in the low back during lifting or performance of other strenuous activities.3,11,30,125,153 This research helps clinicians and members of governmental agencies develop safety guidelines and limits for lifting, especially in the workplace.* Of particular interest with regard to lifting injury are the variables of peak force (or torque) produced by muscles; tension developed within stretched ligaments; and compression and shear forces developed against the intervertebral discs and apophyseal joints. Measurement of these variables is typically not made directly but estimated through sophisticated mathematic or computer-based models. A simple but less accurate method of estimating forces imposed on the low back uses calculations based on the assumption of static equilibrium.
Figure 10-33 (top box) shows the data required to make an approximate estimate of the compression force against the L2 vertebra during lifting. The subject is depicted midway through a vertical lift of a moderately heavy load, weighing 25% of his body weight. The axis of rotation for the sagittal plane motion is oriented in the medial-lateral direction, arbitrary set at L2 (see Figure 10-33, open circle). Estimating the compression force is a two-step process; each step assumes a condition of static rotary and linear equilibrium.
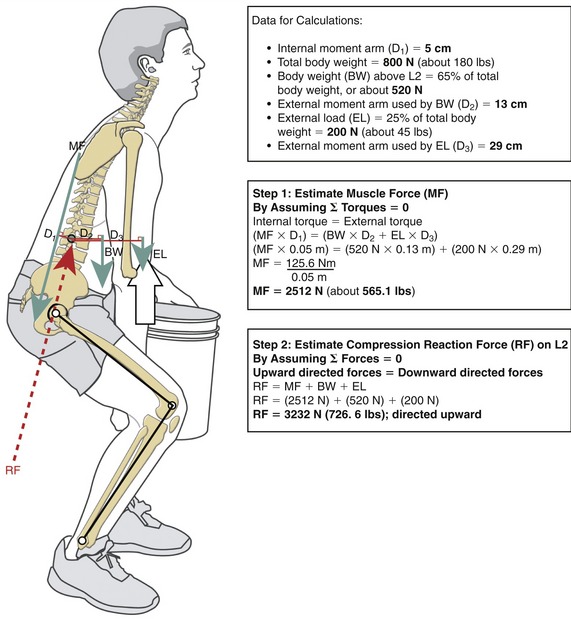
Step 2 estimates the compressive reaction force (RF) imposed on the L2 vertebra during lifting. (This reaction force implies that the L2 vertebra must “push” back against the other downward acting forces.) A rough estimate of this force can be made by assuming static linear equilibrium. (For the sake of simplicity, the calculations assume that muscle force [MF] acts totally in the vertical direction and is therefore parallel with body weight and the external load forces.) The RF vector (see Figure 10-33) is also assumed to be equal in magnitude but opposite in direction to the sum of MF, BW, and EL.
The solution to this example suggests that a compression force of approximately 3232 N (over 725 lb) is exerted on L2 while an external load weighing 200 N (about 45 lb) is lifted. To put this magnitude of force into practical perspective, consider the following two points. First, the National Institute of Occupational Safety and Health (NIOSH) has set guidelines to protect workers from excessive loads on the lumbar region caused by lifting and handling materials. NIOSH has recommended an upper safe limit of 3400 N (764 lb) of compression force on the L5-S1 junction.1,151 Second, the maximal load-carrying capacity of the lumbar spine is estimated to be 6400 N (1439 lb),76 almost twice the maximal safe force recommended by NIOSH. The limit of 6400 N of force applies to a 40-year-old man; this limit decreases by 1000 N each subsequent decade. These force values are very general guidelines that may not always apply to all persons in all lifting situations.
WAYS TO REDUCE THE FORCE DEMANDS ON THE BACK MUSCLES DURING LIFTING
The calculations performed in Step 2 of Figure 10-33 show that muscle force (MF) is, by far, the most influential variable for determining the magnitude of the compressive (reaction) force on the lumbar spine. Proportional reductions in muscle force, therefore, have the greatest effect on reducing the overall compression force on the structures in the low back.
An important factor responsible for the large forces in the low-back muscles during lifting is the disparity in the length of the associated internal and external moment arms. The internal moment arm (D1) depicted in Figure 10-33 is assumed to be 5 cm. The extensor muscles are therefore at a sizable mechanical disadvantage and must produce a force many times larger than the weight of the load being lifted. As previously demonstrated, lifting an external load weighing 25% of one’s body weight produces a compression force on L2 of four times one’s body weight!
Third, reduce the length of the external moment arm of the external load. This is likely the most effective and practical method of decreasing compressive reaction forces on the low back.23 As demonstrated in Figure 10-33, ideally a load should be lifted from between the knees, thereby minimizing the distance between the load and the lumbar region. Based on the calculations, this ideal method of lifting produced a compression force on the lumbar region that remained close to the upper limits of safety proposed by NIOSH. Lifting the same load with a longer external moment arm may create very large and potentially dangerous compression forces on the low back. Figure 10-34 shows a plot of predicted compression (reaction) forces on the L5-S1 disc as a function of both load size and distance between the load and the front of the chest.31 Although perhaps an extreme and unrealistic example, the plot predicts that holding an external load weighing 200 N (45 lb) 50 cm in front of the body creates about 4500 N of compression force, greatly exceeding the upper safe limit of 3400 N.
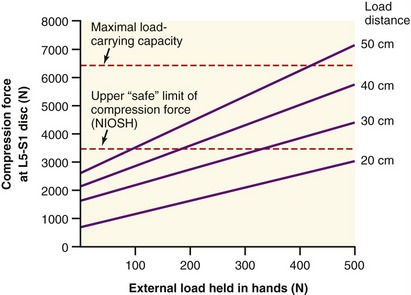
Fourth, increase the internal moment arm available to the low-back extensor muscles. A larger internal moment arm for extension allows a given extension torque to be generated with less muscle force. As stated, less muscle force typically equates to less force on the vertebral elements. Increased lumbar lordosis does indeed increase the internal moment arm available to the lumbar erector spinae muscles.10,140 Lifting with an accentuated lumbar lordosis, however, is not always possible or desirable. Lifting a very heavy load off the floor, for example, typically requires a flexed lumbar spine, which decreases the extensor muscles’ moment arm.81 (Biomechanically, this situation would require greater muscle force per given extensor torque.) Even if possible, maintaining an exaggerated lumbar lordosis may have the negative consequences of generating excessive compression loads on the apophyseal joints and other posterior elements of the spine.
ROLE OF INCREASING INTRA-ABDOMINAL PRESSURE DURING LIFTING
Bartelink first introduced the notion that the Valsalva maneuver, typically used while large loads are lifted, may help unload and thereby protect the lumbar spine.17 The Valsalva maneuver describes the action of voluntarily increasing intra-abdominal pressure by vigorous contraction of the abdominal muscles against a closed glottis. The Valsalva maneuver creates a rigid column of high pressure within the abdomen that pushes upward against the diaphragm, anteriorly against the deeper abdominal muscles (transversus abdominis and internal oblique), posteriorly against the lumbar spine, and downward against the pelvic floor muscles. With this column acting as an inflated “intra-abdominal balloon,” Bartelink proposed that performing the Valsalva maneuver while lifting would create an extension torque on the lumbar spine, thereby reducing the demands on the lumbar extensor muscles and ultimately lowering the muscular-based compression forces on the lumbar spine.
Although the notion of strongly increasing intra-abdominal pressure as a way to reduce compression forces on the spine is intriguing, studies have generally refuted the overall biomechanical validity of the concept.10,13,105,111 Although evidence exists that the Valsalva maneuver does indeed generate a modest lumbar extension torque,67 the strong activation of the abdominal muscles actually creates a net increase in compression forces on the lumbar spine. Because all abdominal muscles (except the transversus abdominis) are strong flexors of the trunk and lumbar spine, their strong activation requires even greater counterbalancing forces from the antagonistic extensor muscles. The resulting increased activation of virtually all the trunk muscles creates an overall increase in muscle-based compression forces on the lumbar spine.11
Most persons, however, likely benefit from the Valsalva maneuver while lifting. In a healthy person without low back pathology, the resulting increased compression force on the lumbar spine can be a useful and relatively safe source of stability to the region. A strong contraction of the abdominal muscles also provides an important bracing effect to the lumbopelvic region, which is helpful in resisting unwanted torsions created by the asymmetric lifting of external loads.38,55 Forces produced by the transversus abdominis may be particularly effective in stabilizing the lumbopelvic region during lifting, for at least two reasons. First, the transversus abdominis has extensive attachments into the thoracolumbar fascia. Forces produced by muscle activation generate a circumferential corset effect around the entire low-back region. Second, by acting primarily in the transverse direction, the transversus abdominis can increase intra-abdominal pressure without creating a concurrent flexion torque or an increase in vertical compression force on the lumbar spine.11,90 The transverse fibers of the internal oblique muscles are able to assist the transversus abdominis with these aforementioned functions.
ADDITIONAL SOURCES OF EXTENSION TORQUE USED FOR LIFTING
The maximal force-generating capacity of the low-back extensor muscles in a typical young adult is estimated to be approximately 4000 N (900 lb).23 If an average internal moment arm of 5 cm is assumed, this muscle group is then expected to produce about 200 Nm of trunk extension torque (i.e., 4000 N × 0.05 m). What is perplexing, however, is the fact that maximal-effort lifting likely requires extensor torques that may greatly exceed 200 Nm. For instance, the person depicted lifting the load in Figure 10-33 would have exceeded his theoretic 200-Nm strength limit if the external load were increased to about 80% of his body weight. Although this is a considerable weight, it is not unusual for a person to successfully lift much greater loads, such as those regularly encountered by heavy labor workers and by competitive “power lifters.” In attempts to explain this apparent discrepancy, two secondary sources of extension torque are proposed: (1) passive tension generated from stretching the posterior ligamentous system, and (2) muscular-generated tension transferred through the thoracolumbar fascia.
Passive Tension Generation from Stretching the Posterior Ligamentous System: When stretched, healthy ligaments and fascia exhibit some degree of natural elasticity. This feature allows connective tissue to temporarily store a small part of the force that initially causes the elongation. Bending forward in preparation for lifting progressively elongates several connective tissues in the lumbar region, and presumably the passive tension developed in these tissues can assist with an extension torque.10,45 These connective tissues, collectively known as the posterior ligamentous system, include the posterior longitudinal ligament, ligamentum flavum, apophyseal joint capsule, interspinous ligament, and posterior layer of the thoracolumbar fascia.53
In theory, about 72 Nm of total passive extensor torque are produced by maximally stretching the posterior ligamentous system (Table 10-6).23 Adding this passive torque to the hypothetic 200 Nm of active torque yields a total of 272 Nm of extension torque available for lifting. A fully engaged (stretched) posterior ligamentous system can therefore generate about 25% of the total extension torque for lifting. Note, however, that this 25% passive torque reserve is available only after the lumbar spine is maximally flexed, which in reality is rare during lifting. Even some competitive power lifters, who appear to lift with a fully rounded low back, avoid the extremes of flexion.34 It is generally believed that maximum or near-maximum flexion of the lumbar spine should be avoided during lifting.23,104 The lumbar region should be held in a near-neutral position.104 This position favors a near-maximal contact area within the apophyseal joints, which may help reduce articular stress. Furthermore, maintaining the neutral position during lifting may align the local extensor muscles to be most effective at resisting anterior shear.102
TABLE 10-6.
Maximal Passive Extensor Torque Produced by Stretched Connective Tissues in the Lumbar Region
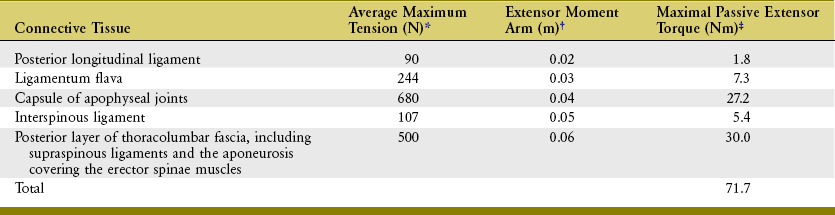
*Average maximum tension is the tension within each stretched tissue at the point of rupture.
†Extensor moment arm is the perpendicular distance between the attachment sites of the ligaments and the medial-lateral axis of rotation within a representative lumbar vertebra.
‡Maximal passive extensor torque is estimated by the product of maximum tension (force) and extensor muscle moment arm.
Data from Bogduk N, Twomey L: Clinical anatomy of the lumbar spine, ed 4, New York, 2005, Churchill Livingstone.
Although the neutral position of the lumbar spine while lifting may reduce the chance of injury to the low back, it engages only a small portion of the total passive torque reserve available to assist with extension. Most of the extension torque must therefore be generated by active muscle contraction.121 It is important, therefore, that the extensor muscles be strong enough to meet the potentially large demands placed on the low back by heavy lifting. Adequate strength in the lumbar multifidi is particularly critical in this regard.21,47,57 Without adequate strength in these muscles, the lumbar spine may be pulled into excessive flexion by the external torque imposed by the large load. Excessive flexion in the lumbar region while loads are lifted is generally not considered a safe lifting technique.
Muscular-Generated Tension Transferred through the Thoracolumbar Fascia: The thoracolumbar fascia is thickest and most extensively developed in the lumbar region (see Figure 9-72). Much of the tissue attaches to the lumbar spine, sacrum, and pelvis in a position well posterior to the axis of rotation at the lumbar region. Theoretically, therefore, passive tension within stretched thoracolumbar fascia can produce an extension torque in the lumbar region and thus augment the torque created by the low-back musculature.
In order for the thoracolumbar fascia to generate useful tension, it must be first stretched and rendered taut. This can occur in two ways. First, the fascia is stretched simply when one bends forward and flexes the lumbar spine in preparation for lifting. Second, the fascia is stretched by active contraction of muscles that attach directly into the thoracolumbar fascia. The prevailing horizontal fiber direction of most of the thoracolumbar fascia, however, limits the amount of extension torque that can be produced at the lumbar spine.24 Theoretically, the force transferred to the thoracolumbar fascia by contraction of the transversus abdominis and internal oblique may contribute 6 Nm of extensor torque across the lumbar spine (compared with the approximately 200 Nm of active torque generated by the low-back extensor muscles99). Although the actual extension torque is small, the tension transferred through the thoracolumbar fascia provides an additional element of stabilization to the region.
The latissimus dorsi and gluteus maximus may also indirectly contribute to lumbar extension torque via their extensive attachments to the thoracolumbar fascia. Both are active during lifting, but for different reasons (Figure 10-35). The gluteus maximus stabilizes and controls the hips. The latissimus dorsi helps transfer the external load being lifted from the arms to the trunk. In addition to attaching into the thoracolumbar fascia, the latissimus dorsi attaches into the posterior aspect of the pelvis, sacrum, and spine. Based on these attachments and its relative moment arm for producing lumbar extension (see Figure 10-16), the latissimus dorsi has all the attributes of a respected extensor of the low back. The oblique fiber direction of the muscle as it ascends the trunk can also provide torsional stability to the axial skeleton, especially when bilaterally active. This stability may be especially useful when large loads are handled in an asymmetric fashion.
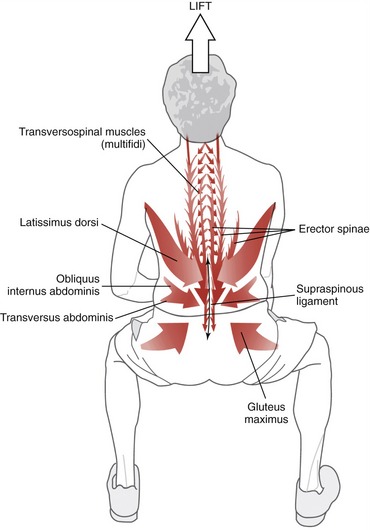
SPECIAL FOCUS 10-7  Two Contrasting Lifting Techniques: the Stoop versus the Squat Lift
Two Contrasting Lifting Techniques: the Stoop versus the Squat Lift
The stoop lift and the squat lift represent the biomechanical extremes of a broad continuum of possible lifting strategies (Figure 10-36). Understanding some of the biomechanic and physiologic differences between these methods of lifting may provide insight into the advantages or disadvantages of other, more common lifting strategies.
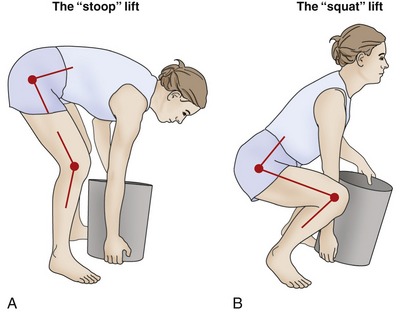
The stoop lift is performed primarily by extending the hips and lumbar region while the knees remain slightly flexed (see Figure 10-36, A). This lifting strategy is associated with greater flexion of the low back, especially at the initiation of the lift. By necessity, the stoop lift creates a long external moment arm between the trunk (and load) and the low back. The greater external torque requires greater extension forces from the low-back and trunk extensor muscles. In combination with a markedly flexed lumbar spine, the stoop lift can create large and possibly damaging compression and shear forces on the discs.
The squat lift, in contrast, typically begins with near maximally flexed knees (see Figure 10-36, B). The knees and hips extend during the lift, powered by the quadriceps and hip extensor muscles. Depending on the physical characteristics of the load and the initial depth of the squat, the lumbar region may remain extended, in a neutral position, or partially flexed throughout the lift. Perhaps the greatest advantage of the squat lift is that it typically allows the load to be raised more naturally from between the knees. The squat lift can, in theory, reduce the external moment arm of the load and trunk and, as a consequence, diminish the extensor torque demands on the muscles of the back.
The squat lift is most often advocated as the safer of the two techniques in terms of producing less stress on the low back and therefore preventing back injuries.18 Little overwhelming direct proof, however, can be found to support this strongly held clinical belief.10,23,143 As with many espoused clinical principles, the advantage of one particular concept or technique is often at least partially offset by a disadvantage. This holds true for the apparent advantage of the squat lift over the stoop lift. Although the squat lift may reduce the demands on the extensor muscles and other tissues in the low back, it usually creates greater demands on the knees.129 The extreme degree of initial knee flexion associated with the full squat places high force demands on the quadriceps muscles to extend the knees. The forces impose very large pressures across the tibiofemoral and patellofemoral joints. Healthy persons may tolerate high pressures at these joints without negative consequences; however, someone with painful or arthritic knees may not. The adage that lifting with the legs “spares the back and spoils the knees” does, therefore, have some validity.
Another factor to consider when comparing the benefits of the squat lift over the stoop lift is the total work required to lift the load. The mechanical work performed during lifting is equal to the weight of the body and the load multiplied by the vertical displacement of the body and the load. The stoop lift is 23% to 34% more metabolically “efficient” than the squat lift in terms of work performed per level of oxygen consumption.152 The squat lift requires greater work because a greater proportion of the total body mass must be moved through space.
Rather than performing a squat lift or a stoop lift, in reality most people choose an individualized or freestyle lifting technique. A freestyle technique allows the lifter to combine some of the benefits of the squat lift with the more metabolically efficient stoop lift. Workers have reported a higher self-perceived maximal safe limit when allowed to lift with a freestyle technique rather than with a set technique.134
Summary of Factors That Likely Contribute to Safe Lifting
The lifting technique used in Figure 10-33 illustrates two fundamental features that likely contribute to safe lifting technique: (1) the lumbar spine is held in a neutral lordotic position, and (2) the load is lifted from between the knees. The rationales for these and other factors considered to contribute to safe lifting are listed in Table 10-7. Other, more general considerations include (1) knowing one’s physical limits, (2) thinking the lift through before the event, and (3) within practical and health limits, remaining in optimal physical and cardiovascular condition.
Additional Clinical Connections
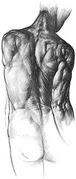
CLINICAL CONNECTION 10-1 A Closer Look at the Spinal Stabilizing Functions of Selected Abdominal Muscles and the Lumbar Multifidi
The following discussion highlights examples of spinal stabilization functions performed by the abdominal muscles (most notably the transversus abdominis and internal oblique) and the multifidi. These muscles are featured primarily because of the large body of research that has focused on their ability (or lack thereof) to stabilize the lumbopelvic region of the trunk (which includes the lumbar spine, lumbosacral junction, and sacroiliac joints).4,142 The topic of lumbopelvic muscular stabilization has attracted the attention of researchers and clinicians primarily because of the high incidence of instability and stress-related degeneration in this region.
Much of what is known about the kinesiology of the muscular stabilizers of the lumbopelvic region is based on electromyographic (EMG) research, often with the use of fine-wire (needle) electrodes. One common methodology used in this research involves the recording of the order in which various trunk muscles respond to expected or unexpected whole-body perturbations. As an example, Figure 10-37, A shows the onset of the EMG responses of a selected set of abdominal muscles as a healthy, pain-free person rapidly flexes his arm after a visual stimulus.142 The top EMG signal (depicted in red) is from a shoulder flexor—the anterior deltoid—and the remaining EMG signals are from the external oblique, the middle and lower regions of the internal oblique, and the upper, middle, and lower regions of the transversus abdominis. All muscles recorded from this one subject responded at slightly different times (indicated by vertical arrows) relative to the initiation of the deltoid’s EMG signal (red dashed line). Figure 10-37, B shows the overall data from the experiment, based on 11 healthy subjects.72
As previously discovered through research in this area, the lower and middle fibers of the transversus abdominis and internal oblique muscles consistently activate before the activation of the deltoid muscle.141 This anticipatory muscle response is believed to be a subconscious, feedforward mechanism employed by the nervous system to minimize reactive countermovements of the trunk.142 Although subtle and not completely understood, this anticipatory muscular response may help protect the lumbopelvic region from potentially damaging shear forces.5
It is interesting that multiple regions of the transversus abdominis and internal oblique activate at different times in response to the rapidly elevated arm. It is as though the different regions within these muscles respond as distinct anatomic entities. Although separated by only a very short time period, the sequential muscular responses provide insight into the complex stabilizing functions of these muscles. Consider, in this regard, the following proposed functions for each of the three regions of the transversus abdominis.124,131,142 Contraction of the upper fibers of the transversus abdominis may help stabilize the rib cage and linea alba. The lower fibers are believed to compress and thereby help stabilize the sacroiliac joints.68 Contraction of the middle fibers of the transversus abdominis transfers tension directly to the lumbar spinous processes and sacrum by connections into the thoracolumbar fascia (see Chapter 9). This action is part of the “corset” effect described earlier in this chapter for this muscle.
Furthermore, bilateral contraction of the middle fibers of the transversus abdominis is particularly effective (along with other abdominal muscles) at compressing the abdominal cavity and thereby increasing intra-abdominal pressure (described earlier as the Valsalva maneuver). Evidence exists that the rise in intra-abdominal pressure not only exerts a modest extension torque on the lumbar spine, but also stabilizes the region.67,68 For the most effective stabilization, the cylinder-like abdominal cavity must also be simultaneously compressed from both its cranial and caudal ends. This is normally accomplished by concurrent activation and descent of the diaphragm muscle—the roof of the abdominal cavity—and activation and ascent of the pelvic floor muscles—the ultimate floor of the abdominal cavity. Although sparse, evidence does exist from both animal and human subjects that these muscular interactions do indeed occur in a coordinated manner, with a resulting increased stiffness in the lumbar spine.70
The experimental methodology illustrated in Figure 10-37 has also been used to study the sequential activation of the abdominal muscles in response to rapidly flexing the lower limb.72 Consistently, the abdominal muscles (including the rectus abdominis) respond before the activation of the hip flexor muscles. It is interesting that the transversus abdominis and internal oblique are consistently the first of the trunk muscles to respond, on average 50 to 100 msec before the hip flexor muscles. This activation pattern of the abdominal muscles, as a group, reflects their need to stabilize the lower trunk during the leg movement, as well as to fixate the lumbopelvic region against the pull of the contracting hip flexor muscles. The transversus abdominis and the oblique abdominal muscles also respond before rapid active hip abduction and extension movements as well. These abdominal muscles appear “dedicated” to stabilizing the lower trunk, regardless of the direction of the forces produced by the contracting hip musculature.
Hodges and colleagues used a similar experimental protocol to study the sequential muscle activation in persons with chronic low back pain.71,110,123 Remarkably, this research showed a consistent, short delay in the onset of EMG signals from the transversus abdominis—the activation of this muscle occurring most often after the activation of the prime movers of the rapid limb motion. Whether a short delay in abdominal muscle activation can create sufficient reactive stress in the lumbopelvic region to ultimately cause low-back pain is not known, although it is an intriguing question. Cadaveric studies have indeed shown that axial rotation of as little as 2 to 3 degrees per intervertebral lumbar junction can potentially injure the apophyseal and interbody joints (see Chapter 9). A single “unprotected” stress event may not be significant; however, multiple events that accumulate over many years may predispose the region to injury.
Research has shown that in addition to the transversus abdominis and internal oblique muscles, the lumbar multifidi are consistently recruited early in healthy persons in response to various perturbations imposed against the body.27,57,88,89,154 The multifidi are extremely capable stabilizers of the lumbar spine, especially in the lower segments.21,149 The muscles’ regional extensor strength is augmented by their relatively large size; they account for about one third of the total cross-sectional area of all deep paraspinal muscles at the L4 level.23,98 In addition to their thickness, the multifidi also have a highly segmented morphology and innervation and are rich with muscle spindles.103,123,154 These anatomic features favor precise and, when needed, authoritative control over intersegmental lumbar stability.*
The lumbar multifidi have consistently shown preferential and persistent atrophy in persons with acute or chronic low-back pain.63,96 This finding is noteworthy, considering the muscles’ proposed importance in stabilizing the lumbar region. The amount of atrophy in the lumbar multifidi is striking; a 30% reduction in cross-sectional area has been reported,64 in some cases within days of the onset of the painful symptoms.66 The reason for preferential atrophy in these muscles is uncertain. Evidence suggests several explanations, however, ranging from denervation after nerve root injury to reflexive neural inhibition after trauma to the disc or capsule of the apophyseal joints.21
Marked and persisting atrophy of the lumbar multifidi has also been demonstrated in pain-free healthy subjects who were subjected to 8 weeks of strict bed rest.16,123 Of particular interest was the response of a subgroup of the subjects who, while remaining on strict bed rest, were allowed to exercise twice daily (performing resistive exercise in conjunction with receiving whole-body vibration). These subjects demonstrated statistically less multifidi atrophy, and the atrophy did not persist as long as in the inactive, control group of subjects. It appears that the lumbar multifidi are particularly sensitive to musculoskeletal pathology in the lumbar region, as well as reduced weight bearing through the axial skeleton. Regardless of the underlying mechanism, it is reasonable to assume that marked and prolonged atrophy of these muscles reduces the mechanical stability of the lumbar spine, potentially leaving it vulnerable to stress-related injury. For this reason, exercises designed for the treatment of persons with low-back pain often incorporate specific exercises to strengthen the lumbar multifidi.14,47,63
CLINICAL CONNECTION 10-3 Torticollis and Sleeping Position: Is There a Link?
Torticollis (from the Latin tortus, twisted, + collum, neck) or “wryneck” typically describes a pathologic condition of chronic shortening of the sternocleidomastoid muscle. The condition, generally identified in the young child or infant, may be congenital or acquired. A child or infant with torticollis typically has an asymmetric craniocervical posture that reflects the primary actions of the tightened muscle. The child illustrated in Figure 10-38 has a tight left sternocleidomastoid (see arrow), with a corresponding posture of slight left lateral flexion combined with right axial rotation of his craniocervical region.
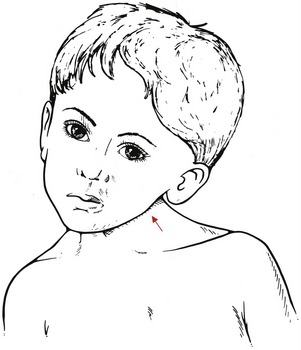
The incidence of torticollis is 0.4% to 3.9% of newborn infants in the United States.32,43 The range of these estimates reflects different methods for detecting the condition. The underlying cause of torticollis most often involves muscle tissue, although it may involve nonmuscular systems. The far more common muscular-based torticollis usually involves fibrous growths within the sternocleidomastoid—a condition termed idiopathic muscular fibrosis.43 Although the exact cause of this condition is unknown, it is frequently associated with a difficult childbirth labor, breech delivery, or intrauterine malpositioning or crowding.32 The more serious, non–muscular-based torticollis involves pathology associated with the nervous system (including vision) or the skeletal system (typically associated with cervical dysplasia).43
Approximately one third of infants with torticollis also develop plagiocephaly.43 This condition is an abnormal molding and subsequent distortion in the shape of a young infant’s naturally soft cranium. The distorted shape is typically caused by the infant’s head resting in a single prolonged position against another surface. Some authors believe that an infant with an existing torticollis may develop a secondary plagiocephaly (involving the posterior-lateral cranium) before or shortly after birth, as a result of the prolonged and concentrated contact against the infant’s rotated cranium. Alternately, other authors assert that an infant born free of torticollis may eventually develop plagiocephaly with a secondary torticollis simply because of a favored rotated position of the head while the infant sleeps in a supine position.2 Once developed, the positional plagiocephaly strongly reinforces the established asymmetric (rotated) head position adapted for sleeping. The constant rotated position of the head produces a chronic slackening of the contralateral sternocleidomastoid, which eventually develops into a contracture and the classic expression of torticollis. According to deChalain, many infants who develop torticollis after plagiocephaly do not have fibrotic changes in the tightened sternocleidomastoid muscle; the deformity develops purely as a consequence of muscle tightness caused by the abnormal craniocervical positioning.43,155
The notion that plagiocephaly can, in some cases, lead to a positional torticollis was reinforced by a series of events that occurred in the 1990s. Within this decade, the American Academy of Pediatrics published recommendations that healthy infants be placed in a supine position for sleeping as a way to reduce the incidence of sudden infant death syndrome (SIDS).155 The so-called “back-to-sleep” recommendation had a dramatic effect on the sleeping pattern of many infants in the United States. The incidence of infants positioned prone for sleep decreased by 66% from 1992 to 1996.9,139 Although a direct cause-and-effect relationship cannot be unequivocally stated, the rate of SIDS declined approximately 38% during this same time period.43 The remarkable and simultaneous decline in the incidence of SIDS nevertheless strongly reinforced the fundamental premise of the “back-to-sleep” campaign. Subsequent evidence strongly suggests that the increased frequency of supine-only sleeping has also led to an increase in the incidence of positional plagiocephaly, most notably affecting the posterior-lateral cranium.79 Furthermore, additional data show that the dramatic increase in positional plagiocephaly has led to a parallel increase in positional torticollis.119
Without a doubt, the huge and life-saving success of the “back-to-sleep” campaign of the 1990s far outweighs the potential negative consequence caused by the increased incidence of plagiocephaly and secondary torticollis. Efforts are ongoing to minimize the incidence of the latter two conditions. Clinicians have advised parents or guardians to alternate the head position of the supine-positioned infant.41,54,77,126 Clinicians also advocate that parents or guardians set aside short periods of supervised and interactive “prone-play” (or “tummy time”) with the infant, while still strictly adhering to the “back-to-sleep” principle.33 Encouraging more prone-lying while infants are awake will very likely reduce the likelihood of developing the plagiocephaly (and secondary torticollis), and may also facilitate the infant’s natural motor development.33
REFERENCES
1. National Institute for Occupational Safety and Health (NIOSH): Work Practices Guide for Manual Lifting, Report No. 81-122, Cincinnati, Ohio, NIOSH 1992.
2. American Academy of Pediatrics Task Force on Infant Positioning and SIDS. Positioning and sudden infant death syndrome (SIDS): update. Pediatrics. 1996;98:1216–1218.
3. Adams, MA, Dolan, P. A technique for quantifying the bending moment acting on the lumbar spine in vivo. J Biomech. 1991;24:117–126.
4. Akuthota, V, Ferreiro, A, Moore, T, Fredericson, M. Core stability exercise principles. Curr Sports Med Rep. 2008;7:39–44.
5. Allison, GT, Morris, SL, Lay, B. Feedforward responses of transversus abdominis are directionally specific and act asymmetrically: implications for core stability theories. J Orthop Sports Phys Ther. 2008;38:228–237.
6. Anderson, JS, Hsu, AW, Vasavada, AN. Morphology, architecture, and biomechanics of human cervical multifidus. Spine. 2005;30:E86–E91.
7. Andersson, EA, Grundstrom, H, Thorstensson, A. Diverging intramuscular activity patterns in back and abdominal muscles during trunk rotation. Spine. 2002;27:E152–E160.
8. Andersson, EA, Nilsson, J, Ma, Z, Thorstensson, A. Abdominal and hip flexor muscle activation during various training exercises. Eur J Appl Physiol Occup Physiol. 1997;75:115–123.
9. Argenta, LC, David, LR, Wilson, JA, Bell, WO. An increase in infant cranial deformity with supine sleeping position. J Craniofac Surg. 1996;7:5–11.
10. Arjmand, N, Shirazi-Adl, A. Biomechanics of changes in lumbar posture in static lifting. Spine. 2005;30:2637–2648.
11. Arjmand, N, Shirazi-Adl, A. Role of intra-abdominal pressure in the unloading and stabilization of the human spine during static lifting tasks. Eur Spine J. 2006;15:1265–1275.
12. Arjmand, N, Shirazi-Adl, A, Parnianpour, M. Trunk biomechanics during maximum isometric axial torque exertions in upright standing. Clin Biomech (Bristol, Avon). 2008;23:969–978.
13. Aspden, RM. Intra-abdominal pressure and its role in spinal mechanics. Clin Biomech (Bristol, Avon). 1987;2:168–174.
14. Ballock, RT, Song, KM. The prevalence of nonmuscular causes of torticollis in children. J Pediatr Orthop. 1996;16:500–504.
15. Barr, KP, Griggs, M, Cadby, T. Lumbar stabilization: core concepts and current literature, Part 1. Am J Phys Med Rehabil. 2005;84:473–480.
16. Barr, KP, Griggs, M, Cadby, T. Lumbar stabilization: a review of core concepts and current literature, Part 2. Am J Phys Med Rehabil. 2007;86:72–80.
17. Bartelink, DL. The role of abdominal pressure in relieving the pressure on the lumbar intervertebral discs. J Bone Joint Surg Br. 1957;39:718–725.
18. Bazrgari, B, Shirazi-Adl, A, Arjmand, N. Analysis of squat and stoop dynamic liftings: muscle forces and internal spinal loads. Eur Spine J. 2007;16:687–699.
19. Beattie, PF. Current understanding of lumbar intervertebral disc degeneration: a review with emphasis upon etiology, pathophysiology, and lumbar magnetic resonance imaging findings. J Orthop Sports Phys Ther. 2008;38:329–340.
20. Beimborn, DS, Morrissey, MC. A review of the literature related to trunk muscle performance. Spine. 1988;13:655–660.
21. Belavý, DL, Hides, JA, Wilson, SJ, et al. Resistive simulated weightbearing exercise with whole body vibration reduces lumbar spine deconditioning in bed-rest. Spine. 2008;33:E121–E131.
22. Bergmark, A. Stability of the lumbar spine. A study in mechanical engineering. Acta Orthop Scand Suppl. 1989;230:1–54.
23. Bogduk, N. Clinical Anatomy of the Lumbar Spine and Sacrum, ed 4. New York: Churchill Livingstone; 2005.
24. Bogduk, N, Macintosh, JE. The applied anatomy of the thoracolumbar fascia. Spine. 1984;9:164–170.
25. Boyd-Clark, LC, Briggs, CA, Galea, MP. Muscle spindle distribution, morphology, and density in longus colli and multifidus muscles of the cervical spine. Spine. 2002;27:694–701.
26. Brenner, AK, Gill, NW, Buscema, CJ, Kiesel, K. Improved activation of lumbar multifidus following spinal manipulation: a case report applying rehabilitative ultrasound imaging. J Orthop Sports Phys Ther. 2007;37:613–619.
27. Briggs, AM, Greig, AM, Bennell, KL, Hodges, PW. Paraspinal muscle control in people with osteoporotic vertebral fracture. Eur Spine J. 2007;16:1137–1144.
28. Brown, SH, Vera-Garcia, FJ, McGill, SM. Effects of abdominal muscle coactivation on the externally preloaded trunk: variations in motor control and its effect on spine stability. Spine. 2006;31:E387–E393.
29. Buford, JA, Yoder, SM, Heiss, DG, Chidley, JV. Actions of the scalene muscles for rotation of the cervical spine in macaque and human. J Orthop Sports Phys Ther. 2002;32:488–496.
30. Butler, HL, Hubley-Kozey, CL, Kozey, JW. Changes in trunk muscle activation and lumbar-pelvic position associated with abdominal hollowing and reach during a simulated manual material handling task. Ergonomics. 2007;50:410–425.
31. Chaffin, DB, Andersson, GBJ. Occupational Biomechanics, 2nd ed. New York: John Wiley and Sons; 1991.
32. Chen, MM, Chang, HC, Hsieh, CF, et al. Predictive model for congenital muscular torticollis: analysis of 1021 infants with sonography. Arch Phys Med Rehabil. 2005;86:2199–2203.
33. Cheng, JC, Wong, MW, Tang, SP, et al. Clinical determinants of the outcome of manual stretching in the treatment of congenital muscular torticollis in infants. A prospective study of eight hundred and twenty-one cases. J Bone Joint Surg Am. 2001;83:679–687.
34. Cholewicki, J, McGill, SM. Lumbar posterior ligament involvement during extremely heavy lifts estimated from fluoroscopic measurements. J Biomech. 1992;25:17–28.
35. Cholewicki, J, Reeves, NP. All abdominal muscles must be considered when evaluating the intra-abdominal pressure contribution to trunk extensor moment and spinal loading. J Biomech. 2004;37:953–954.
36. Cholewicki, J, VanVliet, JJ. Relative contribution of trunk muscles to the stability of the lumbar spine during isometric exertions. Clin Biomech (Bristol, Avon). 2002;17:99–105.
37. Cook, TM, Neumann, DA. The effects of load placement on the EMG activity of the low back muscles during load carrying by men and women. Ergonomics. 1987;30:1413–1423.
38. Cresswell, AG, Grundstrom, H, Thorstensson, A. Observations on intra-abdominal pressure and patterns of abdominal intra-muscular activity in man. Acta Physiol Scand. 1992;144:409–418.
39. Cutter, N. Handbook of Manual Muscle Testing. St Louis: McGraw-Hill; 1999.
40. Danneels, LA, Vanderstraeten, GG, Cambier, DC, et al. CT imaging of trunk muscles in chronic low back pain patients and healthy control subjects. Eur Spine J. 2000;9:266–272.
41. Davis, BE, Moon, RY, Sachs, HC, Ottolini, MC. Effects of sleep position on infant motor development. Pediatrics. 1998;102:1135–1140.
42. Dawson, AP, McLennan, SN, Schiller, SD, et al. Interventions to prevent back pain and back injury in nurses: a systematic review. Occup Environ Med. 2007;64:642–650.
43. de Chalain, TM, Park, S. Torticollis associated with positional plagiocephaly: a growing epidemic. J Craniofac Surg. 2005;16:411–418.
44. Deng, YC, Goldsmith, W. Response of a human head/neck/upper-torso replica to dynamic loading—I. Physical model. J Biomech. 1987;20:471–486.
45. Dolan, P, Mannion, AF, Adams, MA. Passive tissues help the back muscles to generate extensor moments during lifting. J Biomech. 1994;27:1077–1085.
46. Ebenbichler, GR, Oddsson, LI, Kollmitzer, J, Erim, Z. Sensory-motor control of the lower back: implications for rehabilitation. Med Sci Sport Exerc. 2001;33:1889–1898.
47. Ekstrom, RA, Osborn, RW, Hauer, PL. Surface electromyographic analysis of the low back muscles during rehabilitation exercises. J Orthop Sports Phys Ther. 2008;38:736–745.
48. Escamilla, RF, Babb, E, DeWitt, R, et al. Electromyographic analysis of traditional and nontraditional abdominal exercises: implications for rehabilitation and training. Phys Ther. 2006;86:656–671.
49. Falla, D, Jull, G, Russell, T, et al. Effect of neck exercise on sitting posture in patients with chronic neck pain. Phys Ther. 2007;87:408–417.
50. Ferguson, SA, Marras, WS. A literature review of low back disorder surveillance measures and risk factors. Clin Biomech (Bristol, Avon). 1997;12:211–226.
51. Ferreira, PH, Ferreira, ML, Maher, CG, et al. Specific stabilisation exercise for spinal and pelvic pain: a systematic review. Aust J Physiother. 2006;52:79–88.
52. Fujishiro, K, Weaver, JL, Heaney, CA, et al. The effect of ergonomic interventions in healthcare facilities on musculoskeletal disorders. Am J Ind Med. 2005;48:338–347.
53. Gracovetsky, S, Farfan, HF, Lamy, C. The mechanism of the lumbar spine. Spine. 1981;6:249–262.
54. Graham, JM, Jr. Tummy time is important. Clin Pediatr. 2006;45:119–121.
55. Grenier, SG, McGill, SM. Quantification of lumbar stability by using 2 different abdominal activation strategies. Arch Phys Med Rehabil. 2007;88:54–62.
56. Halpern, AA, Bleck, EE. Sit-up exercises: an electromyographic study. Clin Orthop Relat Res. 1979;145:172–178.
57. Hansen, L, de, ZM, Rasmussen, J, et al. Anatomy and biomechanics of the back muscles in the lumbar spine with reference to biomechanical modeling. Spine. 2006;31:1888–1899.
58. Hayden JA, van Tulder MW, Malmivaara A, Koes BW: Exercise therapy for treatment of non-specific low back pain, Cochrane Database Syst Rev 20;(3):CD000335, 2005 Jul.
59. Henry, SM, Teyhen, DS. Ultrasound imaging as a feedback tool in the rehabilitation of trunk muscle dysfunction for people with low back pain. J Orthop Sports Phys Ther. 2007;37:627–634.
60. Herbert, WJ, Heiss, DG, Basso, DM. Influence of feedback schedule in motor performance and learning of a lumbar multifidus muscle task using rehabilitative ultrasound imaging: a randomized clinical trial. Phys Ther. 2008;88:261–269.
61. Hides, J, Wilson, S, Stanton, W, et al. An MRI investigation into the function of the transversus abdominis muscle during “drawing-in” of the abdominal wall. Spine. 2006;31:E175–E178.
62. Hides, JA, Richardson, CA, Jull, GA. Multifidus muscle recovery is not automatic after resolution of acute, first-episode low back pain. Spine. 1996;21:2763–2769.
63. Hides, JA, Stanton, WR, McMahon, S, et al. Effect of stabilization training on multifidus muscle cross-sectional area among young elite cricketers with low back pain. J Orthop Sports Phys Ther. 2008;38:101–108.
64. Hides, JA, Stokes, MJ, Saide, M, et al. Evidence of lumbar multifidus muscle wasting ipsilateral to symptoms in patients with acute/subacute low back pain. Spine. 1994;19:165–172.
65. Hislop, HJ, Montgomery, J. Daniel’s and Worthingham’s Muscle Testing, 6th ed. Philadelphia: Saunders; 1995.
66. Hodges, P, Holm, AK, Hansson, T, Holm, S. Rapid atrophy of the lumbar multifidus follows experimental disc or nerve root injury. Spine. 2006;31:2926–2933.
67. Hodges, P, Kaigle, HA, Holm, S, et al. Intervertebral stiffness of the spine is increased by evoked contraction of transversus abdominis and the diaphragm: in vivo porcine studies. Spine. 2003;28:2594–2601.
68. Hodges, PW, Cresswell, AG, Daggfeldt, K, Thorstensson, A. In vivo measurement of the effect of intra-abdominal pressure on the human spine. J Biomech. 2001;34:347–353.
69. Hodges, PW, Cresswell, AG, Thorstensson, A. Perturbed upper limb movements cause short-latency postural responses in trunk muscles. Exp Brain Res. 2001;138:243–250.
70. Hodges, PW, Richardson, CA. Contraction of the abdominal muscles associated with movement of the lower limb. Phys Ther. 1997;77:132–142.
71. Hodges, PW, Richardson, CA. Feedforward contraction of transversus abdominis is not influenced by the direction of arm movement. Exp Brain Res. 1997;114:362–370.
72. Hodges, PW, Richardson, CA. Inefficient muscular stabilization of the lumbar spine associated with low back pain. A motor control evaluation of transversus abdominis. Spine. 1996;21:2640–2650.
73. Hoek van Dijke, GA, Snijders, CJ, Stoeckart, R, Stam, HJ. A biomechanical model on muscle forces in the transfer of spinal load to the pelvis and legs. J Biomech. 1999;32:927–933.
74. Hoogendoorn, WE, Bongers, PM, de Vet, HC, et al. Flexion and rotation of the trunk and lifting at work are risk factors for low back pain: results of a prospective cohort study. Spine. 2000;25:3087–3092.
75. Hyun, JK, Lee, JY, Lee, SJ, Jeon, JY. Asymmetric atrophy of multifidus muscle in patients with unilateral lumbosacral radiculopathy. Spine. 2007;32:E598–E602.
76. Jager, M, Luttmann, A. The load on the lumbar spine during asymmetrical bi-manual materials handling. Ergonomics. 1992;35:783–805.
77. Jantz, JW, Blosser, CD, Fruechting, LA. A motor milestone change noted with a change in sleep position. Arch Pediatr Adolesc Med. 1997;151:565–568.
78. Johnston, V, Jull, G, Souvlis, T, Jimmieson, NL. Neck movement and muscle activity characteristics in female office workers with neck pain. Spine. 2008;33:555–563.
79. Jones, MW. The other side of “back to sleep.”. Neonatal Netw. 2003;22:49–53.
80. Jordan, A, Mehlsen, J, Bülow, PM, et al. Maximal isometric strength of the cervical musculature in 100 healthy volunteers. Spine. 1999;24:1343–1348.
81. Jorgensen, MJ, Marras, WS, Gupta, P, Waters, TR. Effect of torso flexion on the lumbar torso extensor muscle sagittal plane moment arms. Spine. 2003;3:363–369.
82. Juker, D, McGill, S, Kropf, P, Steffen, T. Quantitative intramuscular myoelectric activity of lumbar portions of psoas and the abdominal wall during a wide variety of tasks. Med Sci Sports Exerc. 1998;30:301–310.
83. Kapandji, IA. The physiology of the joints, 5 ed. Edinburgh: Churchill Livingstone; 1982.
84. Kavcic, N, Grenier, S, McGill, SM. Determining the stabilizing role of individual torso muscles during rehabilitation exercises. Spine. 2004;29:1254–1265.
85. Kelsey, JL. The epidemiology of diseases of the hip: a review of the literature. Int J Epidemiol. 1977;6:269–280.
86. Kendall, FP, McCreary, AK, Provance, PG. Muscles: Testing and function, 4th ed. Baltimore: Williams & Wilkins; 1993.
87. Kettler, A, Hartwig, E, Schultheiss, M, et al. Mechanically simulated muscle forces strongly stabilize intact and injured upper cervical spine specimens. J Biomech. 2002;35:339–346.
88. Kiefer, A, Shirazi-Adl, A, Parnianpour, M. Stability of the human spine in neutral postures. Eur Spine J. 1997;6:45–53.
89. Kiefer, A, Shirazi-Adl, A, Parnianpour, M. Synergy of the human spine in neutral postures. Eur Spine J. 1998;7:471–479.
90. Kingma, I, Faber, GS, Suwarganda, EK, et al. Effect of a stiff lifting belt on spine compression during lifting. Spine. 2006;31:E833–E839.
91. Kjaer, P, Bendix, T, Sorensen, JS, et al. Are MRI-defined fat infiltrations in the multifidus muscles associated with low back pain? BMC Medicine. 2007;5:2.
92. Koumantakis, GA, Watson, PJ, Oldham, JA. Trunk muscle stabilization training plus general exercise versus general exercise only: randomized controlled trial of patients with recurrent low back pain. Phys Ther. 2005;85:209–225.
93. Krakenes, J, Kaale, BR. Magnetic resonance imaging assessment of craniovertebral ligaments and membranes after whiplash trauma. Spine. 2006;31:2820–2826.
94. Kuijer, W, Dijkstra, PU, Brouwer, S, et al. Safe lifting in patients with chronic low back pain: comparing FCE lifting task and NIOSH lifting guideline. J Occup Rehabil. 2006;16:579–589.
95. Kumar, S. Moment arms of spinal musculature determined from CT scans. Clin Biomech (Bristol, Avon). 1988;3:137–144.
96. Laasonen, EM. Atrophy of sacrospinal muscle groups in patients with chronic, diffusely radiating lumbar back pain. Neuroradiology. 1984;26:9–13.
97. Liddle, SD, Baxter, GD, Gracey, JH. Exercise and chronic low back pain: what works? Pain. 2004;107:176–190.
98. Macintosh, JE, Bogduk, N. The biomechanics of the lumbar multifidus. Clin Biomech (Bristol, Avon). 1986;1:205–213.
99. Macintosh, JE, Bogduk, N, Gracovetsky, S. The biomechanics of the thoracolumbar fascia. Clin Biomech (Bristol, Avon). 1987;2:78–83.
100. Macintosh, JE, Valenica, F, Bogduk, N. The morphology of the human lumbar multifidus. Clin Biomech. 1986;1:196–204.
101. Martimo KP, Verbeek J, Karppinen J, et al: Manual material handling advice and assistive devices for preventing and treating back pain in workers. Cochrane Database Syst Rev 3:CD005958, 2007.
102. McGill, SM. Biomechanics of the thoracolumbar spine. In: Dvir Z, ed. Clinical biomechanics. Philadelphia: Churchill Livingstone, 2000.
103. McGill, SM. Kinetic potential of the lumbar trunk musculature about three orthogonal orthopaedic axes in extreme postures. Spine. 1991;16:809–815.
104. McGill, SM, Hughson, RL, Parks, K. Changes in lumbar lordosis modify the role of the extensor muscles. Clin Biomech (Bristol, Avon). 2000;15:777–780.
105. McGill, SM, Norman, RW. Reassessment of the role of intra-abdominal pressure in spinal compression. Ergonomics. 1987;30:1565–1588.
106. McGill, SM, Patt, N, Norman, RW. Measurement of the trunk musculature of active males using CT scan radiography: implications for force and moment generating capacity about the L4/L5 joint. J Biomech. 1988;21:329–341.
107. McGill, SM, Santaguida, L, Stevens, J. Measurement of the trunk musculature from T5 to L5 using MRI scans of 15 young males corrected for muscle fibre orientation. Clin Biomech (Bristol, Avon). 1993;8:171–178.
108. McLean, L. The effect of postural correction on muscle activation amplitudes recorded from the cervicobrachial region. J Electromyogr Kinesiol. 2005;15:527–535.
109. Moroney, SP, Schultz, AB, Miller, JA. Analysis and measurement of neck loads. J Orthop Res. 1988;6:713–720.
110. Moseley, GL, Hodges, PW, Gandevia, SC. External perturbation of the trunk in standing humans differentially activates components of the medial back muscles. J Physiol. 2003;547:581–587.
111. Nachemson, AL, Andersson, BJ, Schultz, AB. Valsalva maneuver biomechanics. Effects on lumbar trunk loads of elevated intraabdominal pressures. Spine. 1986;11:476–479.
112. National Institute for Occupational Safety and Health (NIOSH): The National Occupational Exposure Survey. Report No. 89-103. 1989. Cincinnati, NIOSH
113. Nitz, AJ, Peck, D. Comparison of muscle spindle concentrations in large and small human epaxial muscles acting in parallel combinations. Am Surg. 1986;52:273–277.
114. Nordin, M, Kahanovitz, N, Verderame, R, et al. Normal trunk muscle strength and endurance in women and the effect of exercises and electrical stimulation. Part 1: Normal endurance and trunk muscle strength in 101 women. Spine. 1987;12:105–111.
115. Oh, JS, Cynn, HS, Won, JH, et al. Effects of performing an abdominal drawing-in maneuver during prone hip extension exercises on hip and back extensor muscle activity and amount of anterior pelvic tilt. J Orthop Sports Phys Ther. 2007;37:320–324.
116. Panjabi, MM, Cholewicki, J, Nibu, K, et al. Critical load of the human cervical spine: an in vitro experimental study. Clin Biomech (Bristol, Avon). 1998;13:11–17.
117. Panjabi, MM, Ivancic, PC, Maak, TG, et al. Multiplanar cervical spine injury due to head-turned rear impact. Spine. 2006;31:420–429.
118. Patwardhan, AG, Havey, RM, Ghanayem, AJ, et al. Load-carrying capacity of the human cervical spine in compression is increased under a follower load. Spine. 2000;25:1548–1554.
119. Persing, J, James, H, Swanson, J, et al. Prevention and management of positional skull deformities in infants. American Academy of Pediatrics Committee on Practice and Ambulatory Medicine, Section on Plastic Surgery and Section on Neurological Surgery. Pediatrics. 2003;112:199–202.
120. Porterfield, JA, DeRosa, C. Mechanical low back pain: perspectives in functional anatomy. Philadelphia: Saunders; 1998.
121. Potvin, JR, McGill, SM, Norman, RW. Trunk muscle and lumbar ligament contributions to dynamic lifts with varying degrees of trunk flexion. Spine. 1991;16:1099–1107.
122. Reese, NB. Muscle and sensory testing. Philadelphia: Saunders; 1999.
123. Richardson, C, Hodges, PW, Hides, JA. Therapeutic exercise for lumbopelvic stabilization, ed 2. St. Louis: Churchill Livingstone; 2004.
124. Richardson, CA, Snijders, CJ, Hides, JA, et al. The relation between the transversus abdominis muscles, sacroiliac joint mechanics, and low back pain. Spine. 2002;27:399–405.
125. Rohlmann, A, Graichen, F, Bergmann, G. Loads on an internal spinal fixation device during physical therapy. Phys Ther. 2002;82:44–52.
126. Saeed, NR, Wall, SA, Dhariwal, DK. Management of positional plagiocephaly. Arch Dis Child. 2008;93:82–84.
127. Santaguida, PL, McGill, SM. The psoas major muscle: a three-dimensional geometric study. J Biomech. 1995;28:339–345.
128. Sato, H, Kikuchi, S. The natural history of radiographic instability of the lumbar spine. Spine. 1993;18:2075–2079.
129. Schipplein, OD, Trafimow, JH, Andersson, GB, Andriacchi, TP. Relationship between moments at the L5/S1 level, hip and knee joint when lifting. J Biomech. 1990;23:907–912.
130. Smith, CE, Nyland, J, Caudill, P, et al. Dynamic trunk stabilization: a conceptual back injury prevention program for volleyball athletes. J Orthop Sports Phys Ther. 2008;38:703–720.
131. Snijders, CJ, Ribbers, MT, de Bakker, HV, et al. EMG recordings of abdominal and back muscles in various standing postures: validation of a biomechanical model on sacroiliac joint stability. J Electromyogr Kinesiol. 1998;8:205–214.
132. Standring, S. Gray’s anatomy: the anatomical basis of clinical practice, ed 40. St Louis: Elsevier; 2009.
133. Stemper, BD, Yoganandan, N, Pintar, FA, Rao, RD. Anterior longitudinal ligament injuries in whiplash may lead to cervical instability. Med Eng Phys. 2006;28:515–524.
134. Stevenson, J, Bryant, T, Greenhorn, D, et al. The effect of lifting protocol on comparisons with isoinertial lifting performance. Ergonomics. 1990;33:1455–1469.
135. Tarnanen, SP, Ylinen, JJ, Siekkinen, KM, et al. Effect of isometric upper-extremity exercises on the activation of core stabilizing muscles. Arch Phys Med Rehabil. 2008;89:513–521.
136. Tesh, KM, Dunn, JS, Evans, JH. The abdominal muscles and vertebral stability. Spine. 1987;12:501–508.
137. Teyhen, DS, Gill, NW, Whittaker, JL, et al. Rehabilitative ultrasound imaging of the abdominal muscles. J Orthop Sports Phys Ther. 2007;37:450–466.
138. Teyhen, DS, Rieger, JL, Westrick, RB, et al. Changes in deep abdominal muscle thickness during common trunk-strengthening exercises using ultrasound imaging. J Orthop Sports Phys Ther. 2008;38:596–605.
139. Turk, AE, McCarthy, JG, Thorne, CH, Wisoff, JH. The “back to sleep campaign” and deformational plagiocephaly: is there cause for concern? J Craniofac Surg. 1996;7:12–18.
140. Tveit, P, Daggfeldt, K, Hetland, S, Thorstensson, A. Erector spinae lever arm length variations with changes in spinal curvature. Spine. 1994;19:199–204.
141. Urquhart, DM, Hodges, PW. Differential activity of regions of transversus abdominis during trunk rotation. Eur Spine J. 2005;14:393–400.
142. Urquhart, DM, Hodges, PW, Story, IH. Postural activity of the abdominal muscles varies between regions of these muscles and between body positions. Gait Posture. 2005;22:295–301.
143. van Dieen, JH, Hoozemans, MJ, Toussaint, HM. Stoop or squat: a review of biomechanical studies on lifting technique. Clin Biomech (Bristol, Avon). 1999;14:685–696.
144. Van, K, Hides, JA, Richardson, CA. The use of real-time ultrasound imaging for biofeedback of lumbar multifidus muscle contraction in healthy subjects. J Orthop Sports Phys Ther. 2006;36:920–925.
145. Vasavada, AN, Li, S, Delp, SL. Influence of muscle morphometry and moment arms on the moment-generating capacity of human neck muscles. Spine. 1998;23:412–422.
146. Vasseljen, O, Dahl, HH, Mork, PJ, Torp, HG. Muscle activity onset in the lumbar multifidus muscle recorded simultaneously by ultrasound imaging and intramuscular electromyography. Clin Biomech (Bristol, Avon). 2006;21:905–913.
147. Vera-Garcia, FJ, Brown, SH, Gray, JR, McGill, SM. Effects of different levels of torso coactivation on trunk muscular and kinematic responses to posteriorly applied sudden loads. Clin Biomech (Bristol, Avon). 2006;21:443–455.
148. Vera-Garcia, FJ, Elvira, JL, Brown, SH, McGill, SM. Effects of abdominal stabilization maneuvers on the control of spine motion and stability against sudden trunk perturbations. J Electromyogr Kinesiol. 2007;17:556–567.
149. Ward, SR, Kim, CW, Eng, CM, et al. Architectural analysis and intraoperative measurements demonstrate the unique design of the multifidus muscle for lumbar spine stability. J Bone Joint Surg Am. 2009;91:176–185.
150. Waters, TR, Lu, ML, Occhipinti, E. New procedure for assessing sequential manual lifting jobs using the revised NIOSH lifting equation. Ergonomics. 2007;50:1761–1770.
151. Waters, TR, Putz-Anderson, V, Garg, A, Fine, LJ. Revised NIOSH equation for the design and evaluation of manual lifting tasks. Ergonomics. 1993;36:749–776.
152. Welbergen, E, Kemper, HC, Knibbe, JJ, et al. Efficiency and effectiveness of stoop and squat lifting at different frequencies. Ergonomics. 1991;34:613–624.
153. Wilke, HJ, Rohlmann, A, Neller, S, et al. ISSLS prize winner: A novel approach to determine trunk muscle forces during flexion and extension—a comparison of data from an in vitro experiment and in vivo measurements. Spine. 2003;28:2585–2593.
154. Wilke, HJ, Wolf, S, Claes, LE, et al. Stability increase of the lumbar spine with different muscle groups. A biomechanical in vitro study. Spine. 1995;20:192–198.
155. Willinger, M, Hoffman, HJ, Wu, KT, et al. Factors associated with the transition to nonprone sleep positions of infants in the United States: the National Infant Sleep Position Study. JAMA. 1998;280:329–335.
156. Zhao, F, Pollintine, P, Hole, BD, et al. Discogenic origins of spinal instability. Spine. 2005;30:2621–2630.
STUDY QUESTIONS
1. Describe the most likely craniocervical posture resulting from (a) unilateral and (b) bilateral spasm (or shortening) in the sternocleidomastoid muscle(s).
2. Why are the superficial and intermediate muscles of the posterior back classified as “extrinsic” muscles? Describe how the specific innervation of these muscles is associated with this classification.
3. List structures that receive sensory innervation from the recurrent meningeal nerve. What nerves provide sensory innervation to the capsule of the apophyseal joints?
4. Justify why an isolated strong contraction of the semispinalis thoracis would likely produce contralateral axial rotation, whereas a strong isolated contraction of the longissimus cervicis or capitis would likely produce ipsilateral axial rotation. Use Figures 10-7 and 10-9 as a reference for answering this question.
5. Assume a person has a complete spinal cord injury at the level of T8. Based on your knowledge of muscle innervation, predict which muscles of the trunk would be unaffected and which would be partially or completely paralyzed. Consider only the abdominal muscles, multifidi, and erector spinae in your response.
6. List three muscles that attach to anterior tubercles and three that attach to posterior tubercles of transverse processes of cervical vertebrae. What important structure passes between these muscle attachments?
7. As a group, the trunk extensor muscles produce greater maximal-effort torque than the trunk flexor muscles (abdominals). Cite two factors that can account for this difference in strength.
8. Which of the major trunk muscles would experience the most significant stretch (elongation) after a motion of full trunk extension, right lateral flexion, and right axial rotation?
9. Based on Figure 10-16, which muscle has the greatest moment arm for (a) flexion and (b) lateral flexion at L3?
10. Describe how an overshortened (contracted) iliacus muscle can cause an increased lumbar lordosis while a person is standing. What effect could this posture have on the stress at the lumbosacral junction?
11. At the level of the third lumbar vertebra, which connective tissues form the anterior rectus sheath (of the abdominal wall)?
12. What is the primary difference between a dorsal ramus of a spinal nerve root and a dorsal nerve root?
13. Using Figure 10-23 as a reference, which muscle, based solely on its size, would theoretically produce the greatest extension force across the atlanto-occipital joints?
14. Describe the similarities and differences in the structure of the multifidi and the semispinales muscles.
15. As indicated in Figure 10-30, why is the axial rotation function of the rectus capitis posterior major muscle limited to the atlanto-axial joint only?
 Answers to the study questions can be found on the Evolve website.
Answers to the study questions can be found on the Evolve website.
[/level-membership-for-rheumatology-category][not-level-membership-for-rheumatology-category]Chapter 10
Axial Skeleton
Muscle and Joint Interactions
INNERVATION OF THE MUSCLES AND JOINTS WITHIN THE TRUNK AND CRANIOCERVICAL REGIONS
TRUNK AND CRANIOCERVICAL REGIONS
Special Considerations for the Study of Muscle Actions within the Axial Skeleton
Muscles of the Trunk: Anatomy and Their Individual Actions
Set 1: Muscles of the Posterior Trunk (“Back” Muscles)
Set 2: Muscles of the Anterior-Lateral Trunk (“Abdominal” Muscles)
Set 3: Additional Muscles (Iliopsoas and Quadratus Lumborum)
Muscles of the Trunk: Functional Interactions among Muscles
Muscles of the Craniocervical Region: Anatomy and Their Individual Actions
Muscles of the Craniocervical Region: Functional Interactions among Muscles That Cross the Craniocervical Region
SELECTED BIOMECHANICAL ISSUES OF LIFTING: A FOCUS ON REDUCING BACK INJURY
Muscular Mechanics of Extension of the Low Back during Lifting
Osteologic and arthrologic components of the axial skeleton are presented in Chapter 9. Chapter 10 focuses on the many muscle and joint interactions occurring within the axial skeleton. The muscles control posture and stabilize the axial skeleton, protect the spinal cord and internal organs, produce torques required for movement of the body as a whole, and, lastly, furnish fine mobility to the head and neck for optimal placement of the eyes, ears, and nose. Muscles associated with ventilation and mastication (chewing) are presented in Chapter 11.
INNERVATION OF THE MUSCLES AND JOINTS WITHIN THE TRUNK AND CRANIOCERVICAL REGIONS
An understanding of the organization of the innervation of the craniocervical and trunk muscles begins with an appreciation of the formation of a typical spinal nerve root (Figure 10-1). Each spinal nerve root is formed by the union of ventral and dorsal nerve roots: the ventral nerve roots contain primarily “outgoing” (efferent) axons that supply motor commands to muscles and other effector organs associated with the autonomic system. The dorsal nerve roots contain primarily “incoming” (afferent) dendrites, with the cell body of the neuron located in an adjacent dorsal root ganglion. Sensory neurons transmit information to the spinal cord from the muscles, joints, skin, and other organs associated with the autonomic nervous system.
Once a spinal nerve root exits its intervertebral foramen, it immediately divides into a ventral and dorsal ramus (the Latin word ramus means “path”) (see Figure 10-1). Depending on location, the ventral ramus forms nerves that innervate, in general, the muscles, joints, and skin of the anterior-lateral trunk and neck, and the extremities. The dorsal ramus, in contrast, forms nerves that innervate, in general, the muscles, joints, and skin of the posterior trunk and neck. This anatomic organization is depicted generically by the illustration in Figure 10-2.
Ventral Ramus Innervation
INDIVIDUAL NAMED NERVES
Many of the ventral rami within the trunk and craniocervical regions do not join a plexus; rather, they remain as individual named nerves. Each of these nerves typically innervates only a part or a segment of a muscle or connective tissue. This is why, for instance, many muscles that extend across a large part of the axial skeleton possess multiple levels of segmental innervation. The two most recognized sets of individual segmental nerves derived from the ventral rami are the intercostal (thoracic) and the recurrent meningeal nerves (see Figure 10-2).
Intercostal Nerves (T1 to T12): Each of the 12 ventral rami of the thoracic spinal nerve roots forms an intercostal nerve, innervating an intercostal dermatome and the set of intercostal muscles that share the same intercostal space. (Refer to dermatome chart in Appendix III, Part B, Figure III-2.) The T1 ventral ramus forms the first intercostal nerve and part of the lower trunk of the brachial plexus. The ventral rami of T7-T12 also innervate the muscles of the anterior-lateral trunk (i.e., the “abdominal” muscles). The T12 ventral ramus forms the last intercostal (subcostal) nerve and part of the L1 ventral ramus of the lumbar plexus.
Recurrent Meningeal Nerves: A single recurrent meningeal (sinuvertebral) nerve branches off the extreme proximal aspect of each ventral ramus. After its bifurcation, the recurrent meningeal nerve courses back into the intervertebral foramen (hence the name “recurrent” [see Figure 10-2]). As a set, these often very small nerves provide sensory and sympathetic nerve supply to the meninges that surround the spinal cord, and to connective tissues associated with the interbody joints.23 Most notably, the recurrent meningeal nerve supplies sensation to the posterior longitudinal ligament and adjacent areas of the superficial part of the annulus fibrosus. Sensory nerves innervating the anterior longitudinal ligament reach the spinal cord via small branches from nearby ventral rami and adjacent sympathetic connections.23
Dorsal Ramus Innervation
A dorsal ramus branches from every spinal nerve root, innervating structures in the posterior trunk usually in a highly segmental fashion. With the exception of the C1 and C2 dorsal rami, which are discussed separately, all dorsal rami are smaller than their ventral rami counterparts (see Figure 10-2). In general, dorsal rami course a relatively short distance posteriorly (dorsally) before innervating selected adjacent muscles and connective tissues on the back of the trunk (Box 10-1).
TRUNK AND CRANIOCERVICAL REGIONS
The muscles of the axial skeleton are organized into two broad and partially overlapping areas: the trunk and the craniocervical region (Table 10-1). The muscles within each area are further organized into sets, based more specifically on their location.
TABLE 10-1.
Anatomic Organization of the Muscles of the Axial Skeleton*
*A muscle is classified as belonging to the “trunk” or “craniocervical region” based on the location of most of its attachments.
The muscles within each area of the body are presented in two sections, the first covering anatomy and individual muscle actions, and the second covering examples of the functional interactions among related muscles. Throughout this chapter, the reader is encouraged to consult Chapter 9 for a review of the pertinent osteology related to the attachments of muscles. Appendix III, Part C should be consulted for a summary of more detailed muscular anatomy and innervation of the muscles of the axial skeleton.
Production of Internal Torque
By convention, the “strength” of a muscle action within the axial skeleton is expressed as an internal torque, defined for the sagittal, frontal, and horizontal planes. Within each plane, the maximal internal torque potential is equal to the product of (1) the muscle force generated parallel to a given plane, and (2) the length of the internal moment arm available to the muscle (Figure 10-3).
The spatial orientation of a muscle’s line of force determines its effectiveness for producing a torque for a particular action. Consider, for example, the obliquus externus abdominis muscle producing a force across the lateral thorax, with a line of force oriented about 30 degrees from the vertical (Figure 10-4). The muscle’s resultant force vector can be trigonometrically partitioned into unequal vertical and horizontal force components. The vertical force component—about 86% of the muscle’s maximal force—is available for producing lateral flexion or flexion torques. The horizontal force component—about 50% of the muscle’s maximal force—is available for producing an axial rotation torque. (This estimation is based on the cosine and sine of 30 degrees, respectively.) For any muscle of the axial skeleton to contribute all its force potential toward axial rotation, its overall line of force must be directed solely in the horizontal plane. For a muscle to contribute all its force potential toward either lateral flexion or flexion-extension, its overall line of force must be directed vertically. (Realize, though, that a vertically oriented muscle cannot produce any axial rotation because it lacks the moment arm required to produce a torque in the horizontal plane. As described in Chapter 1, a muscle force is incapable of producing a torque within a given plane if it either parallels or pierces the associated axis of rotation.)
The lines of force of muscles that control movement of the axial skeleton have a spatial orientation that varies over a wide spatial spectrum, from nearly vertical to nearly horizontal. This fact is important with regard to a muscle or muscle group’s torque potential for a given action. For instance, because more of the total muscle mass of the trunk is biased vertically than horizontally, maximal efforts usually produce greater frontal and sagittal plane torques than horizontal plane torques.145
Muscles of the Trunk: Anatomy and Their Individual Actions
The following section describes the relationships between the anatomy and the actions of the muscles of the trunk. Musculature is divided into three sets: (1) muscles of the posterior trunk, (2) muscles of the anterior-lateral trunk, and (3) additional muscles (see Table 10-1).
SET 1: MUSCLES OF THE POSTERIOR TRUNK (“BACK” MUSCLES)
The muscles of the posterior trunk are organized into three layers: superficial, intermediate, and deep (see Table 10-1).
Muscles in the Superficial and Intermediate Layers of the Back: The muscles in the superficial layer of the back are presented in the study of the shoulder (see Chapter 5). They include the trapezius, latissimus dorsi, rhomboids, levator scapula, and serratus anterior. The trapezius and latissimus dorsi are most superficial, followed by the deeper rhomboids and levator scapula. The serratus anterior muscle is located more laterally on the thorax.
The muscles included in the intermediate layer of the back are the serratus posterior superior and the serratus posterior inferior. They are located just deep to the rhomboids and latissimus dorsi. The serratus posterior superior and inferior are thin muscles that contribute little to the movement or stability of the trunk. Their function is more likely related to the mechanics of ventilation and therefore is described in Chapter 11.
SPECIAL FOCUS 10-1 Muscles of the Superficial Layer of the Back: an Example of Muscles “Sharing” Actions between the Axial and Appendicular Skeletons
Chapter 5 describes the actions of the muscles of the superficial layer of the back, based on their ability to rotate the appendicular skeleton (i.e., humerus, scapula, or clavicle) toward a fixed axial skeleton (i.e., head, sternum, vertebral column, or ribs). The same muscles, however, are equally capable of performing the “reverse” action (i.e., rotating segments of the axial skeleton toward the fixed appendicular skeleton). This muscular action is demonstrated by highlighting the functions of the trapezius and rhomboids during use of a bow and arrow. As indicated in Figure 10-5, several muscles produce a force needed to stabilize the position of the scapula and abducted arm. Forces produced in the upper trapezius, middle trapezius, and rhomboids simultaneously rotate the cervical and upper thoracic spine to the left, indicated by the bidirectional arrows.23,82 This “contralateral” axial rotation effect is shown for C6 in the inset within Figure 10-5. As the muscle pulls the spinous process of C6 to the right, the anterior side of the vertebra is rotated to the left. The trapezius and rhomboids also stabilize the scapula against the pull of the posterior deltoid, long head of the triceps, and serratus anterior. The shared actions of these muscles demonstrate the inherent efficiency of the musculoskeletal system. In this example, a few muscles accomplish multiple actions across both the axial and the appendicular skeletons.
Muscles in the Deep Layer of the Back: Muscles in the deep layer of the back are the (1) erector spinae group, (2) transversospinal group, and (3) short segmental group (Table 10-2). The anatomic organization of the erector spinae and transversospinal groups is illustrated in Figure 10-6.
Although a few exceptions prevail, muscles in the deep layer of the back are innervated segmentally through the dorsal rami of spinal nerves.132 A particularly long muscle within the erector spinae group, for instance, is innervated by multiple dorsal rami throughout the spinal cord. A shorter muscle such as one multifidus, however, is innervated by a single dorsal ramus.100
Erector Spinae Group: The erector spinae are an extensive and rather poorly defined group of muscles that run on both sides of the vertebral column, roughly within one hand’s width from the spinous processes (Figure 10-7). Most are located deep to the posterior layer of thoracolumbar fascia (see Chapter 9) and the muscles in the intermediate and superficial layers of the back. The erector spinae consist of the spinalis, longissimus, and iliocostalis muscles. Each muscle is further subdivided topographically into three regions, producing a total of nine named muscles (see Table 10-2). Individual muscles overlap and vary greatly in size and length.23
The bulk of the erector spinae muscles have a common attachment on a broad and thick common tendon, located in the region of the sacrum (see Figure 10-7). This common tendon anchors the erector spinae to many locations (Box 10-2). From this common tendon arise three poorly organized vertical columns of muscle: the spinalis, longissimus, and iliocostalis.132 The general muscle attachments are described in the following sections; more specific attachments can be found in Appendix III, Part C.
Spinalis Muscles: Spinalis muscles include the spinalis thoracis, spinalis cervicis, and spinalis capitis. In general, this small and often indistinct (or missing) column of muscle arises from the upper part of the common tendon. The muscle ascends by attaching to adjacent spinous processes of most thoracic vertebrae or, in the cervical region, the ligamentum nuchae. The spinalis capitis, if present, often blends with the semispinalis capitis.132
Longissimus Muscles: The longissimus muscles include the longissimus thoracis, longissimus cervicis, and longissimus capitis. As a set, these muscles form the largest and most developed column of the erector spinae group. The fibers of the longissimus thoracic muscles fan cranially from the common tendon, attaching primarily to the posterior end of most ribs. In the neck, the longissimus cervicis angles slightly medially before attaching to the posterior tubercle of the transverse processes of the cervical vertebrae (see Figure 10-7). The longissimus capitis, in contrast, courses slightly laterally and attaches to the posterior margin of the mastoid process of the temporal bone. The slightly more oblique angulation of the superior portion of the longissimus capitis and cervicis suggests that these muscles assist with ipsilateral axial rotation of the craniocervical region.
Iliocostalis Muscles: The iliocostalis muscles include the iliocostalis lumborum, iliocostalis thoracis, and iliocostalis cervicis. This group occupies the most lateral column of the erector spinae. The iliocostalis lumborum arises from the common tendon and courses upward and slightly outward to attach lateral to the angle of the lower ribs. The iliocostalis thoracis continues vertically to attach just lateral to the angle of the middle and upper ribs. From this point, the iliocostalis cervicis continues cranially and slightly medially to attach to posterior tubercles of the transverse processes of the midcervical vertebrae, along with the longissimus cervicis.
Summary: The erector spinae muscles cross a considerable distance throughout the axial skeleton. This anatomic feature suggests a design more suited for control of gross movements across a large part of axial skeleton (such as extending the trunk while rising from a low chair) rather than finer movements at selected intervertebral junctions.15 Bilateral contraction of the erector spinae as a group extends the trunk, neck, or head (Figure 10-8).47 The muscles’ relatively large cross-sectional areas enable them to generate large extension torque across the axial spine, such as for lifting or carrying heavy objects.37
By attaching to the sacrum and to the pelvis, the erector spinae can anteriorly tilt the pelvis, thereby accentuating the lumbar lordosis. (Pelvic tilt describes a sagittal plane rotation of the pelvis around the hip joints. The direction of the tilt is indicated by the rotation direction of the iliac crests.) As depicted in Figure 10-8, A, the anterior pelvic tilt is accentuated by the increased tension in stretched hip flexor muscles, such as the iliacus.
Transversospinal Muscles: Located immediately deep to the erector spinae muscles is the transversospinal muscle group: the semispinalis, multifidi, and rotatores (Figures 10-9 and 10-10). Semispinalis muscles are located superficially; the multifidi, intermediately; and the rotatores, deeply.
FIGURE 10-10. A posterior view shows the deeper muscles within the transversospinal group (multifidi on entire left side of A; rotatores bilaterally in B). The muscles within the short segmental group (intertransversarius and interspinalis) are depicted in A and B, respectively. Note that intertransversarius muscles are shown for the right side of the lumbar region only. The levator costarum muscles are involved with ventilation and are discussed in Chapter 11. (Modified from Luttgens K, Hamilton N: Kinesiology: scientific basis of human motion, ed 9, Madison, Wis, 1997, Brown and Benchmark.)
The name transversospinal refers to the general attachments of most of the muscles (i.e., from the transverse processes of one vertebra to the spinous processes of a more superiorly located vertebra). With a few exceptions, these attachments align most muscle fibers in a cranial-and-medial direction. Many of the muscles within the transversospinal group are morphologically similar, varying primarily in length and in the number of intervertebral junctions that each muscle crosses (Figure 10-11). Although somewhat oversimplified, this concept can greatly assist in learning the overall anatomy and actions of these muscles.
Semispinalis Muscles: The semispinalis muscles consist of the semispinalis thoracis, semispinalis cervicis, and semispinalis capitis (see Figure 10-9). In general, each muscle, or main set of fibers within each muscle, crosses six to eight intervertebral junctions. The semispinalis thoracis consists of many thin muscle fasciculi, interconnected by long tendons. Muscle fibers attach from transverse processes of T6 to T10 to spinous processes of C6 to T4. The semispinalis cervicis, much thicker and more developed than the semispinalis thoracis, attaches from upper thoracic transverse processes to spinous processes of C2 to C5. Muscle fibers that attach to the prominent spinous process of the axis (C2) are particularly well developed, serving as important stabilizers for the suboccipital muscles (described ahead).
The semispinalis capitis lies deep to the splenius and trapezius muscles. The muscle arises primarily from upper thoracic transverse processes. The muscle thickens superiorly as it attaches to a relatively large region on the occipital bone, filling much of the area between the superior and inferior nuchal lines (see Figure 9-3).
The semispinalis cervicis and capitis are the largest muscles that cross the posterior side of the neck. Their large size and near-vertical fiber direction account for the fact that these muscles provide 35% to 40% of the total extension torque of the craniocervical region.145 Right and left semispinalis capitis muscles are readily palpable as thick and round cords on either side of the midline of the upper neck, especially evident in infants and in thin, muscular adults (Figure 10-12).
Multifidi: Multifidi are situated just deep to the semispinalis muscles. The plural “multifidi” indicates a collection of multiple fibers, rather than a set of individual muscles. All multifidi share a similar fiber direction and length, extending between the posterior sacrum and the axis (C2).6,23,132 In general, the multifidi originate from the transverse process of one vertebra and insert on the spinous process of a vertebra located two to four intervertebral junctions above (see Figure 10-10, A).
Multifidi are thickest and most developed in the lumbosacral region (see multiple attachments listed in Box 10-3).100 The overlapping fibers of the multifidi fill much of the concave space formed between the spinous and transverse processes. The multifidi provide an excellent source of extension torque and associated stability to the base of the spine. Excessive force in the lumbar multifidi—from either active contraction or protective spasm—may be expressed clinically as an exaggerated lordosis.
Rotatores: The rotatores are the deepest of the transversospinal group of muscles. Like the multifidi, the rotatores consist of a large set of individual muscle fibers. Although the rotatores exist throughout the entire vertebral column, they are best developed in the thoracic region (see Figure 10-10, B).132 Each fiber attaches between the transverse process of one vertebra and the lamina and base of the spinous process of a vertebra located one or two intervertebral junctions above. By definition, the rotator brevis spans one intervertebral junction, and the rotator longus spans two intervertebral junctions.
Summary: On average, the transversospinal muscles cross fewer intervertebral junctions than the erector spinae group. This feature suggests that, in general, the transversospinal muscles favor a design for producing relatively fine controlled movements and stabilizing forces across the axial skeleton.15,22
Contracting bilaterally, the transversospinal muscles extend the axial skeleton (see Figure 10-8, B). Increased extension torque exaggerates the cervical and lumbar lordosis and decreases the thoracic kyphosis. The size and thickness of the transversospinal muscles are greatest at either end of the axial skeleton. Cranially, the semispinalis cervicis and capitis are very well-developed extensors of the craniocervical region; caudally, the multifidi are very well-developed extensors of the lower lumbar region, accounting for two thirds of the muscular-based stability in this region.154
Contracting unilaterally, the transversospinal muscles laterally flex the spine; however, their leverage for this action is limited because of their close proximity to the vertebral column. The more obliquely oriented transversospinal muscles assist with contralateral axial rotation. From a relatively fixed transverse process, contraction of a single left multifidus or rotator longus, for example, can rotate a superiorly located spinous process toward the left and, as a result, rotate the anterior side of the vertebra to the right. Compared with all the trunk muscles, however, the transversospinal muscles are secondary axial rotators. The leverage for this rotation is relatively poor because of the muscle’s proximity to the vertebral column. (Compare the multifidi with the obliquus abdominis externus, for example, in Figure 10-3, C). Furthermore, the prevailing line of force of most transversospinal muscle fibers is directed more vertically than horizontally, thereby providing a greater force potential for extension than for axial rotation.
Short Segmental Group of Muscles: The short segmental group of muscles consists of the interspinalis and the intertransversarius muscles (see Figure 10-10). (The plural “interspinales and intertransversarii” is often used to describe all the members within the entire set of these muscles.) They lie deep to the transversospinal group of muscles. The name “short segmental” refers to the extremely short length and highly segmented organization of the muscles. Each individual interspinalis or intertransversarius muscle crosses just one intervertebral junction. These muscles are most developed in the cervical region, where fine control of the head and neck is so critical.132
Each right and left pair of intertransversarius muscles is located between adjacent transverse processes. The anatomy of the intertransversarii as a group is more complex than that of the interspinales.132 In the cervical region, for example, each intertransversarius muscle is divided into small anterior and posterior muscles, between which pass the ventral rami of spinal nerves.
Summary: The highly segmented nature of the interspinalis and intertransversarius muscles is ideal for fine motor control of the axial skeleton. Because these unisegmental muscles possess a relatively high density of muscle spindles, they likely provide the nervous system (and therefore other muscles) a rich source of sensory feedback, especially in the craniocervical region.23
SET 2: MUSCLES OF THE ANTERIOR-LATERAL TRUNK (“ABDOMINAL” MUSCLES)
The muscles of the anterior-lateral trunk include the rectus abdominis, obliquus externus abdominis, obliquus internus abdominis, and transversus abdominis (Figure 10-13). As a group, these muscles are often collectively referred to as the “abdominal” muscles. The rectus abdominis is a long straplike muscle located on both sides of the midline of the body. The obliquus externus abdominis, obliquus internus abdominis, and transversus abdominis—the lateral abdominals—are wide and flat, layered superficial to deep, across the anterior-lateral aspects of the abdomen.
The abdominal muscles have several important physiologic functions, including supporting and protecting abdominal viscera and increasing intrathoracic and intra-abdominal pressures. As will be further described in Chapter 11, increasing the pressures in these cavities assists with functions such as forced expiration of air from the lungs, coughing, defecation, and child birth. This chapter focuses more on the kinesiologic functions of the abdominal muscles.
Formation of the Rectus Sheaths and Linea Alba: The obliquus externus abdominis, obliquus internus abdominis, and transversus abdominis muscles from the right and left sides of the body fuse at the midline of the abdomen through a blending of connective tissues. Each muscle contributes a thin bilaminar sheet of connective tissue that ultimately forms the anterior and posterior rectus sheaths. As depicted in Figure 10-14, the anterior rectus sheath is formed from connective tissues from the obliquus externus abdominis and the obliquus internus abdominis muscles. The posterior rectus sheath is formed from connective tissues from the obliquus internus abdominis and transversus abdominis. Both sheaths surround the vertically oriented rectus abdominis muscle and continue medially to fuse with identical connective tissues from the other side of the abdomen. The connective tissues thicken and crisscross as they traverse the midline, forming the linea alba (the Latin word linea means “line,” and albus, “white”). The linea alba runs longitudinally between the xiphoid process and pubic symphysis and pubic crest.
Anatomy of the Abdominal Muscles: The rectus abdominis muscle consists of right and left halves, separated by the linea alba. Each half of the muscle runs longitudinally, widening as it ascends within an open sleeve formed between the anterior and posterior rectus sheaths. The muscle is intersected and reinforced by three fibrous bands, known as tendinous intersections. These bands blend with the anterior rectus sheath. The rectus abdominis arises from the region on and surrounding the crest of the pubis, and it attaches superiorly on the xiphoid process and cartilages of the fifth through seventh ribs.
The anatomic organization of the obliquus externus abdominis, obliquus internus abdominis, and transversus abdominis muscles is different from that of the rectus abdominis. As a group, the more laterally placed muscles originate laterally or posterior-laterally on the trunk and run in a different direction toward the midline, eventually blending with the linea alba and contralateral rectus sheaths (Table 10-3).
The obliquus externus abdominis (informally referred to as the “external oblique”) is the largest and most superficial of the lateral abdominal muscles. The external oblique muscle travels in an inferior-and-medial direction, similar to the direction of the hands placed diagonally in front pockets of pants. The obliquus internus abdominis (or less formally the “internal oblique”) is located immediately deep to the external oblique muscle, forming the second layer of the lateral abdominals. Fibers originate from the iliac crest and, to a varying degree, blend with the adjacent thoracolumbar fascia. From this lateral attachment point, the fibers course in a cranial-and-medial direction toward the linea alba and lower ribs. As evident in Figure 10-13, C, the inferior attachments of the internal oblique muscle extend to the inguinal ligament. The average fiber direction of the internal oblique muscle is nearly perpendicular to the average fiber direction of the overlying external oblique muscle.
The transversus abdominis is the deepest of the abdominal muscles. The muscle is also known as the “corset muscle,” reflecting its role in compressing the abdomen as well as stabilizing the lower back through attachments into the thoracolumbar fascia.132 Of all the abdominal muscles, the transversus abdominis has the most extensive and consistent attachments into the thoracolumbar fascia,136 followed closely by the internal oblique muscle.
Actions of the Abdominal Muscles: Bilateral action of the rectus abdominis and oblique abdominal muscles reduces the distance between the xiphoid process and the pubic symphysis. Depending on which body segment is the most stable, bilateral contraction of these abdominal muscles flexes the thorax and upper lumbar spine, posteriorly tilts the pelvis, or both. Figure 10-15 depicts a diagonally performed sit-up maneuver that places a relatively large demand on the oblique abdominal muscles. During a standard sagittal plane sit-up, however, the opposing axial rotation and lateral flexion tendencies of the various abdominal muscles are neutralized by opposing right and left muscles.
As described in Chapter 9, the axes of rotation for all motions of the vertebral column are located in the region of the interbody joints. The relative posterior placement of the axes relative to the trunk equips the abdominal muscles, most notably the rectus abdominis, with very favorable leverage for generating trunk flexion torque (Figure 10-16). Note in Figure 10-16 that, with the exception of the psoas major, all muscles have a moment arm to produce torques in both sagittal and frontal planes.
Contracting unilaterally, the abdominal muscles laterally flex the trunk. The external and internal obliques are particularly effective in this action owing to their relatively favorable leverage (i.e., long moment arms) (see Figure 10-16) and, as a pair, relatively large cross-sectional area. The combined cross-sectional area of the external and internal obliques at the level of the L4-L5 junction is almost twice that of the rectus abdominis muscle.106
Lateral flexion of the trunk often involves activation of both trunk flexor and extensor muscles. For example, lateral flexion against resistance to the right demands a contraction from the right external and internal oblique, right erector spinae, and right transversospinal muscles. Coactivation amplifies the total frontal torque while simultaneously stabilizing the trunk within the sagittal plane.12
By far, the internal and external oblique muscles are the most effective axial rotators of the trunk.7,12,73,141 The external oblique muscle is a contralateral rotator, and the internal oblique muscle is an ipsilateral rotator. The strong axial rotation potential of these muscles reflects their relatively large cross-sectional area and favorable leverage (see Figure 10-3, C for long moment arm length of the obliquus externus abdominis). During active axial rotation in a particular direction, the external oblique muscle on one side functions synergistically with the internal oblique on the other side.141 This functional synergy produces a diagonal line of force that crosses the midline through the muscles’ mutual attachment into the linea alba (see Figure 10-15). Contraction of the two muscles therefore reduces the distance between one shoulder and the contralateral iliac crest.
Several electromyographic (EMG) studies using intramuscular (fine-wire) electrodes demonstrate some degree of bilateral activation of the transversus abdominis during axial rotation.38,82,141 It has been shown that during axial rotation the middle and lower fibers of the transversus abdominis coactivate at slightly different times than the upper fibers.141 Although the exact role of the transversus abdominis during axial rotation is uncertain, the muscle appears to function more as a stabilizer for the oblique abdominal muscles than a torque generator of axial rotation. Bilateral activation of the transversus abdominis can stabilize the ribs, linea alba, and thoracolumbar fascia—areas that serve as attachments for the internal or external oblique muscles.
The torque demands placed on the axial rotators of the trunk vary considerably based on the nature of an activity and position of the body.12 Torque demands are relatively large during high-power axial rotations, such as sprinting, wrestling, and throwing a discus or javelin. The demands may be very low, however, during activities that involve slow twisting of the trunk while in an upright position, such as during walking over level surfaces.
SPECIAL FOCUS 10-2 Role of Trunk Extensors as “Rotational Synergists” to the Oblique Abdominal Muscles
The external and internal oblique muscles are the primary axial rotators of the trunk. Secondary axial rotators include the ipsilateral latissimus dorsi, the more oblique components of the ipsilateral iliocostalis lumborum, and the contralateral transversospinal muscles. These secondary axial rotators are also effective extensors of the trunk. During a strong axial rotation movement, these extensor muscles are able to offset or neutralize the potent trunk flexion potential of the oblique abdominal muscles.98,154 Without this neutralizing action, a strenuous action of axial rotation would automatically be combined with flexion of the trunk. The aforementioned extensor muscles resist the flexion tendency of the oblique abdominal muscles, but also contribute slightly to the axial rotation torque.
The multifidi muscles provide a particularly important element of extension stability to the lumbar region during axial rotation.128,156 Pathology involving the apophyseal joints or discs in the lumbar region may be associated with weakness, fatigue, or reflexive inhibition of these muscles. Without adequate activation from the multifidi during axial rotation, the partially unopposed oblique muscles would, in theory, create a subtle and undesirable flexion bias to the base of the spine.
Comparing Trunk Flexor versus Trunk Extensor Peak Torque: In the healthy adult, on average, the magnitude of a maximal-effort trunk flexion torque is typically less than maximal-effort trunk extension torque. Although data vary based on gender, age, history of back pain, and angular velocity of the testing device, the flexor-to-extensor torque ratios determined isometrically for the trunk and craniocervical regions are between 0.45 and 0.77.20,80,114,145 Although the trunk flexor muscles normally possess greater leverage for sagittal plane torque (see Figure 10-16), the trunk extensor muscles possess greater mass and, equally important, greater overall vertical orientation of muscle fibers.95,106 The typically greater torque potential of the trunk extensor muscles reflects the muscles’ predominant role in counteracting gravity, either for the maintenance of upright posture or for carrying loads in front of the body.
SET 3: ADDITIONAL MUSCLES (ILIOPSOAS AND QUADRATUS LUMBORUM)
Iliopsoas: The iliopsoas is a large muscle consisting of two parts: the iliacus and the psoas major (see Figure 12-27). As are most hip flexors, the iliopsoas is innervated by the femoral nerve, a large branch from the lumbar plexus. The iliacus has a proximal attachment on the iliac fossa and lateral sacrum, just anterior and superior to the sacroiliac joint. The psoas major attaches proximally to the transverse processes of T12 to L5, including the intervertebral discs. The two muscles fuse distal to the inguinal ligament and typically attach as a single tendon to the lesser trochanter of the femur.
The iliopsoas is a long muscle, exerting a potent kinetic influence across the trunk, lumbar spine, lumbosacral junction, and hip joints. Crossing anterior to the hip, it is a dominant flexor, drawing the femur toward the pelvis or the pelvis toward the femur. In the latter movement, the iliopsoas can anteriorly tilt the pelvis, a motion that increases the lordosis of the lumbar region (review in Figure 9-63, A). With muscular assistance from the abdominal muscles, a strong bilateral contraction of the iliopsoas can also rotate the pelvis and superimposed trunk over fixed femurs. Based on this ability, the iliopsoas is as much a respected trunk flexor as a hip flexor. This discussion resumes later in the chapter.
Function of the Psoas Major at the Lumbosacral Region: In the anatomic position the psoas major demonstrates leverage for lateral flexion of the lumbar spine (see Figure 10-16).95 Little, if any, leverage exists for axial rotation.
The flexor and extensor capacity of the psoas major differs throughout the lumbosacral region. Across the L5-S1 junction, the psoas major has an approximate 2-cm moment arm for flexion (Figure 10-17).107 The psoas major is therefore an effective flexor of the lower end of the lumbar spine relative to the sacrum. Progressing superiorly toward L1, however, the line of force of the psoas major gradually shifts slightly posterior, falling either through or just posterior to the multiple medial-lateral axes of rotation (see cross-section at L3 in Figure 10-16). The muscle’s location reduces or eliminates its flexor or extensor capacity. The psoas major therefore is neither a dominant flexor nor extensor of the lumbar region, but rather a dominant vertical stabilizer of the region.57,127 (The term “vertical stabilizer” describes a muscular function of stabilizing a region of the axial skeleton in a near-vertical position while maintaining its natural physiologic curve.) Because of the lack of effective leverage in the lumbar region, the psoas major has a minimal role in directly influencing the degree of lordosis.127 The iliopsoas, however (as all hip flexor muscles), can indirectly increase the lordotic posture of the lumbar spine by tilting the pelvis anteriorly across the hip joints.
Quadratus Lumborum: Anatomically, the quadratus lumborum is considered a muscle of the posterior abdominal wall. The muscle attaches inferiorly to the iliolumbar ligament and iliac crest and superiorly to the twelfth rib and the tips of the stout transverse processes of L1 to L4 (Figure 10-18). The relative thickness of the muscle is evident by viewing Figure 10-16. The quadratus lumborum is innervated by the ventral rami of spinal nerves T12-L3.
Contracting bilaterally, the quadratus lumborum is an extensor of the lumbar region. Its action is based on the line of force passing about 3.5 cm posterior to the medial-lateral axis of rotation at L3.107
Contracting unilaterally, the quadratus lumborum has very favorable leverage as a lateral flexor of the lumbar region.57 The axial rotation potential of the quadratus lumborum, however, is minimal.
The psoas major and the quadratus lumborum run nearly vertically on both sides of the lumbar vertebrae (see Figure 10-16). A strong bilateral contraction of these muscles affords excellent vertical stability throughout the lumbar spine, including the L5-S1 junction. Theoretically, exercises that increase the volitional control and conditioning of these muscles may benefit a person with pain related to instability of the lumbar region.
Muscles of the Trunk: Functional Interactions among Muscles
Thus far in this chapter, the discussion of the muscles of the trunk has focused primarily on their anatomy and, for the most part, individual actions (Table 10-4). The upcoming discussion pays more attention to the functional interactions among the muscles or muscle groups. Two themes are explored: (1) muscular-based stability of the trunk, and (2) muscular kinesiology of performing a standard sit-up movement. The second interaction exemplifies a classic kinesiologic relationship between the trunk and hip muscles.
TABLE 10-4.
Actions of Most Muscles of the Trunk
[/not-level-membership-for-rheumatology-category]
