CHAPTER 50 Animal Models of Epilepsy
Epilepsy affects a sizable proportion of the population worldwide and is responsible for a heavy social and economic burden.1 Given that epileptic disorders frequently result in uncontrolled and often medically intractable seizures, further experimental studies offer hope for a deeper understanding of the underlying pathophysiology and, ultimately, better treatments. To achieve these goals, human studies, albeit of great value, may not be sufficient because of both ethical and practical limitations.2 Accordingly, it is not surprising that animal models were developed and have significantly contributed to the epilepsy research literature. A common concern with all animal models is how reliable they are in mimicking the human condition. Are the anatomic and electrophysiologic similarities with human epileptic disorders real or superficial? To what extent can findings in animal research be extended to humans? Furthermore and perhaps most important, what new insights can be gained from animal model studies that cannot be predicted from clinical studies? In the design and interpretation of animal models, it is important to remember that epilepsy is not a single disease, a syndrome, or a homogeneous entity. Although a common feature of epilepsy is the tendency to have spontaneous epileptic seizures,3 the many ways in which seizures are generated (and manifested) are quite varied. Seizures can be motor, sensory, or autonomic and are caused by excessive and abnormal neuronal discharge. Causes of epilepsy range from genetic to acquired to “of unknown origin” (i.e., idiopathic).4 Its symptoms are also many because they depend on the brain areas that are involved in a certain type of seizure and the stage of brain maturation. The type (or types) of seizures that one individual has and other symptoms that are also present can be used to define a specific epileptic syndrome. Because epilepsy involves many levels of structures and activity in the brain, be it from molecules to networks, with causes that range from genes to environmental insults, it is no surprise that many epilepsy models are needed.
A good model of epilepsy should reproduce as many salient aspects as possible for a specific type of human epilepsy.5,6 Ideally, there should be evidence in a model for spontaneous behavioral seizures of the kind (e.g., partial, absence, tonic-clonic) that its human counterpart exhibits, as well as electroencephalographic (EEG; ictal and interictal discharges, focal or generalized) and structural (if any) abnormalities similar to the ones seen in the human epilepsy that it mirrors. The etiology (e.g., cortical malformation [CM], genetic predisposition, focal gliosis) should, if possible, be the same in the animal model as in the human condition. If the human epilepsy has a specific age at onset, the model should do the same. The animal model should have behavioral characteristics (such as memory deficits or developmental retardation) that parallel the human condition. Finally, the model in question should respond to antiepileptic drugs (AEDs) in a manner similar to the human condition after which it is modeled. Even though these goals are scientifically sound, few (if any) animal models actually fulfill all these criteria. Here we review animal models designed to mimic three broad classes of epileptic disorders: (1) temporal lobe epilepsy (TLE), (2) epilepsy associated with a brain malformation, and (3) other focal epilepsies.
Temporal Lobe Epilepsy
Seizures that arise from the temporal lobe are very common in humans and are often resistant to pharmacologic treatment. The most common type of TLE is that associated with hippocampal sclerosis.7 Kindling, pilocarpine, and kainic acid (kainate) are the most common animal models of TLE.
Stimulus-Induced Model of Temporal Lobe Epilepsy
Kindling
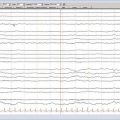
Kindling has been used as a model of seizures and epilepsy for more than 3 decades. Initially, these animals were not used for epilepsy, but rather as a physiologic model of learning and memory.8 This model is based on the kindling phenomenon, which involves the progressive development of EEG and behavioral seizures evoked by repeated electrical stimulation of certain brain structures. The first studies from the 1960s showed that repetitive electrical brain stimulation produces an increase in convulsive behavior and eventually generalized motor seizures.8–10 The progression of motor seizures proceeds through several stages as defined by Racine for the amygdala kindling protocol.11 With initial stimulation, there is an initial ictus with orofacial movements (stage 1) and head nodding (stage 2), which is associated with a focal EEG seizure. Further stimulation results in contralateral forelimb clonus (stage 3) and rearing (stage 4). Additional loss of balance, rearing, and falling (stage 5) are accompanied by tonic-clonic seizures involving all four limbs.12 If the kindling stimulus continues to be applied to the focus after a stage 5 seizure is reached, the behavioral sequence evolves further to more severe seizures. Thus, the more advanced the animal in this process, the more intense the seizures and the more widespread and prolonged the electrical seizures or afterdischarges (i.e., the most sensitive parameter of focal seizure activity in kindled rats). Kindling occurs best in neuroplastic areas of the brain; the amygdala is the brain region most susceptible to kindling.13
Although other, but not all encephalic structures can be kindled, there are some differences between them regarding the speed of the kindling process and seizure manifestation.14–16 Stimulation of the cerebellum and superior colliculus, for example, cannot start kindling. Kindling from the hippocampus produces seizures quite similar to but slower to develop than those induced by amygdala kindling. The profile of afterdischarges is different from amygdala kindling in that they recur daily during the kindling process until the seizures recruit other temporal areas.14,17 One peculiar early behavioral response seen during hippocampal kindling in some rats is the so-called wet dog shakes, which start and progressively disappear during the kindling process before the expression of any Racine motor stages.
The neurological alterations associated with the types of kindling behavior seem to be permanent. Kindled animals that have not been stimulated for many months after stage 5 kindling often respond with full seizure behavior on repeat exposure to the original kindling stimulus.8,18 Moreover, kindling of a second structure is usually achieved with fewer stimuli than needed for the primary kindled site.8,19,20 This “transfer phenomenon” might be the physiologic basis for secondary epileptogenesis.21–23 In contrast, if two different nonhomologous areas receive concurrent and alternate stimulation, only one of them undergoes a typical kindling progression, and seizure genesis in the other site is suppressed. This “kindling antagonism” may involve inhibitory mechanisms recruited by the seizure activity from the more dominant site24,25 and could reflect the types of endogenous compensatory mechanisms that the brain has evolved to combat seizure activity.
When the amygdala or other brain areas such as the frontal and posterior cortex, entorhinal cortex, and perforant pathway are kindled, spontaneous seizures can develop in the animals after a period. However, just a few kindled animals reach this point.26,27 Recent evidence from the laboratory of Dan McIntyre suggests that there may be a genetic explanation for this interesting observation. Over time, they isolated strains of “fast”- and “slow”-kindling rats, with the former being highly susceptible to kindling treatment and the latter more resistant.28–30 These rats exhibit different seizure predispositions and behavior that might be attributed to differences in the expression of GABAA receptor subunits and in the kinetics of miniature inhibitory postsynaptic currents in different limbic structures.28,31–33 Behaviorally, the fast-kindling rat strains are more prone than the slow-kindling strains to the development of seizures and appear much like humans with attention-deficit/hyperactivity disorder in that they show easy distraction, hyperactivity, and impulsivity.32,34,35 Importantly, spontaneous seizures in kindled animals are not just a rat phenomenon inasmuch as epilepsy has developed in every animal tested thus far with this stimulation protocol: frogs, reptiles, mice, rats, rabbits, cats, dogs, rhesus monkeys, and baboons.36–38 A persistent question with this model is whether a kindling-like phenomenon occurs in patients with epilepsy. This discussion was initiated by Gowers, who affirmed that “seizures do beget seizures.” In other words, epilepsy could be a progressive condition wherein the chronic recurrence of seizures would increase the likelihood of new seizures, and they could come from new areas (e.g., homologous contralateral sites). There are some hints in epilepsy associated with brain tumors that secondary epileptogenesis does occur in humans.23 Although deliberately causing kindling in humans would be ethically suspicious, to say the least, some reports of such do exist in the literature.39
As a model of epilepsy, the kindling model has been successfully used as a screening tool for AEDs. Lamotrigine, used as an AED in patients with partial and generalized seizures, has long-acting anticonvulsant effects on both amygdala- and hippocampal-kindled seizures in rats.40 GABA uptake inhibitors, such as tiagabine, were also found to be highly effective in the kindling model,41 consistent with the GABA receptor involvement described earlier. Another example is levetiracetam, a relatively new AED that is effective in the kindling, pilocarpine, and kainate models of TLE and in human TLE.42–45 Interestingly, these three drugs are inactive against seizures induced by maximum electroshock (MES), a model of acute seizures in nonepileptic animals and additionally used to screen for potential AEDs.41 More recently, amygdala kindling has also been proposed as an animal model for drug-resistant epilepsy.46 Studies have shown that kindled seizures are less sensitive than primarily generalized seizures to anticonvulsant treatment, as demonstrated with the MES test.41 The response to phenytoin, for example, differs in kindled rats (i.e., in some animals phenytoin has an anticonvulsant effect consistently and in others never).47 Unfortunately, the preparation of kindled rats and drug testing in this model are both labor and time intensive, which limits its use in high-throughput drug discovery and development programs. Some less laborious models are discussed in this respect, including corneal kindling, acute models of focal seizures (e.g., electrical induction of afterdischarges), and genetic models. However, when compared with conventional amygdala kindling, none of these models have been characterized sufficiently as yet to judge their usefulness for drug development.48
Finally, the kindling model offers some distinct advantages over systemically drug-induced models of TLE. With kindling, one knows precisely where the epileptogenesis begins (i.e., the site and intensity of the stimuli applied) and can track by numerous methods the progressive changes that occur during a very prolonged latent period, which ends with spontaneous seizures. Additionally, gross brain damage does not appear to be a prerequisite for the initial manifestation of spontaneous seizures.26 After repeated secondary generalized seizures evoked by kindling, cumulative neuronal loss can be detected in the CA1 and CA3 subfields of the hippocampus and in the hilus of the dentate gyrus (DG), similar to classic human hippocampal sclerosis, which is associated with long-term seizure-induced memory dysfunction.49–51 Although kindling does not result in a large loss of neurons, there is clear reorganization of the neuronal circuitry. This reorganization is based on synaptogenesis52 or sprouting of mossy fibers (or both) in the inner molecular layer of the DG53 and in the stratum oriens of the CA3 hippocampal area.54 In addition, kindling-induced seizures result in changes in the morphology of astrocytes that (1) appear to be dependent on seizure intensity, (2) occur early in the kindling process, and (3) persist for weeks after the last seizure. An increase in astrocytic proliferation in the hippocampus, amygdala, and piriform cortex was also observed after kindling.55 Some of these structural alterations induced by kindling are similar to those in the human epileptic temporal lobe, thus raising the possibility that mechanisms operative in kindling may play a role in the pathogenesis of human TLE associated with hippocampal sclerosis.56 As one might expect, the seizure progression and structural changes are accompanied by progressive functional alterations in the kindled animal. These changes have been particularly well studied in the DG. When the first seizure is induced, there is an increase in excitability through N-methyl-D-aspartate (NMDA)-mediated synaptic transmission. Even though this increment is not permanent,57 it appears to play an important long-term role by initiating structural and functional modifications that contribute to long-lasting seizure susceptibility in the hippocampal circuitry.58 Neurochemical and neurophysiologic studies have indicated that the activity of the GABAergic projection from the striatum to the substantia nigra pars reticulata is reduced and the density of GABAergic striatal interneurons is increased in kindled rats.59–62 In addition, GABAergic inhibition is altered in the piriform cortex of kindled rats.63
Chemical-Induced Models of Temporal Lobe Epilepsy
Pilocarpine and Kainate
Pilocarpine and kainate models replicate several phenomenologic features of human TLE, and both can be used as animal preparations to understand the basic mechanisms of epileptogenesis.64–66 Pilocarpine is a cholinergic muscarinic agonist originally isolated from the leaflets of South American shrubs (Pilocarpus jaborandi). The first evidence that the brain damage produced by pilocarpine-induced status epilepticus (SE) could lead to spontaneous recurrent seizures (SRSs) in rats over a long-term period occurred in 1983.67 Kainate is an excitatory amino acid extracted from seaweed (Digenea simplex) and has been used as a model of epilepsy since the late 1970s.68,69 Local or systemic administration of either pilocarpine or kainate to rodents leads to a pattern of repetitive limbic seizures and SE that can last for several hours.67,70,71 Systemic administration of a single dose of pilocarpine or repetitive low doses of kainate to rodents leads to a sequence of seizures that build progressively into a limbic SE lasting up to 24 hours (acute period). In these models, induction of SE is critical for the later development of SRSs. By managing the severity of SE one can decrease animal mortality without decreasing the development of recurrent epilepsy. For pilocarpine, this can be accomplished by using thionembutal or diazepam after the beginning of SE. Pretreatment with scopolamine also minimizes the peripheral cholinergic side effects of pilocarpine.72,73 In some protocols, lithium is applied before pilocarpine to increase its effect and reduce mortality.74 For kainate, a multiple low-dose protocol may decrease mortality and produce animals with robust SRSs.75,76 SE induced by kainate can also be abolished by diazepam. However, some animals may die before the benzodiazepine injection, and this approach may lead to a lower percentage of animals in which robust and long-lasting convulsive SE and subsequent SRSs develop. Seizures during the acute period are similar with both drugs. After injection of pilocarpine, the animal begins with staring and facial automatisms, followed by motor limbic seizures with rearing, forelimb clonus, salivation, intense masticatory movements, and falls. This progression follows the Racine stages.77 For kainate, the seizures are characterized by wet dog shakes, staring, searching, and gnawing, subsequently leading to forelimb clonus (stage 3 of Racine) and followed by Racine stages 4 and 5.78 The electrographic pattern during the acute period is characterized by theta rhythm activity in the hippocampus and low-voltage fast activity in the cortex with progression to high-voltage fast activity with spikes in the hippocampus. Sequentially, spiking activity spreads to the cortex and evolves into electrographic recurrent seizures that build up to continuous epileptiform activity.71,79 This acute period is followed by a “silent period” (seizure-free phase) with progressive normalization of EEG activity and behavior of variable extent that lasts from days to weeks. This sort of latent interval is believed to play an important role in “ripening of the focus,” a process that renders the animal chronically epileptic.80 After this latent period, animals begin to have frequent SRSs (chronic period).67,70,73,81–83 The main features of the SRSs observed during the long-term period resemble those of human complex partial seizures.66,67,79,84 SRSs are characterized by facial automatisms, head nodding, forelimb clonus, rearing, and falling and electrographically by paroxysmal hippocampal discharges that rapidly spread to cortical regions. The ensuing spontaneous seizures show gradual electrographic synchronization of cortical and hippocampal activities and a longer duration of ictal events. Forelimb clonus and rearing with falling (kindling stages 4 or 5) are the clinical hallmarks of this SRS phase.71,79
As models for temporolimbic epilepsy, the pilocarpine and kainate models are important instruments for screening new AEDs. Studies have shown that diazepam, phenobarbital, valproic acid, and trimethadione protect against acute pilocarpine-induced SE whereas phenytoin and carbamazepine are ineffective.66 In contrast, during the chronic period, phenobarbital, carbamazepine, phenytoin, and valproic acid but not ethosuximide are able to control pilocarpine-induced SRSs. Valproic acid was likewise able to abolish SRSs in kainate-treated animals.85 These AEDs can also control complex partial seizures in humans.86 SRSs may thus be more reliable than acute seizure models for finding new AEDs with better efficacy against complex partial seizures. In the kainate model, a ketogenic diet also seems to be effective in reducing SRSs.87
In hippocampal tissue removed from patients with refractory TLE, neuronal loss is most prominent in the hippocampal formation, particularly in the DG and the CA1 and CA3 subfields.88–90 Mossy fiber sprouting,91,92 hippocampal gliosis,88,89 cell dispersion in the granular cell layer of the DG, and local ectopic areas91 are additional neuropathologies. Although these morphologic changes are much more prominent in pilocarpine- and kainate-treated rats than in humans,73 the structural changes in these models of epilepsy during the acute and chronic periods resemble the anatomic changes found in many patients with mesial TLE.64,65,67,69,73,93–95 Morphologic analysis of the rodent brain during the acute period shows that both kainate- and pilocarpine-induced SE invariably leads to cellular loss and injury in the hippocampal subfields CA1 and CA3 and in the DG hilus. Cell loss is also often seen in the septum, olfactory tubercle, amygdala, piriform cortex, neocortex, thalamic nuclei, and substantia nigra.67,96–98 In fact, the thalamus can exhibit severe atrophy in various nuclei.99 Damage to subpopulations of GABAergic inhibitory neurons is also extensive throughout the hippocampus.100 In the following months the cell damage tends to progressively involve other areas. In the chronic period, cell loss is often noted in the DG (mainly attributed to a decrement in GABAergic interneurons) and in the hippocampal subfields CA1 and CA3 (loss of pyramidal cells). The decreased subpopulation of interneurons in the DG is associated with a functional hyperexcitability.65 In the kainate model, the CA3 area seems to be preferentially damaged in comparison to area CA1, which probably reflects the higher distribution of hippocampal kainate receptors in CA3. This differs from the typical hippocampal sclerosis in humans, in which CA1 is usually more damaged than CA3. As in humans, the DG and the CA2 area are relatively spared.69,88,89,93 Cell dispersion is evident in the granule cell layer of the DG101; increased neurogenesis102 and supragranular and intragranular mossy fiber sprouting73,103,104 are also important features in these models during the period of SRSs. Interestingly, SRSs can occur in animals with complete absence of mossy fiber sprouting.105,106 These morphologic changes seem more likely to represent a reactive response rather than a direct consequence of the initial insult. As observed in kainate-treated rats, axonal sprouting and synaptic reorganization take place not only in the DG but also in other hippocampal areas, such as CA1 and CA3.107 These cellular and synaptic modifications are associated with neurochemical and cellular dysfunctions that lead to permanent hyperexcitability. For example, the decrease in GABAergic cells in the DG and entorhinal cortex in the pilocarpine model of epilepsy is associated with decreased inhibitory synaptic input to the granule cells and to the layer II stellate cells, respectively, with a consequent increase in network excitability. Reduced granule cell inhibition in the DG seems to precede the onset of SRSs by days to weeks.108,109 Decreased expression of some postsynaptic GABAA receptor subunits,110–116 loss of GABAergic interneurons in the entorhinal cortex, and changes in neuropeptide Y expression, including ectopic expression in the granule cells and mossy fibers, have also been observed in rats with pilocarpine-induced epilepsy.117 Some of these changes precede the onset of SRSs by weeks and correlate with profound alterations in receptor function that contribute to the process of epileptogenesis. Increased glutamate release in the hippocampus was also observed during the acute period118,119 and could further contribute to the epileptogenic process in this model. For example, constant activation of NMDA-type glutamate receptors, which leads to increased intracellular Ca2+ and consequent activation of lipases, proteases, and nucleases, can kill cells by necrosis, apoptosis, or both. Thus, activation of NMDA receptors plays an important role in SE and brain damage in these models.120 The expression of proteins related to ionotropic NMDA and metabotropic glutamate receptors in the hippocampus is also modified in the pilocarpine model of epilepsy.121,122
As much as the pilocarpine and kainate effects in rats can resemble the human condition, species differences between rodents and primates are frequently used to explain existing discrepancies. To this end, developmental changes are often called on to explain features such as tectonic malformations123 and DG cell dispersion91 in humans even though these findings have been demonstrated to take place in adult epileptic rats.101 As a means to more properly evaluate the extent to which rodent models mimic the human condition, pilocarpine has also been used in a nonhuman primate, the marmoset (Callithrix jachus).124,125
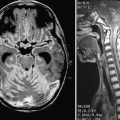
In seminal work by Luiz Mello and colleagues, administration of pilocarpine was used to induce SE in marmosets followed, after a silent period, by SRSs. It was observed from this model that prolonged SE is required to promote injury during the acute period and that the mortality rate is higher during SE in marmosets than in rats. That rodents seem to be more resistant to pilocarpine-induced SE (in wild types, 20% to 30% of animals will not reach full SE after drug administration) may be the reason that more damage is necessary for spontaneous seizures to develop. Different from the rodent pilocarpine and kainate models but similar to human TLE, brain damage is minimal during the chronic period in marmosets and generally limited to the limbic structures (mainly the hippocampus), and neurogenesis, exuberant in rodents,102 is mild in this primate.126 Cellular dispersion in the granular cell layer of the DG; neuronal loss in CA1, CA3, and the DG hilus; and supragranular mossy fiber sprouting were detected in marmosets with SRSs. Regarding morphology, one interesting observation in the marmoset pilocarpine model was the finding of tectonic malformations in the DG characterized by invaginations and lateral displacement of the granular cell layer, mainly in the posterior plates, similar to that described by Sloviter and coworkers in humans with TLE.123 This new model of TLE in marmosets may have a greater resemblance to the alterations seen in human TLE and may provide a new tool to more properly evaluate the causes of and consequences associated with TLE.
Cortical Malformations and Epilepsy
CMs are developmental neuronal disorders in humans characterized by (1) disorganization of cortical architecture or (2) the presence of abnormal, immature, and eventually nondifferentiated neurons (or both).127–130 Genetic or environmental prenatal factors can lead to focal or diffuse CMs, or both, which are often associated with refractory epilepsy.127–129 The anatomic features of CMs associated with epilepsy have been widely studied.131–134 Although no single animal model can reproduce the precise anatomic disorganization and cell abnormalities observed in humans, these models have proved useful in studying the functional characteristics of dysplastic neuronal networks. Some CM models are based on induced injury in the immature brain (e.g., freeze, undercut, irradiation, teratogen exposure); others are based on spontaneous genetic mutations or on trangenesis that gives rise to abnormal cortical architecture. Here we review some of the injury-based and genetic models of CMs.
Injury-Based Models of Cortical Malformations
Brain malformation can be induced in animals by teratogenic compounds or by mechanical treatment. These models include prenatal exposure (embryonic days 14 to 16) to methylazoxymethanol acetate (MAM)135 or to irradiation (γ- or x-rays)136 and the neonatal application (postnatal days 0 to 2) of a freeze lesion in the cortical plate of rodents.137,138 There are similarities in the embryonic development of the human and rat brain, and neurogenesis occurs in a relatively known spatiotemporal manner.139 Deleterious agents applied at selected stages of development can reduce neuronal populations in the injured brain regions,140 similar to those seen in some forms of CM in humans. This reduction and altered architecture subsequently lead to a focal or diffuse structural and functional impairment of the brain that contributes to epileptogenesis.
Methylazoxymethanol Acetate
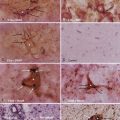
MAM is an antimitotic methylating agent known since the 1960s to induce brain malformations141–145 in rats when the exposure takes place at the beginning of the third week of intrauterine development. MAM has a short half-life and exerts an antiproliferative action on dividing, but not quiescent neuroepithelial cells via methylation of nucleic acids.135,143 Prenatal injection of MAM into female rats at gestational day 15 selectively affects the proliferation of specific neuronal cell populations by disrupting the sequence of normal brain development. The offspring resulting from prenatal MAM exposure exhibit multifocal brain malformations, microcephaly,141,146 loss of lamination, vascular abnormalities, and neuronal heterotopia in the hippocampal and periventricular locations.147–153 The abnormal cell clusters (heterotopia) first appear postnatally in the hippocampus (postnatal days 1 to 2), and their appearance is preceded by a distinct sequence of perturbations in neocortical development.154 These induced neuronal heterotopias possess many features of CMs associated with epilepsy in humans, such as focal cortical dysplasia, periventricular nodular heterotopia, and tectonic hippocampal malformations in patients with TLE.123,148,155 In addition, heterotopic neurons have an abnormal synaptic network, with neurons communicating directly between the neocortex and hippocampus,156 and they exhibit characteristics of neocortical cells. For example, molecular analysis has revealed that hippocampal heterotopic cells express mRNA markers normally found abundantly in layer II and III supragranular neocortical neurons and exhibit firing properties strikingly similar to those of supragranular cortical neurons.157 MAM-treated rats have increased susceptibility to induced seizures in vivo,158–160 and hyperexcitability in vitro is refractory to the commonly available AEDs.161 This hyperexcitability can be attributed to a combination of different cellular mechanisms.162,163 Functionally, heterotopic hippocampal cells (1) lack Kv4.2 A-type potassium channels,162 which results in abnormal neuronal firing, and (2) exhibit changes in glutamate receptor function and expression.164–166 The inhibitory synaptic system is also altered in hippocampal heterotopia, including changes in the inhibitory drive such as a prolonged duration of inhibitory synaptic events associated with a decrease in transporter-mediated GABA reuptake.131 These alterations in GABAergic inhibition could represent some type of postnatal compensatory response to the intrinsic hyperexcitability of these animals because spontaneous electrographic seizures, although present, are rare in MAM-exposed rats.165 Interestingly, some of these same deficits were found to occur in dysplastic tissue samples from patients with focal cortical dysplasia and epilepsy.132
Irradiation
Different radiation sources (x-rays, γ-rays) have been used to induce CMs in animal models, but the results have been similar in all cases. The histologic abnormalities in irradiated animals are attributed to the initial injury from irradiation associated with continued cortical development in an altered cellular environment. Immature, migrating neurons and radial glia are particularly sensitive to radiation.136,167 Different CMs were produced in rats exposed to x-rays in utero (200 cGy)168 and γ-irradiation (150 to 250 cGy). Pregnant rats exposed to external γ-irradiation on gestational day 16 to 17 produce offspring with various degrees of CMs and architectural abnormalities, depending on the dose of in utero radiation.169 Timing of the exposure is also critical,136 with exposure on gestational day 16 to 17 producing the most severe malformations. These abnormalities include microcephaly; diffuse CM with dyslamination and lack of columnar organization in multiple areas of the neocortex; subcortical, periventricular, and hippocampal neuronal heterotopias; and dysgenesis of the corpus callosum.169–172 Not only do radiation-exposed rats exhibit epileptiform discharges, but spontaneous seizures also develop in vivo.172–174 The epileptogenicity in vivo increased with mild to moderate radiation doses. However, high doses of in utero radiation leading to more severe pathologic changes in the neocortex and hippocampus are not associated with the occurrence of spontaneous seizures,173,175,176 again suggesting potential compensatory mechanisms or widespread brain damage that could preclude the ability to generate network activity. Extracellular recordings from brain slices of in utero irradiated rats showed enhanced epileptiform activity in the dysplastic areas in vitro.177 Anatomic reduction of GABAergic interneurons (parvalbumin- and calbindin-positive cells178) was associated with decreased inhibitory synaptic networks in the malformed cortical regions of irradiated animals134; reduced synaptic inhibition, measured in voltage-clamp experiments, was later confirmed in this model.
Freeze Lesion
Several studies reported on the development of focal cortical microgyria in the cortex of neonatal rats (up to 3 to 4 days postnatally) as a result of different types of injuries such as transcortical freeze lesions137,138 and focal injection of ibotenate (glutamate agonist).179 The induced microgyria in these models is characterized by four cortical layers instead of six, with an absence of layers V, VIa, and sometimes part or all of layer IV,138,180–182 and it mimics all histologic aspects of human four-layered polymicrogyria.183 One of the original models of focal CM is the neonatal freeze lesion model.184 The polymicrogyria resulting from a lesion induced by the application of a deep-freeze probe to the skull of newborn rats is believed to be caused by cell death secondary to focal hypoxia rather than a migratory defect.182,185 There is a correlation between the presence of layered polymicrogyria and hypoxic events occurring during the late stages of cortical migration.183,186,187 Hyperexcitability in vitro has been recorded in the paramicrogyral zone180,184,188–190 and is associated with an increase in NMDA-mediated excitation and a decrease in glutamatergic input onto inhibitory interneurons.191 An anatomic substrate for this hyperexcitability could be the increased expression of NMDA receptors within the microgyrus and α-amino-3-hydroxy-5-methyl-4-isoxazoleproprionate (AMPA) receptors in both the microgyral and paramicrogyral zones.192 An increase in excitatory glutamatergic projections from the thalamus has also been described in the area bordering the microgyrus.192 This rearrangement of thalamic afferents is attributed to the absence of layer IV neurons (their normal target) in the microgyric area.190,193 This excitatory input adjacent to the microgyrus may be increased as a result of abnormal circuitry in the paramicrogyral zone. In addition, variable decreases in inhibitory interneurons and GABAA binding194 were observed within the microgyrus and in the paramicrogyral zone,194,195 as well as widespread, regionally differential reduction of the GABAA receptor subunits α1, α2, α3, α5, and γ2,196 thus suggesting a disturbance in excitation-inhibition balance. Although hyperexcitability was reported in the freeze lesion model, some studies failed to demonstrate epileptogenicity in vivo.197 This illustrates one of the potential problems with a number of animal models of epileptic pathology: the dissociation between in vitro and in vivo epileptogenicity.
Genetic Models of Cortical Malformations
Telencephalic Internal Structural Heterotopia (tish) Mutant Rat
The tish mutant rat is a genetic model with a CM similar to the subcortical band heterotopia seen in humans (double cortex syndrome).198–200 These rats display a bilateral band of heterotopic neocortical neurons that extends dorsally from the frontal to the dorsoparietal neocortex.199 The bilateral heterotopia is prominent below the frontal and parietal neocortices but is rarely observed in the temporal neocortex. Heterotopic cells exhibit neocortical-like morphology, have regional connectivity characteristic of the neocortex, and are composed of cells generated during the normal period of neocortical neurogenesis.199 Double cortex syndrome in humans is linked to the X chromosome,201,202 whereas the tish mutation is not. As in patients with subcortical band heterotopia, in which seizures arise from normotopic and heterotopic areas,203 spontaneous seizures occur in most tish mutants and also appear to initiate simultaneously in normotopic and heterotopic neocortex.204 However, normotopic neurons are more prone than heterotopic neurons to exhibit epileptiform activity in the tish cortex, and heterotopic neurons are recruited into spiking by activity initiated in normotopic neurons.204 This distribution and initiation of epileptiform activity could be linked to the reported attenuation of GABAergic synaptic transmission in the tish cortex, mainly in normotopic cells, in association with a reduction in the presynaptic GABAergic terminals surrounding pyramidal cell somata in normotopic and heterotopic tish neocortex. This attenuation of inhibitory innervation was more prominent in normotopic neurons and was correlated with a reduction in a subset of GABAergic interneurons expressing parvalbumin.205 From an anatomic point of view, embryos of tish mutant rats display a second ectopic proliferative zone external to the normal periventricular proliferative zone,206 thus suggesting that band heterotopia could be a consequence of a neuronal migration disorder. Therefore, tish mutant rats seem to be a good model to study mechanisms involved in the formation of band heterotopia in humans.
The Reeler Mutant Mouse
During embryonic corticogenesis, Cajal-Retzius cells express reelin, an extracellular protein that regulates the radial migration of principal cortical neurons, in the marginal zone of the developing neocortex and hippocampus.207–209 The reeler mutant mouse has an absence of reelin caused by an autosomal recessive mutation in the reelin locus (chromosome 5 in mice).210,211 Consequently, homozygous mice exhibit severe layer disorganization in all cortical structures (e.g., neocortex, hippocampus, and cerebellum).208,212–215 In the reeler cortex, principal neurons initiate their radial migration normally but fail to assemble into layers according to an inside-out mode of development. The neocortex of the adult reeler mutant is best described as “a reversed cortex,” with the deeper layers assuming an external position.210 In the cortex, neurons of all classes survive even though they may be in abnormal positions and some may be decreased in number.216 In the hippocampus, the pyramidal cell layer is split (mainly CA1) and the DG is diffuse and disorganized. The cerebellum is hypoplastic in the reeler mouse.217,218 It has also been shown that lack of reelin signaling results in abnormal positioning and altered morphology of forebrain interneurons.219 Interestingly, a recent study demonstrated that homozygous reeler mice have enhanced seizure susceptibility that is, at least in part, intrinsic to the malformed neocortex and hippocampus.220,221 In these studies, low thresholds and an increased incidence/severity of seizures induced by MES or isoflurane were detected in vivo; in vitro, prolonged periods of spontaneous epileptiform activity in the presence of bicuculline were recorded in neocortical and hippocampal slices.221
Similar, although milder, malformations reminiscent of reeler mutant mice have been also reported in the neocortex and hippocampus of mice with a hemizygous deletion of the Lis1 gene222,223 (see the next section) and in mice lacking p35, a neuronal-specific activator of cyclin-dependent kinase (Cdk5); such mice exhibit cortical lamination defects and seizures.224,225 Mutation at the human gene encoding reelin has been identified in individuals with an autosomal recessive form of lissencephaly associated with cerebellar hypoplasia and epilepsy.226
Lis1
Classic or type I lissencephaly in humans is characterized by generalized agyria/pachygyria, four abnormal cortical layers, enlarged ventricles, generalized neuronal heterotopias, and corpus callosum defects.227,228 The loss-of-function mutation in the microtubule-associated protein–encoding gene PAFAH1B1 (encoding the Lis1 protein, or brain-specific noncatalytic subunit of platelet-activating factor acetylhydrolase 1b)229,230 was identified from patient samples with informative deletions of 17p13.3.231 Because Lis1 protein is important in the microtubule-based motor activity of cytoplasmic dynein,228,231,232 which is vital for the proper migration of neuronal precursors,233 an insufficient amount of Lis1 protein could cause migration problems with subsequent brain malformation.234,235 Mutation of the human gene, which has its homologue Lis1 in the mouse, is associated with embryonic lethality in homozygotes and disruption of neuronal migration across the cerebral parenchyma along with severe hippocampal dyslamination and hyperexcitability in heterozygotes.222,223,236 A recent set of anatomic studies showed significant disruption of the supragranular zone of the DG in adult Lis1+/− mice and aberrant neurogenesis.237 Although fiber termination patterns in the hippocampus are relatively normal, disorganization of hippocampal CA1-CA3 pyramidal cells and dispersion of granule cells are significant in the Lis1+/− mice.238 In vitro extracellular recording of hippocampal slices from Lis1+/− mice showed a reduced threshold for potassium-induced epileptiform bursting and enhanced excitatory synaptic transmission in the Lis1+/− CA1 subfield,222 which presumably contributes to the lowered threshold and intense epileptogenesis observed in the Lis1+/− mouse. Studies of embryonic slice cultures from Lis1+/− mice suggest that the nonradial migration of cortical and hippocampal interneurons is defectively slowed.234 However, an increase in large-amplitude GABA-mediated synaptic events and enhanced glutamatergic excitation of hippocampal interneurons were noted in the dysplastic CA1 pyramidal cell region of these mice.239 Further analysis of these anatomic and physiologic hippocampal defects may provide a better understanding of the neuronal basis for epileptogenesis associated with lissencephaly.
Focal Epilepsies
Aluminum Model
Focal epilepsies, or epilepsies whose seizures arise from a limited area within a hemisphere, are the most common form of epilepsy in humans. Since the 1940s, topical injection of alumina hydroxide onto the cortex of animals has been used as a model for partial seizures. Monkeys appear to be the best animal to use with this method, followed by dogs, cats, and guinea pigs. After cisternal or systemic injection of aluminum compounds, an encephalopathy with multifocal seizures can also be induced in rabbits, cats, and ferrets but not rats.240
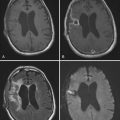
After local alumina hydroxide application onto the cortex of the temporal lobe or rolandic area, focal SRSs develop in animals in 1 to 2 months and may persist for as long as several years. Sometimes SE can occur, especially when a large injection of alumina hydroxide is applied. This can probably be attributed to subpial spread of the drug. The symptomatology depends on the location of the injection. For example, after application to the sensorimotor cortex, seizures develop that are similar to simple partial seizures in humans, with rhythmic jerking of an extremity or the face contralateral to the inflicted lesion and occasional progression to generalization. In primates, seizure symptomatology is very similar to that in humans. Scalp EEG recordings from these animals show interictal and ictal spikes, and the seizures respond to AEDs such as phenobarbital and phenytoin.241,242 The anatomic changes are characterized by cell loss, mainly GABAergic neurons (a potentially unifying theme in nearly all the animal models discussed here), and reactive gliosis at the site of injection.243 The attractive aspect of this model is that it uses animals that are phylogenetically closer to humans and induces partial seizures similar to those in humans. The disadvantages of this model include a long and unpredictable latency period before clinical and electrographic onset of spontaneous seizures (4 to 8 weeks in cats; 6 to 12 weeks in rhesus monkeys), as well as heavy resource allocation, which limits the number of animals that can be studied in any given project.5,244
Ferrous Chloride Model
The focal application of ferrous chloride as a model of epilepsy arose from the observation that deposits of iron on brain tissue after head trauma or stroke can be a risk factor for the development of epilepsy.245 In this model, ferrous chloride is injected into the cortex or amygdala of the rat. After 5 to 7 days, spontaneous seizures develop in more than 90% of the animals. Histologic analysis of the cortical lesions 6 weeks after the injection shows neuronal loss, activated astroglial cells, iron-positive macrophages, and fibroblasts surrounding the iron deposit. Some of the surviving layer V pyramidal neurons stain positive for iron, with loss of dendritic spines and decreased dendritic branching.246 The latter anatomic findings have clear implications for synaptic excitability. Interestingly, most of the AEDs available are effective in controlling the SRSs induced by ferrous chloride.247
Baraban SC, Wenzel HJ, Hochman DW, et al. Characterization of heterotopic cell clusters in the hippocampus of rats exposed to methylazoxymethanol in utero. Epilepsy Res. 2000;39:87-102.
Buckmaster PS, Dudek FE. Neuron loss, granule cell axon reorganization, and functional changes in the dentate gyrus of epileptic kainate-treated rats. J Comp Neurol. 1997;385:385-404.
Calcagnotto ME, Paredes MF, Baraban SC. Heterotopic neurons with altered inhibitory synaptic function in an animal model of malformation-associated epilepsy. J Neurosci. 2002;22:7596-7605.
Cavalheiro EA, Leite JP, Bortolotto ZA, et al. Long-term effects of pilocarpine in rats: structural damage of the brain triggers kindling and spontaneous recurrent seizures. Epilepsia. 1991;32:778-782.
Chevassus-au-Louis N, Baraban SC, Gaiarsa JL, et al. Cortical malformations and epilepsy: new insights from animal models. Epilepsia. 1999;40:811-821.
Dvorak K, Feit J, Jurankova Z. Experimentally induced focal microgyria and status verrucosus deformis in rats—pathogenesis and interrelation. Histological and autoradiographical study. Acta Neuropathol. 1978;44:121-129.
Fleck MW, Hirotsune S, Gambello MJ, et al. Hippocampal abnormalities and enhanced excitability in a murine model of human lissencephaly. J Neurosci. 2000;20:2439-2450.
Goddard GV. Development of epileptic seizures through brain stimulation at low intensity. Nature. 1967;214:1020-1021.
Houser CR. Granule cell dispersion in the dentate gyrus of humans with temporal lobe epilepsy. Brain Res. 1990;535:195-204.
Jacobs KM, Hwang BJ, Prince DA. Focal epileptogenesis in a rat model of polymicrogyria. J Neurophysiol. 1999;81:159-173.
Lee KS, Schottler F, Collins JL, et al. A genetic animal model of human neocortical heterotopia associated with seizures. J Neurosci. 1997;17:6236-6242.
Longo BM, Mello LE. Effect of long-term spontaneous recurrent seizures or reinduction of status epilepticus on the development of supragranular mossy fiber sprouting. Epilepsy Res. 1999;36:233-241.
Luhmann HJ, Karpuk N, Qu M, et al. Characterization of neuronal migration disorders in neocortical structures. II. Intracellular in vitro recordings. J Neurophysiol. 1998;80:92-102.
McIntyre DC, Kelly ME, Dufresne C. FAST and SLOW amygdala kindling rat strains: comparison of amygdala, hippocampal, piriform and perirhinal cortex kindling. Epilepsy Res. 1999;35:197-209.
Mello LE, Cavalheiro EA, Tan AM, et al. Circuit mechanisms of seizures in the pilocarpine model of chronic epilepsy: cell loss and mossy fiber sprouting. Epilepsia. 1993;34:985-995.
Nadler JV, Perry BW, Cotman CW. Intraventricular kainic acid preferentially destroys hippocampal pyramidal cells. Nature. 1978;271:676-677.
Parent JM, Yu TW, Leibowitz RT, et al. Dentate granule cell neurogenesis is increased by seizures and contributes to aberrant network reorganization in the adult rat hippocampus. J Neurosci. 1997;17:3727-3738.
Patrylo PR, Browning RA, Cranick S. Reeler homozygous mice exhibit enhanced susceptibility to epileptiform activity. Epilepsia. 2006;47:257-266.
Racine R, Rose PA, Burnham WM. Afterdischarge thresholds and kindling rates in dorsal and ventral hippocampus and dentate gyrus. Can J Neurol Sci. 1977;4:273-278.
Roper SN, Gilmore RL, Houser CR. Experimentally induced disorders of neuronal migration produce an increased propensity for electrographic seizures in rats. Epilepsy Res. 1995;21:205-219.
Sloviter RS, Kudrimoti HS, Laxer KD, et al. “Tectonic” hippocampal malformations in patients with temporal lobe epilepsy. Epilepsy Res. 2004;59:123-153.
Trotter SA, Kapur J, Anzivino MJ, et al. GABAergic synaptic inhibition is reduced before seizure onset in a genetic model of cortical malformation. J Neurosci. 2006;26:10756-10767.
Turski WA, Cavalheiro EA, Schwarz M, et al. Limbic seizures produced by pilocarpine in rats: behavioural, electroencephalographic and neuropathological study. Behav Brain Res. 1983;9:315-335.
Wang Y, Baraban SC. Granule cell dispersion and aberrant neurogenesis in the adult hippocampus of an LIS1 mutant mouse. Dev Neurosci. 2007;29:91-98.
Wenzel HJ, Robbins CA, Tsai LH, et al. Abnormal morphological and functional organization of the hippocampus in a p35 mutant model of cortical dysplasia associated with spontaneous seizures. J Neurosci. 2001;21:983-998.
Zhu WJ, Roper SN. Reduced inhibition in an animal model of cortical dysplasia. J Neurosci. 2000;20:8925-8931.
1 Begley CE, Famulari M, Annegers JF, et al. The cost of epilepsy in the United States: an estimate from population-based clinical and survey data. Epilepsia. 2000;41:342-351.
2 Engel JJ, Schwartzkroin P. What should be modeled? In: Pitkännen A, Schwartzkroin P, Moshé S, editors. Models of Seizures and Epilepsies. Philadelphia: Academic Press; 2005:1-14.
3 Engel JJ, Pedley T. What is epilepsy? In: Engel JJ, Pedley T, editors. Epilepsy: A Comprehensive Textbook, Vol 1. New York: Lippincott Williams & Wilkins; 1997:1-7.
4 Hauser WA, Annegers JF, Kurland LT. Prevalence of epilepsy in Rochester, Minnesota: 1940-1980. Epilepsia. 1991;32:429-445.
5 Fisher RS. Animal models of the epilepsies. Brain Res Brain Res Rev. 1989;14:245-278.
6 Sarkisian MR. Overview of the current animal models for human seizure and epileptic disorders. Epilepsy Behav. 2001;2:201-216.
7 Engel JJ, Williamson PD, Wieser HG. Mesial temporal lobe epilepsy. In: Engel JJ, Pedley T, editors. Epilepsy: A Comprehensive Textbook. New York: Lippincott Williams & Wilkins; 1997:2417-2426.
8 Goddard GV. Analysis of avoidance conditioning following cholinergic stimulation of amygdala in rats. J Comp Physiol Psychol. 1969;68:1-18.
9 Delgado JMR, Sevillano M. Evolution of repeated hippocampal seizures in the cat. Electroencephalogr Clin Neurophysiol. 1961;13:722-733.
10 Goddard GV. Development of epileptic seizures through brain stimulation at low intensity. Nature. 1967;214:1020-1021.
11 Racine RJ. Modification of seizure activity by electrical stimulation. II. Motor seizure. Electroencephalogr Clin Neurophysiol. 1972;32:281-294.
12 Chen L, Noffel M, Cottrell GA, et al. Amygdala-kindled convulsions in suspended rats. Exp Neurol. 1996;141:347-349.
13 Coulter DA, McIntyre DC, Loscher W. Animal models of limbic epilepsies: what can they tell us? Brain Pathol. 2002;12:240-256.
14 McIntyre DC, Kelly ME. Are differences in dorsal hippocampal kindling related to amygdala-piriform area excitability? Epilepsy Res. 1993;14:49-61.
15 McIntyre DC, Kelly ME, Armstrong JN. Kindling in the perirhinal cortex. Brain Res. 1993;615:1-6.
16 McIntyre DC, Plant JR. Long-lasting changes in the origin of spontaneous discharges from amygdala-kindled rats: piriform vs. perirhinal cortex in vitro. Brain Res. 1993;624:268-276.
17 Racine R, Rose PA, Burnham WM. Afterdischarge thresholds and kindling rates in dorsal and ventral hippocampus and dentate gyrus. Can J Neurol Sci. 1977;4:273-278.
18 Wada JA, Sato M. The generalized convulsive seizure state induced by daily electrical stimulation of the amygdala in split brain cats. Epilepsia. 1975;16:417-430.
19 Burnham WM. Primary and “transfer” seizure development in the kindled rat. Can J Neurol Sci. 1975;2:417-428.
20 McIntyre DC, Goddard GV. Transfer, interference and spontaneous recovery of convulsions kindled from the rat amygdala. Electroencephalogr Clin Neurophysiol. 1973;35:533-543.
21 Morrell F. Experimental focal epilepsy in animals. Arch Neurol. 1959;1:141-147.
22 Morrell F. Secondary epileptogenic lesions. Epilepsia. 1960;1:538-560.
23 Morrell F. Secondary epileptogenesis in man. Arch Neurol. 1985;42:318-335.
24 Applegate CD, Burchfiel JL, Konkol RJ. Kindling antagonism: effects of norepinephrine depletion on kindled seizure suppression after concurrent, alternate stimulation in rats. Exp Neurol. 1986;94:379-390.
25 Burchfiel JL, Applegate CD. Stepwise progression of kindling: perspectives from the kindling antagonism model. Neurosci Biobehav Rev. 1989;13:289-299.
26 Michalakis M, Holsinger D, Ikeda-Douglas C, et al. Development of spontaneous seizures over extended electrical kindling. I. Electrographic, behavioral, and transfer kindling correlates. Brain Res. 1998;793:197-211.
27 Pinel JP. Kindling-induced experimental epilepsy in rats: cortical stimulation. Exp Neurol. 1981;72:559-569.
28 McIntyre DC, Kelly ME, Dufresne C. FAST and SLOW amygdala kindling rat strains: comparison of amygdala, hippocampal, piriform and perirhinal cortex kindling. Epilepsy Res. 1999;35:197-209.
29 McIntyre DC, Kent P, Hayley S, et al. Influence of psychogenic and neurogenic stressors on neuroendocrine and central monoamine activity in fast and slow kindling rats. Brain Res. 1999;840:65-74.
30 Racine RJ, Steingart M, McIntyre DC. Development of kindling-prone and kindling-resistant rats: selective breeding and electrophysiological studies. Epilepsy Res. 1999;35:183-195.
31 McIntyre DC, Hutcheon B, Schwabe K, et al. Divergent GABA(A) receptor–mediated synaptic transmission in genetically seizure-prone and seizure-resistant rats. J Neurosci. 2002;22:9922-9931.
32 McIntyre DC, Poulter MO, Gilby K. Kindling: some old and some new. Epilepsy Res. 2002;50:79-92.
33 Poulter MO, Brown LA, Tynan S, et al. Differential expression of alpha1, alpha2, alpha3, and alpha5 GABAA receptor subunits in seizure-prone and seizure-resistant rat models of temporal lobe epilepsy. J Neurosci. 1999;19:4654-4661.
34 Moshe SL. The effects of age on the kindling phenomenon. Dev Psychobiol. 1981;14:75-81.
35 Thompson JL, Holmes GL, Feldman DS. Transfer following rapid kindling in the prepubescent rat. Epilepsy Res. 1989;3:222-226.
36 Gotman J. Relationships between triggered seizures, spontaneous seizures, and interictal spiking in the kindling model of epilepsy. Exp Neurol. 1984;84:259-273.
37 Wada JA, Osawa T. Spontaneous recurrent seizure state induced by daily electric amygdaloid stimulation in Senegalese baboons (Papio papio). Neurology. 1976;26:273-286.
38 Wauquier A, Ashton D, Melis W. Behavioral analysis of amygdaloid kindling in beagle dogs and the effects of clonazepam, diazepam, phenobarbital, diphenylhydantoin, and flunarizine on seizure manifestation. Exp Neurol. 1979;64:579-586.
39 McIntyre D. The kindling phenomenon. In: Pitkännen A, Schwartzkroin P, Moshé S, editors. Models of Seizures and Epilepsies. Philadelphia: Academic Press; 2005:351-363.
40 Otsuki K, Morimoto K, Sato K, et al. Effects of lamotrigine and conventional antiepileptic drugs on amygdala- and hippocampal-kindled seizures in rats. Epilepsy Res. 1998;31:101-112.
41 Loscher W, Jackel R, Czuczwar SJ. Is amygdala kindling in rats a model for drug-resistant partial epilepsy? Exp Neurol. 1986;93:211-226.
42 De Smedt T, Raedt R, Vonck K, et al. Levetiracetam: the profile of a novel anticonvulsant drug—part I: preclinical data. CNS Drug Rev. 2007;13:43-56.
43 Glien M, Brandt C, Potschka H, et al. Effects of the novel antiepileptic drug levetiracetam on spontaneous recurrent seizures in the rat pilocarpine model of temporal lobe epilepsy. Epilepsia. 2002;43:350-357.
44 Klitgaard H, Matagne A, Gobert J, et al. Evidence for a unique profile of levetiracetam in rodent models of seizures and epilepsy. Eur J Pharmacol. 1998;353:191-206.
45 Loscher W, Honack D, Rundfeldt C. Antiepileptogenic effects of the novel anticonvulsant levetiracetam (ucb L059) in the kindling model of temporal lobe epilepsy. J Pharmacol Exp Ther. 1998;284:474-479.
46 Loscher W. Animal models of intractable epilepsy. Prog Neurobiol. 1997;53:239-258.
47 Rundfeldt C, Honack D, Loscher W. Phenytoin potently increases the threshold for focal seizures in amygdala-kindled rats. Neuropharmacology. 1990;29:845-851.
48 Loscher W. Animal models of epilepsy for the development of antiepileptogenic and disease-modifying drugs. A comparison of the pharmacology of kindling and post–status epilepticus models of temporal lobe epilepsy. Epilepsy Res. 2002;50:105-123.
49 Cavazos JE, Das I, Sutula TP. Neuronal loss induced in limbic pathways by kindling: evidence for induction of hippocampal sclerosis by repeated brief seizures. J Neurosci. 1994;14:3106-3121.
50 Cavazos JE, Sutula TP. Progressive neuronal loss induced by kindling: a possible mechanism for mossy fiber synaptic reorganization and hippocampal sclerosis. Brain Res. 1990;527:1-6.
51 Kotloski R, Lynch M, Lauersdorf S, et al. Repeated brief seizures induce progressive hippocampal neuron loss and memory deficits. Prog Brain Res. 2002;135:95-110.
52 Parent JM, Janumpalli S, McNamara JO, et al. Increased dentate granule cell neurogenesis following amygdala kindling in the adult rat. Neurosci Lett. 1998;247:9-12.
53 Sutula TP. A glimpse into abnormal cortical development and epileptogenesis at epilepsy surgery. Neurology. 1998;50:8-10.
54 Represa A, Ben-Ari Y. Kindling is associated with the formation of novel mossy fibre synapses in the CA3 region. Exp Brain Res. 1992;92:69-78.
55 Khurgel M, Ivy GO. Astrocytes in kindling: relevance to epileptogenesis. Epilepsy Res. 1996;26:163-175.
56 Sutula TP. Mechanisms of epilepsy progression: current theories and perspectives from neuroplasticity in adulthood and development. Epilepsy Res. 2004;60:161-171.
57 Sayin, Rutecki P, Sutula T. NMDA-dependent currents in granule cells of the dentate gyrus contribute to induction but not permanence of kindling. J Neurophysiol. 1999;81:564-574.
58 Sutula T, Koch J, Golarai G, et al. NMDA receptor dependence of kindling and mossy fiber sprouting: evidence that the NMDA receptor regulates patterning of hippocampal circuits in the adult brain. J Neurosci. 1996;16:7398-7406.
59 Loscher W, Honack D, Gramer M. Effect of depth electrode implantation with or without subsequent kindling on GABA turnover in various rat brain regions. Epilepsy Res. 1999;37:95-108.
60 Loscher W, Schirmer M, Freichel C, et al. Distribution of GABAergic neurons in the striatum of amygdala-kindled rats: an immunohistochemical and in situ hybridization study. Brain Res. 2006;1083:50-60.
61 Loscher W, Schwark WS. Evidence for impaired GABAergic activity in the substantia nigra of amygdaloid kindled rats. Brain Res. 1985;339:146-150.
62 Loscher W, Schwark WS. Further evidence for abnormal GABAergic circuits in amygdala-kindled rats. Brain Res. 1987;420:385-390.
63 Gavrilovici C, D’Alfonso S, Dann M, et al. Kindling-induced alterations in GABAA receptor–mediated inhibition and neurosteroid activity in the rat piriform cortex. Eur J Neurosci. 2006;24:1373-1384.
64 Ben-Ari Y. Limbic seizure and brain damage produced by kainic acid: mechanisms and relevance to human temporal lobe epilepsy. Neuroscience. 1985;14:375-403.
65 Buckmaster PS, Dudek FE. Neuron loss, granule cell axon reorganization, and functional changes in the dentate gyrus of epileptic kainate-treated rats. J Comp Neurol. 1997;385:385-404.
66 Turski L, Ikonomidou C, Turski WA, et al. Review: cholinergic mechanisms and epileptogenesis. The seizures induced by pilocarpine: a novel experimental model of intractable epilepsy. Synapse. 1989;3:154-171.
67 Turski WA, Cavalheiro EA, Schwarz M, et al. Limbic seizures produced by pilocarpine in rats: behavioural, electroencephalographic and neuropathological study. Behav Brain Res. 1983;9:315-335.
68 Ben-Ari Y, Lagowska J. [Epileptogenic action of intra-amygdaloid injection of kainic acid.]. C R Acad Sci Hebd Seances Acad Sci D. 1978;287:813-816.
69 Nadler JV, Perry BW, Cotman CW. Intraventricular kainic acid preferentially destroys hippocampal pyramidal cells. Nature. 1978;271:676-677.
70 Cavalheiro EA, Riche DA, Le Gal La Salle G. Long-term effects of intrahippocampal kainic acid injection in rats: a method for inducing spontaneous recurrent seizures. Electroencephalogr Clin Neurophysiol. 1982;53:581-589.
71 Leite JP, Bortolotto ZA, Cavalheiro EA. Spontaneous recurrent seizures in rats: an experimental model of partial epilepsy. Neurosci Biobehav Rev. 1990;14:511-517.
72 Lemos T, Cavalheiro EA. Suppression of pilocarpine-induced status epilepticus and the late development of epilepsy in rats. Exp Brain Res. 1995;102:423-428.
73 Mello LE, Cavalheiro EA, Tan AM, et al. Circuit mechanisms of seizures in the pilocarpine model of chronic epilepsy: cell loss and mossy fiber sprouting. Epilepsia. 1993;34:985-995.
74 Glien M, Brandt C, Potschka H, et al. Repeated low-dose treatment of rats with pilocarpine: low mortality but high proportion of rats developing epilepsy. Epilepsy Res. 2001;46:111-119.
75 Hellier JL, Patrylo PR, Buckmaster PS, et al. Recurrent spontaneous motor seizures after repeated low-dose systemic treatment with kainate: assessment of a rat model of temporal lobe epilepsy. Epilepsy Res. 1998;31:73-84.
76 Leite JP, Garcia-Cairasco N, Cavalheiro EA. New insights from the use of pilocarpine and kainate models. Epilepsy Res. 2002;50:93-103.
77 Cavalheiro EA. The pilocarpine model of epilepsy. Ital J Neurol Sci. 1995;16:33-37.
78 Scerrati M, Onofrj M, Pacifici L, et al. Electrocerebral and behavioural analysis of systemic kainic acid–induced epilepsy in the rat. Drugs Exp Clin Res. 1986;12:671-680.
79 Cavalheiro EA, Leite JP, Bortolotto ZA, et al. Long-term effects of pilocarpine in rats: structural damage of the brain triggers kindling and spontaneous recurrent seizures. Epilepsia. 1991;32:778-782.
80 Williams PA, Hellier JL, White AM, et al. Development of spontaneous seizures after experimental status epilepticus: implications for understanding epileptogenesis. Epilepsia. 2007;48(suppl 5):157-163.
81 Cronin J, Dudek FE. Chronic seizures and collateral sprouting of dentate mossy fibers after kainic acid treatment in rats. Brain Res. 1988;474:181-184.
82 Pisa M, Sanberg PR, Corcoran ME, et al. Spontaneously recurrent seizures after intracerebral injections of kainic acid in rat: a possible model of human temporal lobe epilepsy. Brain Res. 1980;200:481-487.
83 Santos NF, Marques RH, Correia L, et al. Multiple pilocarpine-induced status epilepticus in developing rats: a long-term behavioral and electrophysiological study. Epilepsia. 2000;41(suppl 6):S57-S63.
84 Turski WA, Cavalheiro EA, Bortolotto ZA, et al. Seizures produced by pilocarpine in mice: a behavioral, electroencephalographic and morphological analysis. Brain Res. 1984;321:237-253.
85 Bolanos AR, Sarkisian M, Yang Y, et al. Comparison of valproate and phenobarbital treatment after status epilepticus in rats. Neurology. 1998;51:41-48.
86 Leite JP, Cavalheiro EA. Effects of conventional antiepileptic drugs in a model of spontaneous recurrent seizures in rats. Epilepsy Res. 1995;20:93-104.
87 Muller-Schwarze AB, Tandon P, Liu Z, et al. Ketogenic diet reduces spontaneous seizures and mossy fiber sprouting in the kainic acid model. Neuroreport. 1999;10:1517-1522.
88 Babb TL, Lieb JP, Brown WJ, et al. Distribution of pyramidal cell density and hyperexcitability in the epileptic human hippocampal formation. Epilepsia. 1984;25:721-728.
89 Engel JJr. Recent advances in surgical treatment of temporal lobe epilepsy. Acta Neurol Scand Suppl. 1992;140:71-80.
90 Margerison JH, Corsellis JA. Epilepsy and the temporal lobes. A clinical, electroencephalographic and neuropathological study of the brain in epilepsy, with particular reference to the temporal lobes. Brain. 1966;89:499-530.
91 Houser CR. Granule cell dispersion in the dentate gyrus of humans with temporal lobe epilepsy. Brain Res. 1990;535:195-204.
92 Sutula T, Cascino G, Cavazos J, et al. Mossy fiber synaptic reorganization in the epileptic human temporal lobe. Ann Neurol. 1989;26:321-330.
93 Nadler JV. Minireview. Kainic acid as a tool for the study of temporal lobe epilepsy. Life Sci. 1981;29:2031-2042.
94 Nitecka L, Tremblay E, Charton G, et al. Maturation of kainic acid seizure–brain damage syndrome in the rat. II. Histopathological sequelae. Neuroscience. 1984;13:1073-1094.
95 Tremblay E, Nitecka L, Berger ML, et al. Maturation of kainic acid seizure–brain damage syndrome in the rat. I. Clinical, electrographic and metabolic observations. Neuroscience. 1984;13:1051-1072.
96 Clifford DB, Olney JW, Maniotis A, et al. The functional anatomy and pathology of lithium-pilocarpine and high-dose pilocarpine seizures. Neuroscience. 1987;23:953-968.
97 Olney JW, Collins RC, Sloviter RS. Excitotoxic mechanisms of epileptic brain damage. Adv Neurol. 1986;44:857-877.
98 Turski L, Cavalheiro EA, Sieklucka-Dziuba M, et al. Seizures produced by pilocarpine: neuropathological sequelae and activity of glutamate decarboxylase in the rat forebrain. Brain Res. 1986;398:37-48.
99 Hamani C, Mello LE. Status epilepticus induced by pilocarpine and picrotoxin. Epilepsy Res. 1997;28:73-82.
100 Obenaus A, Esclapez M, Houser CR. Loss of glutamate decarboxylase mRNA–containing neurons in the rat dentate gyrus following pilocarpine-induced seizures. J Neurosci. 1993;13:4470-4485.
101 Mello LE, Cavalheiro EA, Tan AM, et al. Granule cell dispersion in relation to mossy fiber sprouting, hippocampal cell loss, silent period and seizure frequency in the pilocarpine model of epilepsy. Epilepsy Res Suppl. 1992;9:51-59.
102 Parent JM, Yu TW, Leibowitz RT, et al. Dentate granule cell neurogenesis is increased by seizures and contributes to aberrant network reorganization in the adult rat hippocampus. J Neurosci. 1997;17:3727-3738.
103 Buckmaster PS, Zhang GF, Yamawaki R. Axon sprouting in a model of temporal lobe epilepsy creates a predominantly excitatory feedback circuit. J Neurosci. 2002;22:6650-6658.
104 Okazaki MM, Evenson DA, Nadler JV. Hippocampal mossy fiber sprouting and synapse formation after status epilepticus in rats: visualization after retrograde transport of biocytin. J Comp Neurol. 1995;352:515-534.
105 Longo BM, Mello LE. Blockade of pilocarpine- or kainate-induced mossy fiber sprouting by cycloheximide does not prevent subsequent epileptogenesis in rats. Neurosci Lett. 1997;226:163-166.
106 Longo BM, Mello LE. Effect of long-term spontaneous recurrent seizures or reinduction of status epilepticus on the development of supragranular mossy fiber sprouting. Epilepsy Res. 1999;36:233-241.
107 Perez Y, Morin F, Beaulieu C, et al. Axonal sprouting of CA1 pyramidal cells in hyperexcitable hippocampal slices of kainate-treated rats. Eur J Neurosci. 1996;8:736-748.
108 Kobayashi M, Buckmaster PS. Reduced inhibition of dentate granule cells in a model of temporal lobe epilepsy. J Neurosci. 2003;23:2440-2452.
109 Kumar SS, Buckmaster PS. Hyperexcitability, interneurons, and loss of GABAergic synapses in entorhinal cortex in a model of temporal lobe epilepsy. J Neurosci. 2006;26:4613-4623.
110 Brooks-Kayal AR, Shumate MD, Jin H, et al. Selective changes in single cell GABA(A) receptor subunit expression and function in temporal lobe epilepsy. Nat Med. 1998;4:1166-1172.
111 Fritschy JM, Kiener T, Bouilleret V, et al. GABAergic neurons and GABA(A)-receptors in temporal lobe epilepsy. Neurochem Int. 1999;34:435-445.
112 Houser CR, Esclapez M. Downregulation of the alpha5 subunit of the GABA(A) receptor in the pilocarpine model of temporal lobe epilepsy. Hippocampus. 2003;13:633-645.
113 Peng Z, Hauer B, Mihalek RM, et al. GABA(A) receptor changes in delta subunit–deficient mice: altered expression of alpha4 and gamma2 subunits in the forebrain. J Comp Neurol. 2002;446:179-197.
114 Rice A, Rafiq A, Shapiro SM, et al. Long-lasting reduction of inhibitory function and gamma-aminobutyric acid type A receptor subunit mRNA expression in a model of temporal lobe epilepsy. Proc Natl Acad Sci U S A. 1996;93:9665-9669.
115 Schwarzer C, Tsunashima K, Wanzenbock C, et al. GABA(A) receptor subunits in the rat hippocampus II: altered distribution in kainic acid–induced temporal lobe epilepsy. Neuroscience. 1997;80:1001-1017.
116 Tsunashima K, Schwarzer C, Kirchmair E, et al. GABA(A) receptor subunits in the rat hippocampus III: altered messenger RNA expression in kainic acid–induced epilepsy. Neuroscience. 1997;80:1019-1032.
117 Lurton D, Cavalheiro EA. Neuropeptide-Y immunoreactivity in the pilocarpine model of temporal lobe epilepsy. Exp Brain Res. 1997;116:186-190.
118 Cavalheiro EA, Fernandes MJ, Turski L, et al. Spontaneous recurrent seizures in rats: amino acid and monoamine determination in the hippocampus. Epilepsia. 1994;35:1-11.
119 Costa MS, Rocha JB, Perosa SR, et al. Pilocarpine-induced status epilepticus increases glutamate release in rat hippocampal synaptosomes. Neurosci Lett. 2004;356:41-44.
120 Ormandy GC, Jope RS, Snead OC3rd. Anticonvulsant actions of MK-801 on the lithium-pilocarpine model of status epilepticus in rats. Exp Neurol. 1989;106:172-180.
121 Scorza CA, Garrido Ydel C, Arida RM, et al. Levels of the synaptic protein X11 alpha/mint1 are increased in hippocampus of rats with epilepsy. Epilepsy Res. 2003;57:49-57.
122 Tang FR, Chia SC, Chen PM, et al. Metabotropic glutamate receptor 2/3 in the hippocampus of patients with mesial temporal lobe epilepsy, and of rats and mice after pilocarpine-induced status epilepticus. Epilepsy Res. 2004;59:167-180.
123 Sloviter RS, Kudrimoti HS, Laxer KD, et al. “Tectonic” hippocampal malformations in patients with temporal lobe epilepsy. Epilepsy Res. 2004;59:123-153.
124 Perez-Mendes P. Development of a New Model of Temporal Lobe Epilepsy in Marmosets. [PhD thesis]. São Paulo: Physiology, UNIFESP. 2007.
125 Perez-Mendes P, Blanco MM, Calcagnotto ME, et al. Development of a new model of temporal lobe epilepsy in marmosets (submitted).
126 Eriksson PS, Perfilieva E, Bjork-Eriksson T, et al. Neurogenesis in the adult human hippocampus. Nat Med. 1998;4:1313-1317.
127 Palmini A, Andermann F, Olivier A, et al. Focal neuronal migration disorders and intractable partial epilepsy: a study of 30 patients. Ann Neurol. 1991;30:741-749.
128 Palmini A, Andermann F, Olivier A, et al. Neuronal migration disorders: a contribution of modern neuroimaging to the etiologic diagnosis of epilepsy. Can J Neurol Sci. 1991;18(4 suppl):580-587.
129 Robain O. Introduction in the pathology of cerebral cortical dysplasia. In: Guerrini R, Andermann F, Canapicchi R, et al, editors. Dysplasias of Cerebral Cortex and Epilepsy. Philadelphia: Lippincott-Raven; 1996:1-10.
130 Taylor DC, Falconer MA, Bruton CJ, et al. Focal dysplasia of the cerebral cortex in epilepsy. J Neurol Neurosurg Psychiatry. 1971;34:369-387.
131 Calcagnotto ME, Paredes MF, Baraban SC. Heterotopic neurons with altered inhibitory synaptic function in an animal model of malformation-associated epilepsy. J Neurosci. 2002;22:7596-7605.
132 Calcagnotto ME, Paredes MF, Tihan T, et al. Dysfunction of synaptic inhibition in epilepsy associated with focal cortical dysplasia. J Neurosci. 2005;25:9649-9657.
133 Chen HX, Roper SN. Reduction of spontaneous inhibitory synaptic activity in experimental heterotopic gray matter. J Neurophysiol. 2003;89:150-158.
134 Zhu WJ, Roper SN. Reduced inhibition in an animal model of cortical dysplasia. J Neurosci. 2000;20:8925-8931.
135 Cattaneo E, Reinach B, Caputi A, et al. Selective in vitro blockade of neuroepithelial cells proliferation by methylazoxymethanol, a molecule capable of inducing long lasting functional impairments. J Neurosci Res. 1995;41:640-647.
136 Hicks SP, D’Amato CJ, Lowe MJ. The development of the mammalian nervous system: I. Malformation of the brain, especially the cerebral cortex, induced in rats by radiation. II. Some mechanisms of the malformations of the cortex. J Comp Neurol. 1959;113:435-469.
137 Dvorak K, Feit J. Migration of neuroblasts through partial necrosis of the cerebral cortex in newborn rats—contribution to the problems of morphological development and developmental period of cerebral microgyria. Histological and autoradiographical study. Acta Neuropathol. 1977;38:203-212.
138 Dvorak K, Feit J, Jurankova Z. Experimentally induced focal microgyria and status verrucosus deformis in rats—pathogenesis and interrelation. Histological and autoradiographical study. Acta Neuropathol. 1978;44:121-129.
139 Bayer SA, Altman J, Russo RJ, et al. Timetables of neurogenesis in the human brain based on experimentally determined patterns in the rat. Neurotoxicology. 1993;14:83-144.
140 Kimler BF. Prenatal irradiation: a major concern for the developing brain. Int J Radiat Biol. 1998;73:423-434.
141 Haddad RK, Rabe A, Laqueur GL, et al. Intellectual deficit associated with transplacentally induced microcephaly in the rat. Science. 1969;163:88-90.
142 Laqueur GL, Spatz M. Toxicology of cycasin. Cancer Res. 1968;28:2262-2267.
143 Nagata Y, Matsumoto H. Studies on methylazoxymethanol: methylation of nucleic acids in the fetal rat brain. Proc Soc Exp Biol Med. 1969;132:383-385.
144 Spatz M, Dougherty WJ, Smith DW. Teratogenic effects of methylazoxymethanol. Proc Soc Exp Biol Med. 1967;124:476-478.
145 Spatz M, Laqueur GL. Transplacental chemical induction of microencephaly in two strains of rats. I. Proc Soc Exp Biol Med. 1968;129:705-710.
146 Johnston MV, Coyle JT. Histological and neurochemical effects of fetal treatment with methylazoxymethanol on rat neocortex in adulthood. Brain Res. 1979;170(1):135-155.
147 Baraban SC, Wenzel HJ, Hochman DW, Schwartzkroin PA. Characterization of heterotopic cell clusters in the hippocampus of rats exposed to methylazoxymethanol in utero. Epilepsy Res. 2000;39:87-102.
148 Colacitti C, Sancini G, DeBiasi S, et al. Prenatal methylazoxymethanol treatment in rats produces brain abnormalities with morphological similarities to human developmental brain dysgeneses. J Neuropathol Exp Neurol. 1999;58:92-106.
149 Dambska M, Haddad R, Kozlowski PB, et al. Telencephalic cytoarchitectonics in the brains of rats with graded degrees of micrencephaly. Acta Neuropathol. 1982;58:203-209.
150 Germano IM, Sperber EF. Transplacentally induced neuronal migration disorders: an animal model for the study of the epilepsies. J Neurosci Res. 1998;51:473-488.
151 Singh SC. Ectopic neurones in the hippocampus of the postnatal rat exposed to methylazoxymethanol during foetal development. Acta Neuropathol (Berl). 1977;40:111-116.
152 Yurkewicz L, Valentino KL, Floeter MK, et al. Effects of cytotoxic deletions of somatic sensory cortex in fetal rats. Somatosens Res. 1984;1:303-327.
153 Zhang LL, Collier PA, Ashwell KW. Mechanisms in the induction of neuronal heterotopia following prenatal cytotoxic brain damage. Neurotoxicol Teratol. 1995;17:297-311.
154 Paredes M, Pleasure SJ, Baraban SC. Embryonic and early postnatal abnormalities contributing to the development of hippocampal malformations in a rodent model of dysplasia. J Comp Neurol. 2006;495:133-148.
155 Emery JA, Roper SN, Rojiani AM. White matter neuronal heterotopia in temporal lobe epilepsy: a morphometric and immunohistochemical study. J Neuropathol Exp Neurol. 1997;56:1276-1282.
156 Chevassus-Au-Louis N, Congar P, Represa A, et al. Neuronal migration disorders: heterotopic neocortical neurons in CA1 provide a bridge between the hippocampus and the neocortex. Proc Natl Acad Sci U S A. 1998;95:10263-10268.
157 Castro PA, Pleasure SJ, Baraban SC. Hippocampal heterotopia with molecular and electrophysiological properties of neocortical neurons. Neuroscience. 2002;114:961-972.
158 Baraban SC, Schwartzkroin PA. Flurothyl seizure susceptibility in rats following prenatal methylazoxymethanol treatment. Epilepsy Res. 1996;23:189-194.
159 Chevassus-au-Louis N, Ben-Ari Y, Vergnes M. Decreased seizure threshold and more rapid rate of kindling in rats with cortical malformation induced by prenatal treatment with methylazoxymethanol. Brain Res. 1998;812:252-255.
160 Germano IM, Zhang YF, Sperber EF, et al. Neuronal migration disorders increase susceptibility to hyperthermia-induced seizures in developing rats. Epilepsia. 1996;37:902-910.
161 Smyth MD, Barbaro NM, Baraban SC. Effects of antiepileptic drugs on induced epileptiform activity in a rat model of dysplasia. Epilepsy Res. 2002;50:251-264.
162 Castro PA, Cooper EC, Lowenstein DH, et al. Hippocampal heterotopia lack functional Kv4.2 potassium channels in the methylazoxymethanol model of cortical malformations and epilepsy. J Neurosci. 2001;21:6626-6634.
163 Sancini G, Franceschetti S, Battaglia G, et al. Dysplastic neocortex and subcortical heterotopias in methylazoxymethanol-treated rats: an intracellular study of identified pyramidal neurones. Neurosci Lett. 1998;246:181-185.
164 Calcagnotto ME, Baraban SC. Prolonged NMDA-mediated responses, altered ifenprodil sensitivity, and epileptiform-like events in the malformed hippocampus of methylazoxymethanol exposed rats. J Neurophysiol. 2005;94:153-162.
165 Harrington EP, Moddel G, Najm IM, et al. Altered glutamate receptor-transporter expression and spontaneous seizures in rats exposed to methylazoxymethanol in utero. Epilepsia. 2007;48:158-168.
166 Pentney AR, Baraban SC, Colmers WF. NPY sensitivity and postsynaptic properties of heterotopic neurons in the MAM model of malformation-associated epilepsy. J Neurophysiol. 2002;88:2745-2754.
167 Roper SN, Abraham LA, Streit WJ. Exposure to in utero irradiation produces disruption of radial glia in rats. Dev Neurosci. 1997;19:521-528.
168 Ferrer I, Alcantara S, Catala I, et al. Experimentally induced laminar necrosis, status verrucosus, focal cortical dysplasia reminiscent of microgyria, and porencephaly in the rat. Exp Brain Res. 1993;94:261-269.
169 Roper SN. In utero irradiation of rats as a model of human cerebrocortical dysgenesis: a review. Epilepsy Res. 1998;32:63-74.
170 Cowen D, Geller LM. Long-term pathological effects of prenatal x-irradiation on the central nervous system of the rat. J Neuropathol Exp Neurol. 1960;19:488-527.
171 McGrath JJ, Riggs HE, Schwarz HP. Malformation of the adult brain (albino rat) resulting from prenatal irradiation. J Neuropathol Exp Neurol. 1956;15:432-446.
172 Roper SN, Gilmore RL, Houser CR. Experimentally induced disorders of neuronal migration produce an increased propensity for electrographic seizures in rats. Epilepsy Res. 1995;21:205-219.
173 Kellinghaus C, Kunieda T, Ying Z, et al. Severity of histopathologic abnormalities and in vivo epileptogenicity in the in utero radiation model of rats is dose dependent. Epilepsia. 2004;45:583-591.
174 Kondo S, Najm I, Kunieda T, et al. Electroencephalographic characterization of an adult rat model of radiation-induced cortical dysplasia. Epilepsia. 2001;42:1221-1227.
175 Fushiki S, Matsushita K, Yoshioka H, et al. In utero exposure to low-doses of ionizing radiation decelerates neuronal migration in the developing rat brain. Int J Radiat Biol. 1996;70:53-60.
176 Miki T, Sawada K, Sun XZ, et al. Abnormal distribution of hippocampal mossy fibers in rats exposed to X-irradiation in utero. Brain Res Dev Brain Res. 1999;112:275-280.
177 Roper SN, King MA, Abraham LA, et al. Disinhibited in vitro neocortical slices containing experimentally induced cortical dysplasia demonstrate hyperexcitability. Epilepsy Res. 1997;26:443-449.
178 Roper SN, Eisenschenk S, King MA. Reduced density of parvalbumin- and calbindin D28–immunoreactive neurons in experimental cortical dysplasia. Epilepsy Res. 1999;37:63-71.
179 Marret S, Mukendi R, Gadisseux JF, et al. Effect of ibotenate on brain development: an excitotoxic mouse model of microgyria and posthypoxic-like lesions. J Neuropathol Exp Neurol. 1995;54:358-370.
180 Jacobs KM, Gutnick MJ, Prince DA. Hyperexcitability in a model of cortical maldevelopment. Cereb Cortex. 1996;6:514-523.
181 Levine DN, Fisher MA, Caviness VSJr. Porencephaly with microgyria: a pathologic study. Acta Neuropathol. 1974;29:99-113.
182 Williams RS, Ferrante RJ, Caviness VSJr. The cellular pathology of microgyria. A Golgi analysis. Acta Neuropathol. 1976;36:269-283.
183 McBride MC, Kemper TL. Pathogenesis of four-layered microgyric cortex in man. Acta Neuropathol. 1982;57:93-98.
184 Prince DA, Jacobs KM, Salin PA, et al. Chronic focal neocortical epileptogenesis: does disinhibition play a role? Can J Physiol Pharmacol. 1997;75:500-507.
185 Richman DP, Stewart RM, Caviness VSJr. Cerebral microgyria in a 27-week fetus: an architectonic and topographic analysis. J Neuropathol Exp Neurol. 1974;33:374-384.
186 Barkovich AJ, Rowley H, Bollen A. Correlation of prenatal events with the development of polymicrogyria. AJNR Am J Neuroradiol. 1995;16(4 suppl):822-827.
187 Cohen M, Roessmann U. In utero brain damage: relationship of gestational age to pathological consequences. Dev Med Child Neurol. 1994;36:263-268.
188 DeFazio RA, Hablitz JJ. Alterations in NMDA receptors in a rat model of cortical dysplasia. J Neurophysiol. 2000;83:315-321.
189 Hablitz JJ, DeFazio RA. Altered receptor subunit expression in rat neocortical malformations. Epilepsia. 2000;41(suppl 6):S82-S85.
190 Jacobs KM, Hwang BJ, Prince DA. Focal epileptogenesis in a rat model of polymicrogyria. J Neurophysiol. 1999;81:159-173.
191 Luhmann HJ, Karpuk N, Qu M, et al. Characterization of neuronal migration disorders in neocortical structures. II. Intracellular in vitro recordings. J Neurophysiol. 1998;80:92-102.
192 Humphreys P, Rosen GD, Press DM, et al. Freezing lesions of the developing rat brain: a model for cerebrocortical microgyria. J Neuropathol Exp Neurol. 1991;50:145-160.
193 Rosen GD, Burstein D, Galaburda AM. Changes in efferent and afferent connectivity in rats with induced cerebrocortical microgyria. J Comp Neurol. 2000;418:423-440.
194 Zilles K, Qu M, Schleicher A, et al. Characterization of neuronal migration disorders in neocortical structures: quantitative receptor autoradiography of ionotropic glutamate, GABA(A) and GABA(B) receptors. Eur J Neurosci. 1998;10:3095-3106.
195 Rosen GD, Jacobs KM, Prince DA. Effects of neonatal freeze lesions on expression of parvalbumin in rat neocortex. Cereb Cortex. 1998;8:753-761.
196 Redecker C, Luhmann HJ, Hagemann G, et al. Differential downregulation of GABAA receptor subunits in widespread brain regions in the freeze-lesion model of focal cortical malformations. J Neurosci. 2000;20:5045-5053.
197 Holmes GL, Sarkisian M, Ben-Ari Y, et al. Consequences of cortical dysplasia during development in rats. Epilepsia. 1999;40:537-544.
198 Chevassus-au-Louis N, Baraban SC, Gaiarsa JL, et al. Cortical malformations and epilepsy: new insights from animal models. Epilepsia. 1999;40:811-821.
199 Lee KS, Schottler F, Collins JL, et al. A genetic animal model of human neocortical heterotopia associated with seizures. J Neurosci. 1997;17:6236-6242.
200 Prasad AN, Prasad C, Stafstrom CE. Recent advances in the genetics of epilepsy: insights from human and animal studies. Epilepsia. 1999;40:1329-1352.
201 des Portes V, Pinard JM, Billuart P, et al. A novel CNS gene required for neuronal migration and involved in X-linked subcortical laminar heterotopia and lissencephaly syndrome. Cell. 1998;92:51-61.
202 Gleeson JG, Allen KM, Fox JW, et al. Doublecortin, a brain-specific gene mutated in human X-linked lissencephaly and double cortex syndrome, encodes a putative signaling protein. Cell. 1998;92:63-72.
203 Bernasconi A, Martinez V, Rosa-Neto P, et al. Surgical resection for intractable epilepsy in “double cortex” syndrome yields inadequate results. Epilepsia. 2001;42:1124-1129.
204 Chen ZF, Schottler F, Bertram E, et al. Distribution and initiation of seizure activity in a rat brain with subcortical band heterotopia. Epilepsia. 2000;41:493-501.
205 Trotter SA, Kapur J, Anzivino MJ, et al. GABAergic synaptic inhibition is reduced before seizure onset in a genetic model of cortical malformation. J Neurosci. 2006;26:10756-10767.
206 Lee KS, Collins JL, Anzivino MJ, et al. Heterotopic neurogenesis in a rat with cortical heterotopia. J Neurosci. 1998;18:9365-9375.
207 Alcantara S, Ruiz M, D’Arcangelo G, et al. Regional and cellular patterns of reelin mRNA expression in the forebrain of the developing and adult mouse. J Neurosci. 1998;18:7779-7799.
208 D’Arcangelo G, Miao GG, Chen SC, et al. A protein related to extracellular matrix proteins deleted in the mouse mutant reeler. Nature. 1995;374:719-723.
209 Ogawa M, Miyata T, Nakajima K, et al. The reeler gene–associated antigen on Cajal-Retzius neurons is a crucial molecule for laminar organization of cortical neurons. Neuron. 1995;14:899-912.
210 Caviness VSJr, Sidman RL. Olfactory structures of the forebrain in the reeler mutant mouse. J Comp Neurol. 1972;145:85-104.
211 Rakic P, Caviness VSJr. Cortical development: view from neurological mutants two decades later. Neuron. 1995;14:1101-1104.
212 Caviness VSJr. Patterns of cell and fiber distribution in the neocortex of the reeler mutant mouse. J Comp Neurol. 1976;170:435-447.
213 D’Arcangelo G, Miao GG, Curran T. Detection of the reelin breakpoint in reeler mice. Brain Res Mol Brain Res. 1996;39:234-236.
214 Lambert de Rouvroit C, Goffinet AM. The reeler mouse as a model of brain development. Adv Anat Embryol Cell Biol. 1998;150:1-106.
215 Rice DS, Curran T. Role of the reelin signaling pathway in central nervous system development. Annu Rev Neurosci. 2001;24:1005-1039.
216 Caviness VSJr, Rakic P. Mechanisms of cortical development: a view from mutations in mice. Annu Rev Neurosci. 1978;1:297-326.
217 Drakew A, Deller T, Heimrich B, et al. Dentate granule cells in reeler mutants and VLDLR and ApoER2 knockout mice. Exp Neurol. 2002;176:12-24.
218 Stanfield BB, Cowan WM. The morphology of the hippocampus and dentate gyrus in normal and reeler mice. J Comp Neurol. 1979;185:393-422.
219 Yabut O, Renfro A, Niu S, et al. Abnormal laminar position and dendrite development of interneurons in the reeler forebrain. Brain Res. 2007;1140:75-83.
220 Kopjas NN, Jones RT, Bany B, et al. Reeler mutant mice exhibit seizures during recovery from isoflurane-induced anesthesia. Epilepsy Res. 2006;69:87-91.
221 Patrylo PR, Browning RA, Cranick S. Reeler homozygous mice exhibit enhanced susceptibility to epileptiform activity. Epilepsia. 2006;47:257-266.
222 Fleck MW, Hirotsune S, Gambello MJ, et al. Hippocampal abnormalities and enhanced excitability in a murine model of human lissencephaly. J Neurosci. 2000;20:2439-2450.
223 Hirotsune S, Fleck MW, Gambello MJ, et al. Graded reduction of Pafah1b1 (Lis1) activity results in neuronal migration defects and early embryonic lethality. Nat Genet. 1998;19:333-339.
224 Chae T, Kwon YT, Bronson R, et al. Mice lacking p35, a neuronal specific activator of Cdk5, display cortical lamination defects, seizures, and adult lethality. Neuron. 1997;18:29-42.
225 Wenzel HJ, Robbins CA, Tsai LH, et al. Abnormal morphological and functional organization of the hippocampus in a p35 mutant model of cortical dysplasia associated with spontaneous seizures. J Neurosci. 2001;21:983-998.
226 Hong SE, Shugart YY, Huang DT, et al. Autosomal recessive lissencephaly with cerebellar hypoplasia is associated with human RELN mutations. Nat Genet. 2000;26:93-96.
227 Barkovich AJ, Koch TK, Carrol CL. The spectrum of lissencephaly: report of ten patients analyzed by magnetic resonance imaging. Ann Neurol. 1991;30:139-146.
228 Dobyns WB, Truwit CL. Lissencephaly and other malformations of cortical development: 1995 update. Neuropediatrics. 1995;26:132-147.
229 Cardoso C, Leventer RJ, Dowling JJ, et al. Clinical and molecular basis of classical lissencephaly: mutations in the LIS1 gene (PAFAH1B1). Hum Mutat. 2002;19:4-15.
230 Reiner O, Cahana A, Escamez T, et al. LIS1—no more no less. Mol Psychiatry. 2002;7:12-16.
231 Reiner O, Carrozzo R, Shen Y, et al. Isolation of a Miller-Dieker lissencephaly gene containing G protein beta-subunit–like repeats. Nature. 1993;364:717-721.
232 Vallee RB, Tsai JW. The cellular roles of the lissencephaly gene LIS1, and what they tell us about brain development. Genes Dev. 2006;20:1384-1393.
233 Tsai LH, Gleeson JG. Nucleokinesis in neuronal migration. Neuron. 2005;46:383-388.
234 McManus MF, Nasrallah IM, Pancoast MM, et al. Lis1 is necessary for normal non-radial migration of inhibitory interneurons. Am J Pathol. 2004;165:775-784.
235 Wynshaw-Boris A, Gambello MJ. LIS1 and dynein motor function in neuronal migration and development. Genes Dev. 2001;15:639-651.
236 Cahana A, Escamez T, Nowakowski RS, et al. Targeted mutagenesis of Lis1 disrupts cortical development and LIS1 homodimerization. Proc Natl Acad Sci U S A. 2001;98:6429-6434.
237 Wang Y, Baraban SC. Granule cell dispersion and aberrant neurogenesis in the adult hippocampus of an LIS1 mutant mouse. Dev Neurosci. 2007;29:91-98.
238 Wang Y, Baraban SC. Aberrant dentate gyrus cytoarchitecture and fiber lamination in LIS1 mutant mice. Hippocampus. 2008;18:758-765.
239 Jones DL, Baraban SC. Characterization of inhibitory circuits in the malformed hippocampus of Lis1 mutant mice. J Neurophysiol. 2007;98:2737-2746.
240 Avanzini G, Moshé S, Schwartzkroin P, et al. Animal models for localization related epilepsy. In: Engel JJ, Pedley T, editors. Epilepsy: A Comprehensive Textbook, Vol 1. Philadelphia: Lippincott Williams & Wilkins; 1997:427-442.
241 Lockard JS, Levy RH, DuCharme LL, et al. Cinromide’s metabolite in monkey model: gastric administration and seizure control. Epilepsia. 1980;21:177-182.
242 Lockard JS, Uhlir V, DuCharme LL, et al. Efficacy of standard anticonvulsants in monkey model with spontaneous motor seizures. Epilepsia. 1975;16:301-317.
243 Ribak C, Shapiro L, Seress L, et al. Alumina gel injection models of epilepsy in monkeys. In: Pitkännen A, Schwartzkroin P, Moshé S, editors. Models of Seizures and Epilepsies. Philadelphia: Academic Press; 2005:179-187.
244 De Deyn PP, D’Hooge R, Marescau B, et al. Chemical models of epilepsy with some reference to their applicability in the development of anticonvulsants. Epilepsy Res. 1992;12:87-110.
245 Payan H, Toga M, Berard-Badier M. The pathology of post-traumatic epilepsies. Epilepsia. 1970;11:81-94.
246 Pitkanen A, McIntosh TK. Animal models of post-traumatic epilepsy. J Neurotrauma. 2006;23:241-261.
247 Banfi S, Fonio W, Allievi E, et al. Experimental chronic epilepsy in rats: a screening method for antiepileptic drugs. Pharmacol Res Commun. 1983;15:553-558.