Action of physical and chemical agents on microorganisms
Geoffrey W. Hanlon and Norman A. Hodges
Chapter contents
D value, or decimal reduction time
Antimicrobial effects of moist and dry heat
Resistance of microorganisms to moist and dry heat
Factors affecting heat resistance and its measurement
Effect of ionizing radiations on materials
Factors affecting the radiation resistance of microorganisms
Factors affecting resistance to UV light
Antimicrobial effects of chemical agents
Key points
Introduction
The subject of this chapter is of importance because pharmaceutical scientists have a responsibility for:
• the production of sterile medicaments having no living microorganisms, e.g. injections and eye drops
Thus, the major pharmaceutical interest in microorganisms is that of killing them, or at least preventing their growth. Consequently it is necessary to have both an understanding of the physical processes, e.g. heating and irradiation that are used to kill microorganisms and a knowledge of the more diverse subject of antimicrobial chemicals.
This background knowledge must include an understanding of the kinetics of cell inactivation, the calculation of parameters by which microbial destruction and growth inhibition are measured, and an appreciation of the factors that influence the efficiency of the physical and chemical processes used. These aspects, together with a synopsis of the major groups of antimicrobial chemicals, are the subject of this chapter.
Kinetics of cell inactivation
The death of a population of cells exposed to heat or radiation is often found to follow or approximate to first-order kinetics. In this sense, it is similar to bacterial growth during the logarithmic phase of the cycle, the graphs representing these processes being similar but of opposite slope. Assuming first-order kinetics (the exceptions will be considered later), an initial population of No cells per mL will, after a time t minutes, be reduced to Nt cells per mL, according to the following equations in which k is the inactivation rate constant:
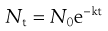 (15.1)
(15.1)
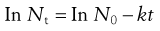 (15.2)
(15.2)
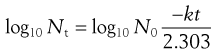 (15.3)
(15.3)
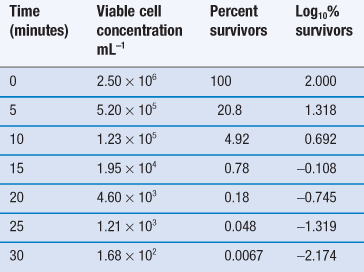
Thus, the data in Table 15.1 may be used to produce a plot of logarithm of cell concentration against exposure time (Fig. 15.1), where the intercept is log No and the slope is −k/2.303. This may be plotted with the logarithm of the percentage of survivors as the ordinate; thus the largest numerical value on this axis is 2.0 (100%). An important feature of Figure 15.1 is the fact that there is no lower endpoint to the ordinate scale – it continues indefinitely. If the initial population was 1000 cells mL−1 the logarithmic value would be 3.0; at 100 cells mL−1 the value would be 2.0; at 10 cells mL−1 1.0, and at 1 cell mL−1 zero. The next incremental point on the logarithmic scale would be −1, which corresponds to 0.1 cells mL−1. It is clearly nonsense to talk of a fraction of a viable cell per mL but this value corresponds to one whole cell in 10 mL of liquid. The next point, −2.0, corresponds to one cell in 100 mL, and so on. Sterility is the complete absence of life, i.e. zero cells mL−1, which has a log value of −∞. Guaranteed sterility would therefore require an infinite exposure time.
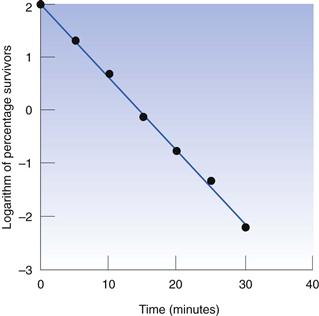
D value, or decimal reduction time
It is characteristic of first-order kinetics that the same percentage change in concentration occurs in successive time intervals. Thus in Figure 15.1 it can be seen that the viable population falls to 10% of its initial value after 7.5 minutes; in the next 7.5-minute period the population again falls to 10% of its value at the start of that period. This time period for a 90% reduction in count is related to the slope of the line and is one of the more useful parameters by which the death rate may be indicated. It is known as the decimal reduction time, or D value, and usually has a subscript showing the temperature at which it was measured, e.g. D121 or D134. It is quite possible to indicate the rate of destruction by the inactivation rate constant calculated from the slope of the line but the significance of this value cannot be as readily appreciated during conversation as that of a D value, and so the former is rarely used.
Z values
When designing steam sterilization processes, it is necessary to know both the D value, which is a measure of the effectiveness of heat at any given temperature, and the extent to which a particular increase in temperature will reduce the D value, i.e. it is necessary to have a measure of the effect of temperature change on death rate. One such measure is the Z value, which is defined as the number of degrees of temperature change required to achieve a 10-fold change in D value, e.g. if the D value for Bacillus stearothermophilus1 spores at 110 °C is 20 minutes and they have a Z value of 9 °C, this means that at 119°C the D value would be 2.0 minutes and at 128 °C the D value would be 0.20 minutes. The relationship between D and Z values is shown in Figure 15.2. The Z value is one of several parameters that relate change in temperature to change in death rate, and is probably the most commonly used and readily understood.
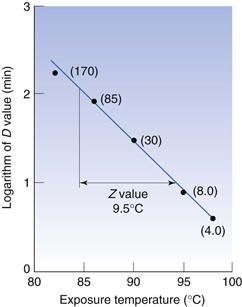
The activation energy obtained from an Arrhenius plot (see Chapter 7) or a temperature coefficient, a Q10 value (change in rate for a 10 °C change in temperature, Chapter 14), do the same but are rarely used.
Alternative survivor plots
It was stated earlier that bacterial death often approximates to first-order kinetics, although exceptions do arise; some of the more common are illustrated in Figure 15.3. The plot labelled A is that conforming to first-order kinetics, which has already been described. A shoulder on the curve, as in case B, is not uncommon and various explanations have been offered. Cell aggregation or clumping may be responsible for such a shoulder, because it would be necessary to apply sufficient heat to kill all the cells in the clump, not merely the most sensitive, before a fall is observed in the number of colonies appearing on the agar. Under normal circumstances one single colony could arise both from one cell alone or, say, from 100 cells aggregated together. In the latter case, if sufficient heat was applied to kill the 99 most sensitive cells in the clump, the colony count would be unaltered. Clumping is not the only explanation, because substantial shoulders may arise using suspensions where the vast majority of cells exist individually.
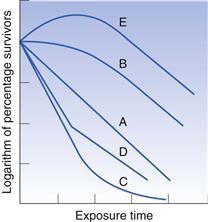
Tailing of survivor curves, as in plot C, is often observed if the initial cell concentration is high. This has been attributed to the presence of mutants that are exceptionally resistant to the lethal agent. If the proportion of mutants was 1 in 106 cells and the initial concentration only 105 cells mL−1 the mutant would not be detected, but an initial population of 109 cells mL−1 would permit easy detection if the inactivation plot were continued down to low levels of survivors. Again there are alternative explanations, one of the most common being that the cells dying during the early exposure period release chemicals which help to protect those that are still alive.
A sharp break in the line, as in D, usually indicates that there are two distinct populations of cells present which have markedly different resistances. Contamination of a cell suspension or culture is a possible explanation, or it may be that a mutant has arisen naturally and the cultural conditions are such that it has a selective advantage and its numbers have increased until it is a substantial proportion of the population.
Plot E is uncommon and is usually only seen as a result of ‘heat activation’ of bacterial spores. This is a situation in which a significant proportion of a population of spores (usually a thermophile) remains dormant and fails to germinate and produce colonies under ‘normal’ conditions. If the suspension receives a heat stimulus or shock which is insufficient to kill the spores, some or all of those that would otherwise remain dormant become activated, germinate and thus produce a rise in the colony count.
First-order kinetics are less commonly observed when microorganisms are being killed by chemicals than when heat or radiation are the lethal agents. This is because the chemical must interact with a target molecule within the cell, and the concentration of both the chemical and the intracellular target might influence death rate; this results in second-order kinetics. In practice, however, the antimicrobial chemical is often present in such a high concentration that the proportion of it that is ‘used up’ by interaction with the cell is negligible; this means its concentration is effectively constant and pseudo first-order kinetics result.
Antimicrobial effects of moist and dry heat
Moist heat (steam) and dry heat (hot air) both have the potential to kill microorganisms but their efficiencies and their mechanisms of action differ. In autoclaves, dry saturated steam, i.e. 100% water vapour with no liquid water present, is used at temperatures between 121 and 135 °C, at which it rapidly kills microorganisms. An advantage of using steam is that it possesses a large latent heat of vaporization, which it transfers to any object upon which it condenses. It is essential to use dry saturated steam if maximal autoclaving efficiency is to be achieved. If the steam is wet, i.e. contains liquid water, penetration of vapour-phase steam into dressings may be retarded. If the steam is superheated, i.e. its temperature has been raised while the pressure remains constant, or the pressure fell while the temperature remains constant, it contains less moisture and latent heat than dry saturated steam at the same temperature. In this case the effect is similar to using a steam–air mixture at that temperature. The process by which steam kills cells is hydrolysis of essential proteins (enzymes) and nucleic acids. In contrast, dry heat causes cell death by oxidative processes, although again it is the proteins and nucleic acids that are the vulnerable targets. Dry heat is much less effective at killing microorganisms than steam at the same temperature. Exposures of not less than 2 hours at 160 °C (or an equivalent temperature/time combination) are recommended in the PhEur for sterilization by dry heat methods. The state of hydration of a cell is thus an important factor determining its resistance to heat.
Resistance of microorganisms to moist and dry heat
Numerous factors influence the observed heat resistance of microbial cells and it is difficult to make comparisons between populations unless these factors are controlled. Not surprisingly, marked differences in resistance exist between different genera, species and strains, and between the spore and vegetative cell forms of the same organism. The resistance may be influenced, sometimes extensively, by: the age of the cell, i.e. lag, exponential or stationary phase; its chemical composition, which in turn is influenced by the medium in which the cell is grown; and by the composition and pH of the fluid in which the cell is heated. It is difficult to obtain strictly comparable heat resistance data for grossly dissimilar organisms, but the values quoted in Table 15.2 indicate the relative order of heat resistance of the various microbial groups. Tabulation of D values at a designated temperature is perhaps the most convenient way of comparing resistance but this is only suitable for first-order kinetics. Alternative methods of comparison include the time to achieve a particular percentage kill or the time required to achieve no survivors; the latter is, of course, dependent upon the initial population level.
Table 15.2
A ‘league table’ of heat resistances of different microorganisms and infectious agents
| Organism or agent | Heat resistance (values are for fully hydrated organisms unless otherwise stated) |
| Prions | The most heat-resistant infectious agent. May survive steam sterilization at 134–138 °C for 1 hour |
| Bacterial spores (endospores) | Little or no inactivation at <80 °C. Some species survive boiling for several hours |
| Fungal spores | Ascospores of Byssochlamys species may survive 88 °C for 60 minutes but most fungal spores are less resistant |
| Actinomycete spores | Spores of Nocardia sebivorans reported to survive for 10 minutes at 90 °C but the majority of species are less resistant |
| Mycobacterium tuberculosis | May survive for 30 minutes at 100 °C in the dry state but when hydrated is killed by pasteurization (63 °C for 30 minutes or 72 °C for 15 seconds) |
| Yeasts | Ascospores and vegetative cells show little difference in resistance. Survival for 20 minutes at 60 °C is typical |
| Most non-sporing bacteria of pharmaceutical or medical importance | D60 of 1–5 minutes is typical of staphylococci and many Gram-negative enteric organisms. Enterococci may be more resistant, and pneumococci may survive for 30 minutes at 110 °C when dry |
| Fungi and actinomycetes | Vegetative mycelia exhibit similar resistance to that of non-sporing bacteria described above |
| Viruses | Rarely survive for >30 minutes at 55–60 °C except perhaps in blood or tissues, but papovaviruses and hepatitis viruses are more resistant |
| Protozoa and algae | Most are no more resistant than mammalian cells and survive only a few hours at 40–45 °C. However, cysts of Acanthamoeba species are more resistant |
The most heat-resistant infectious agents (as distinct from microbial cells) are prions, which are proteins rather than living cells and are the cause of spongiform encephalopathies, e.g. Creutzfeldt–Jakob disease (CJD) and bovine spongiform encephalopathy (BSE or ‘mad cow disease’). Prion proteins are so resistant to heat inactivation that an autoclave cycle of 134–138 °C for 18 minutes has been recommended for the decontamination of prion-contaminated materials, and the efficacy of even this extreme heat treatment has been questioned. The World Health Organization recommends that prion-contaminated surgical instruments are autoclaved at 121 °C for one hour in the presence of 1M sodium hydroxide.
Bacterial endospores are invariably found to be the most heat-resistant cell type, and those of certain species may survive boiling water for many hours. The term ‘endospore’ refers to the spores produced by Bacillus and Clostridium species and is not to be confused with the spores produced by other bacteria, such as actinomycetes, which do not develop within the vegetative cell. The majority of Bacillus and Clostridium species normally form spores which survive in water for 15–30 minutes at 80 °C without significant damage or loss of viability. Because endospores are more resistant than other cells, they have been the subject of a considerable amount of research in the food and pharmaceutical industries and much of the earlier work has been reviewed by Russell (1999).
Mould spores and those of yeasts and actinomycetes usually exhibit a degree of moist heat resistance intermediate between endospores and vegetative cell forms; D-values of the order of 30 minutes at 50 °C would be typical of such organisms, although some species may be substantially more resistant. Bacterial and yeast vegetative cells and mould mycelia all vary significantly in heat resistance: mycobacteria, which possess a high proportion of lipid in their cell wall, tend to be more resistant than others. Protozoa and algae are, by comparison, susceptible to heat and when in the vegetative (uncysted) state they, like mammalian cells, rapidly die at temperatures much in excess of 40 °C. Information on the heat resistance of viruses is limited but the available data suggest that they may vary significantly between types. The majority of viruses are no more heat resistant than vegetative bacteria, but hepatitis viruses, particularly hepatitis B, is less susceptible and exposures of 80 °C for 10 minutes or more are required for effective decontamination.
Resistance to dry heat by different groups of infectious agents and microorganisms usually follows a pattern similar to that in aqueous environments. Again, prions head the ‘league table’ by exhibiting extreme heat resistance and endospores are substantially more resilient than other cell types, with those of B. stearothermophilus and B. subtilis usually more resistant than other species. Exposures of 2 hours at 160 °C are required by the European Pharmacopoeia (2004) to achieve an acceptable level of sterility assurance for materials sterilized by dry heat.
Cells of pneumococci have been reported to survive dry heat at 110 °C for 30 minutes but this represents exceptional resistance for vegetative cells, most of which may be expected to die after a few minutes heating at 100 °C or less.






