[level-membership-for-pathology-category]56
7.2 Humoral immunity58
7.3 Cellular immunity61
7.1. Natural defences and immunity
You should:
• distinguish innate from adaptive immunity
• name the cells important in innate and adaptive immunity and their main properties
• describe the morphological organisation of the immune system
• discuss the principles of specificity, memory, diversity and self-recognition.
The human body is constantly bombarded from the outside world by potentially harmful substances and microorganisms. To protect ourselves against these insults, we have evolved a sophisticated defence system composed of a complex web of cells and chemical mediators that interact to produce an integrated response. The defences can be divided into two main types:
• innate immunity – which has a general protective effect against a wide range of potential insults
• adaptive immunity – which is specific for particular foreign substances and microorganisms.
Together, the two arms of the immune system use a wide variety of different methods to distinguish between the components of the body, or ‘self’, from foreign invaders, or ‘non-self’. Non-self includes unacceptable changes within the body, such as a normal cell becoming neoplastic.
Many different cells take part in host defence; some are involved only in adaptive immune reactions whereas others play a part in innate immunity as well. The characteristics of some of these cells are discussed further in Chapter 9
Innate immunity
Natural (innate) immunity arose early in evolution. It is conferred by physical barriers and cellular responses that are not specific against any one particular insult, but provide an effective first line of defence. The most important types are give below.
Mechanical barriers
The intact skin and mucosal surfaces provide a physical barrier to microorganisms and foreign substances. Tight junctions between the cells and the continuous basement membrane are important components. The keratinised surface of the skin is particularly tough; only a minority of microorganisms can breach intact skin.
Secretion
The acidity of the gastric and vaginal secretions creates a hostile microenvironment which kills many organisms. Lysozyme, found in tears, has an antibacterial action. Paneth cells in the intestine secrete antibacterial proteins called defensins when stimulated by the presence of bacteria. Activation of the complement cascade by the alternative and mannose-binding lectin (MBL) pathways are also part of innate immunity (see Ch. 5).
Secretion and excretion currents
The flow of urine regularly flushes away any bacteria that ascend the urethra, helping to keep the bladder sterile. Bronchial mucus is constantly swept towards the pharynx, carrying with it any contaminating bacteria; when it reaches the pharynx it is coughed out or swallowed into the acid environment of the stomach. Tears have an analogous effect. Stasis of the normal flow of secretions or excretions is a potent cause of infection.
Phagocytes
Although neutrophils and macrophages participate in adaptive immune reactions and their activity is greatly enhanced when stimulated by specific immune responses, they have phagocytic and antimicrobial properties independent of the adaptive immune system. Thus, neutrophils and macrophages also contribute to innate immunity.
Mast cells and eosinophils
Like neutrophils and macrophages, mast cells and eosinophils participate in adaptive immune responses. However, they can also act independently of adaptive immunity. Mast cells (and the closely related basophils) release histamine, cytokines and other substances from their granules when stimulated, promoting inflammation (see Table 13, Ch. 8). Eosinophils secrete a range of substances that are toxic to microorganisms. The actions of mast cells and eosinophils seem to be particularly involved in defence against parasites, but they have other defensive roles and are also involved in allergic reactions.
Natural killer cells
Natural killer (NK) cells are lymphocytes that have the ability to lyse host cells, in particular virally infected cells and neoplastic cells. In this respect, they form part of innate immunity. In addition, NK cells take part in adaptive immunity by their ability to lyse cells opsonised with IgG (antibody-dependent cell-mediated cytotoxicity, see Section 7.4).
Pattern-recognition receptors
These receptors are found on many cells of the immune system and recognise a wide range of molecules found on bacteria, viruses and fungi. They represent a type of host defence found not only in animals but also in bacteria and plants. Binding of a pattern-recognition receptor triggers a response in an effector cell. An important group of pattern-recognition receptors comprises the toll-like receptors. For example, toll-like receptor 1 (TLR1) is expressed on macrophages, dendritic cells and B cells, and recognises a kind of lipopeptide found in bacteria.
Adaptive immunity
Adaptive immunity is specific to the foreign substance which invokes it and becomes quicker and more intense with subsequent exposure to it. Thus the adaptive immune response ‘remembers’ previous challenges with the same substance and can mount a heightened and more rapid response. Adaptive immunity is only seen in higher vertebrates such as mammals.
Cells of the adaptive immune system
Central to the adaptive immune response are two types of lymphocyte, called B cells and T cells. Both these cell types possess membrane receptors which can bind to foreign material (antigen). This triggers a series of intracellular events in the lymphocyte which leads to its activation, multiplication and enhanced ability to destroy or neutralise the antigen. By definition, adaptive immunity (as opposed to innate immunity) involves the antigen-specific receptors on B cells and T cells.
Both types of lymphocyte are derived from precursor cells in the bone marrow. B cell maturation occurs in the bone marrow itself, whereas T cells migrate to the thymus for maturation. B and T cells have distinct functions, but they are interdependent and rely on cooperation with each other and other cells of the immune system.
B cells
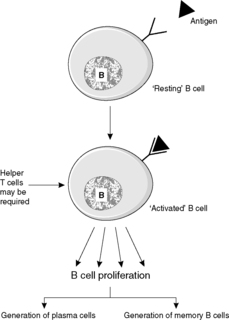
The B cell surface receptor is a type of antibody molecule, namely monomeric IgM. When the appropriate antigen binds to the receptor, the B cell is activated. It multiplies and the offspring ultimately mature into plasma cells, which synthesise and secrete large amounts of antibody into the extracellular fluid; this antibody has the same specificity as the B cell receptor, allowing disposal of the antigen that stimulated it. This process is known as humoral immunity; it is especially important in defence against extracellular bacteria.
T cells
These have a surface antigen recognition system known as the T cell receptor, which is similar to the antibody molecule. There are discrete subsets of T cells which, when triggered by antigen-receptor binding, have functions as diverse as killing cells, coordinating B and T cell responses and stopping the immune response. T cells are particularly involved with the immune response to intracellular organisms (e.g. viruses and certain bacteria such as Mycobacterium tuberculosis). This process is known as cellular immunity.
Organisation of the adaptive immune system
In order to optimise the combined functions of surveillance of the body’s tissues for antigen and production of a coordinated immune response, the cells of the adaptive immune response are organised into structured lymphoid tissues, namely lymph nodes, thymus, mucosa-associated lymphoid tissue and spleen. Lymphocytes are also found in the bone marrow.
The bone marrow and the thymus are sometimes known as primary lymphoid organs, where lymphocytes are produced and mature (B cells in bone marrow, T cells in thymus), whereas the peripheral lymph nodes and other lymphoid sites are called secondary organs, where lymphocytes live and recognise antigens and initiate the immune response. The spleen is primarily involved in clearing the blood of unwanted antigens, while the lymph nodes are responsible for clearing the lymphatic fluid. The mucosal surfaces represent potential portals of entry for microbes, and the mucosa-associated lymphoid tissue forms an important part of the body’s defences. This lymphoid tissue is often concentrated into distinct structures such as the tonsils in the throat, the Peyer patches in the intestine, and the lymphoid tissue of the appendix.
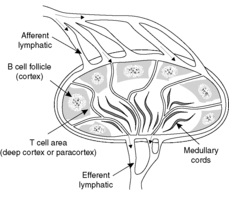
There are similarities in the structure of many of the lymphoid tissues. In general, B cells are found in the discrete nodular structures known as lymphoid follicles. These follicles are surrounded by a sea of T lymphocytes admixed with other cells of the immune response and small blood or lymphatic channels. A lymph node is shown in Figure 17. Lymphocytes can circulate via the lymphatics and bloodstream between the lymphoid organs.
 |
| Figure 17 |
Properties of the immune response
Specificity
The response is specific. Most antigens are foreign macromolecules or microbes that are made up of large numbers of differently shaped structural components (usually proteins or carbohydrates). Some of these components will be recognised by a receptor on the membrane of a restricted number of lymphocytes. Only these few lymphocytes can, therefore, initially respond to the foreign substance. The smallest components recognisable by B cell and T cell receptors are called epitopes or determinants. Some molecules (haptens) are too small to be recognised by themselves, but will be recognised if they are bound to a larger molecule.
Memory
The immune response has memory. Exposure to a previously encountered antigen results in a quicker and larger response. This is, at least in part, through the generation of memory cells during the first challenge with antigen. Memory cells are long-lived cells that have the antigen receptor on their surface and on a subsequent occasion can proliferate quickly to mount a response.
Diversity
The response is diverse. The number of different antigen-specific lymphocytes is vast. We have millions of lymphocytes, each bearing a receptor recognising a slightly different shaped or structured antigenic determinant. When exposed to a particular antigen, e.g. a bacterium, the lymphocyte with the appropriate receptor is selected from this vast available pool and stimulated to multiply. This process is called clonal selection.
The diversity of the system results from the variability in the structure of the lymphocyte surface receptor. The basic structure of the T cell receptor and the immunoglobulin molecule are similar, both being members of the immunoglobulin ‘superfamily’. The enormous diversity of B cell and T cell receptors is due to random rearrangement of a small number of genes within the cell allowing the generation of many different receptors. The result is considerable diversity in the variable region of the antibody molecules and T cell receptors. Molecular techniques such as Southern blot analysis (see Ch. 2) have shown that the portions of DNA containing the immunoglobulin gene are of a different size in B cells and cells that do not produce antibody. This can be explained by the genes being far apart in ‘ordinary’ cells, but close together in B cells. Therefore, B cells must be able to rearrange these genes during their development. This is done in a precise order (e.g. the first rearrangement involves the heavy chains).
Recognition of self
The immune system can distinguish between self and non-self. Thus, under normal circumstances, lymphocytes recognise and respond to molecules that they perceive as foreign but apparently ignore the millions of potential antigens present in normal human tissues and organs. This non-response to our own tissues by our own lymphocytes is known as tolerance and is partly achieved by the early deletion (by apoptosis) of potentially self-reacting cells. If cells reacting with self escape from these control measures, autoimmune disease occurs and our own tissues are damaged and destroyed (see Ch. 8).
7.2. Humoral immunity
The B cell response
Antibodies are synthesised by B cells; they are then either secreted into the tissue fluids for combination with antigen or are inserted into the B cell membrane to act as receptors. If a human is exposed to an antigen, a number of B cells may each recognise a different epitope of the antigen by means of their surface antibody (immunoglobulin) receptors. The proliferation of different B cells with different antibodies being produced by the resultant plasma cells is called a polyclonal response.
Antibody structure
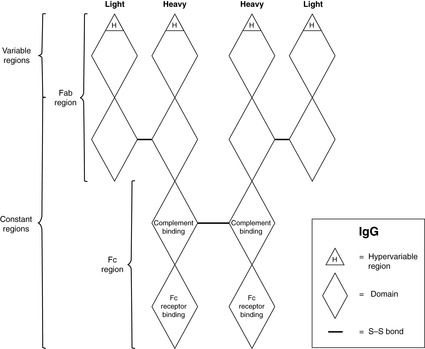
The basic structure of an antibody is shown in Figure 18. In its simplest form, an antibody is composed of two identical light chains and two identical heavy chains. The molecular weight is about 150 000. The light chains are joined to the heavy chains and the heavy chains to each other by covalent disulphide bonds. Overall, the antibody molecule has a ‘Y’ shape, with movement of the arms possible at the hinge. Light chains and heavy chains can be further subdivided into domains, each approximately 110 amino acid residues long. There are four domains per heavy chain and two per light chain. The amino acid sequence in the N-terminal domain of both the light and heavy chains is very variable – the so-called variable region – and it is here that antigen binding occurs. The remaining domains are relatively constant. The different types of immunoglobulin are classified according the Fc region, which is responsible for the actions of the antibody. For example, IgG and IgM opsonise bacteria to which they are bound (because phagocytes have receptors for IgG and IgM Fc regions), and IgE uses its Fc portion to attach to the surface of mast cells and basophils. There are different classes of antibody, which have different functions (Table 9), but they all are based on the simple prototype.
| Antibody class | Functions |
|---|---|
| IgM | First class of antibody, produced before IgG supervenes |
| Pentamer with 10 antigen-binding sites Intravascular: cannot cross placenta | |
| Activates complement and agglutinates foreign substances | |
| IgG | Four subclasses |
| Can attach to phagocytic cells via Fc fragment (opsonisation) | |
| Found in tissues and can cross placenta | |
| Activates complement | |
| IgA | Main antibody in secretions and excretions and in urine and gut contents |
| Two subclasses: can dimerise (molecular weight 320 000) and has an additional secretory piece – this prevents digestion | |
| Stops microbes entering tissues from external surfaces | |
| IgE | Involved in acute allergic reaction (type 1 hypersensitivity) |
| Involved in defence against parasites | |
| IgD | Found on the surface of B cells, but not expressed in activated cells |
| Function unclear, possibly a receptor |
Antibodies have a number of functions. They agglutinate particles by binding them together, and can thus inhibit the movement of motile bacteria. By attaching to molecules they can neutralise the function of those molecules, e.g. microbial cell surface receptors or bacterial toxins. IgG and IgM can activate complement via the classical pathway by interacting with C1. Bound antibody can activate other cells of the immune system such as macrophages, neutrophils and NK cells. In addition, antibodies are opsonins, i.e. when bound to bacteria they facilitate phagocytosis.
Fate of antigen–antibody immune complexes
An antigen–antibody (immune) complex is formed when soluble antigen combines with antibody to form a particle. We are constantly being challenged by antigens, and the formation of small numbers of immune complexes is a normal event. When we mount an antibody response to a significant infection, vast numbers of immune complexes are formed as antibodies bind to and neutralise microbial antigens.
The disposal of immune complexes is important since, potentially, they can lodge in blood vessel walls and cause damage by activating complement and initiating an inflammatory reaction, i.e. type III hypersensitivity (Ch. 8). The macrophages of the liver and spleen perform a large part of this disposal function.
Antigen-presenting cells
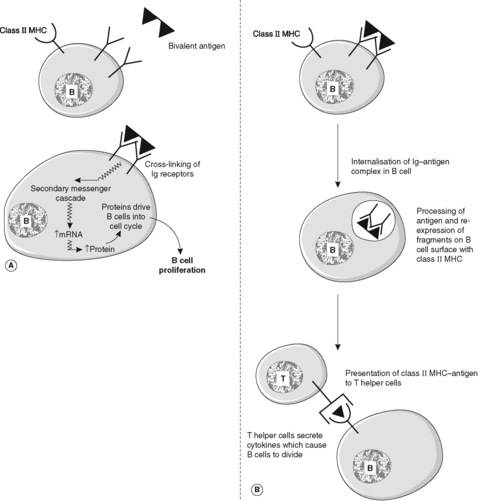
‘Professional’ antigen-presenting cells take antigens into their cytoplasm, process them and then present them to T cells and B cells on their cell surface in association with class II major histocompatibility complex (MHC) molecules (see Section 7.4). They also produce cytokines and other factors which coordinate and amplify the immune response. Antigen-presenting cells are of two main types:
• Cells derived from monocytes produced in the bone marrow. These cells are of two main types:
• macrophages: some are freely mobile, others are fixed at certain sites (e.g. Kupffer cells in the liver)
• dendritic cells: examples include Langerhans’ cells of the skin and follicular dendritic cells of the lymph nodes.
• B cells. B lymphocytes also present antigen with class II molecules.
The antibody response
Certain antigens, e.g. large polysaccharides, can trigger antibody production directly. Others, including most proteins, require T cells – in particular T helper cells – to assist in triggering antibody production. The B cell presents antigen with class II human leucocyte antigen (HLA) molecules to T helper cells which then secrete cytokines that drive the differentiation of B cells into antibody-secreting plasma cells. Helper T cells can be divided broadly into two types called Th1 and Th2, depending on the cytokines they secrete (see Section 7.3).
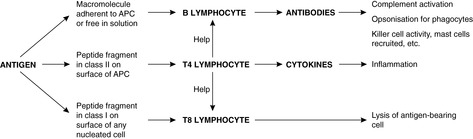
There is a significant difference between the response to antigen of previously unstimulated B cells (the primary antibody response) and the response of memory B cells that have been produced during a previous encounter with the same antigen (the secondary antibody response). The primary response takes longer to develop. Typically, IgM is produced first, followed by IgG. The secondary response is faster; IgM is again produced first but the amount of IgG produced is greatly increased compared to the primary response. The secondary reaction requires helper T cells that recognise the antigen and activate B cells that react to the same antigen. The processes occurring in the B cell to achieve a response to antigen are shown in Figures 19 and 20. Note that free antigen can initiate a B cell response; this is in contrast with the T cell response which always requires processing of antigen by antigen-presenting cells.
 |
| Figure 19 |
7.3. Cellular immunity
T cell-mediated immunity
T cells, which mediate the cellular immune response, have two major functions:
• recognition and attack of cell-surface antigens
• regulation of the T and B cells in the immune response.
There are several subclasses of T cells with different functional characteristics (Table 10). These characteristics are reflected in the cell surface receptor molecules they express. In general, these molecules are named using the CD system (Box 6). CD4+ T helper cells (Th), which represent about 60% of the total T cell population, can only recognise antigen when it is presented together with MHC (see Section 7.4) class II antigen. CD8+ cells can only recognise antigen when presented with class I MHC antigen. This is known as restriction: a response can only be mounted against an antigen that is presented with the correct MHC molecule. The post-stimulus response of the T cell depends on various surface molecules such as CD3.
| T cell subset | Function | CD status | MHC requirement |
|---|---|---|---|
| Helper (Th) | Two main types: Th1 promote cell-mediated immunity and delayed hypersensitivity; Th2 promote humoral immunity | CD4+ CD8− | II |
| Cytotoxic (Tc) | Kill cells with abnormal surface antigens, e.g. virus-infected cells, transplants | CD4− CD8+ | I |
| Suppressor (Ts) | Suppress immune response, including against self-antigens | CD4− CD8+ | I |
Box 6
The CD (cluster of differentiation) system is a way of categorising surface molecules of different types with a variety of different functions. The nomenclature was originally designed for cells of the haemopoietic system, but many other cells also express CD molecules. For example, all mature T cells have CD2 and CD3 on their surface, and are described as CD2+ CD3+. Mature helper T cells express CD4 in addition but, unlike cytotoxic cells, do not express CD8; they are thus CD4+ CD8-. The CD antigens can be recognised in tissue samples using immunohistochemistry or flow cytometry.
Class II antigens display a sample of extracellular proteins that have been endocytosed. With a few exceptions, class II antigens are only found on antigen-presenting cells. Class I antigens are found on virtually all nucleated cells and platelets. Class I antigens display a sample of the intracellular protein content; this can stimulate the CD8+ cytotoxic T cells (Tc) to destroy such cells if they display a foreign protein, e.g. as a result of viral infection.
How do T cells attack antigen?
T cell attack on antigen is seen in many circumstances, e.g. infection with intracellular microorganisms (such as viruses and mycobacteria), fungi and protozoans.
Cytotoxic T cells
Cytotoxic T cells express CD8. They kill cells infected with intracellular organisms, which are inaccessible to antibodies. Microbial antigens are displayed on the host cell surface in association with class I antigen. The infected cell is in effect alerting the cytotoxic T cell, which kills the cell by one of two mechanisms:
2. Expression of Fas ligand (see Ch. 4) or release of granzymes that enter through the perforin pores; these events initiate apoptosis by triggering the caspase cascade. One advantage of apoptosis over oncosis in this context is that the endonucleases activated during apoptosis degrade the DNA not only of the cell but also of any infective organisms within the cell.
Helper T cells
Helper T cells express CD4 and mediate their effects via cytokines. There are two main types: Th1 and Th2.
Th1 cells promote cell-mediated immunity by stimulating macrophages and NK cells. This reaction is often called delayed hypersensitivity; a classic example is mycobacterial infections, e.g. tuberculosis (caused by Mycobacterium tuberculosis). Th1 cells with receptors for antigens of M. tuberculosis are activated and release cytokines which attract macrophages to the site of infection and activate them. The activation of macrophages promotes intracellular killing of infecting organisms. Th1 cells also inhibit the Th2 response.
Th2 cells secrete cytokines that promote humoral immune reactions, especially the IgE response, and activate eosinophils. They also inhibit the Th1 response. One hypothesis that is based on the Th1/Th2 concept is that allergic reactions represent an inflammatory response overly weighted towards Th2.
Suppressor T cells
Suppressor (Ts) cells express CD8 and function to inhibit (deactivate) the immune response. Relatively little is known about how they do this. They may not require processing and display of antigen by antigen-presenting cells and, therefore, may not be MHC restricted, despite being CD8α. They probably perform their suppressor functions by secreting cytokines which stop cell activation or proliferation or directly cause cell death.
7.4. Other components of the immune response
Cytokines
Cytokines include interleukins, which are involved in lymphocyte proliferation and differentiation, interferons, which protect against viral infection, and colony-stimulating factors, which regulate the production of bone marrow blood cells (Box 7). The role of cytokines in inflammation is considered in more depth in Chapter 9
Box 7
Although it was originally named ‘tumour necrosis factor’ because of its effects on tumour cells, TNF alpha (TNF-α) (like most cytokines) has a wide variety of different actions, all of which are modulated by other cytokines. TNF-α is secreted by several different cell types, but macrophages are particularly important. Examples of its effects include, among others:
• activation of neutrophils and macrophages to promote microbial killing
• stimulation of T cells and B cells
• stimulation of endothelial cells to promote leucocyte adhesion
• raising the thermoregulatory set point of the hypothalamus, i.e. it produces fever
• stimulation of the liver to produce acute phase reactants such as C-reactive protein.
Thus, a macrophage secreting TNF-α can have autocrine actions on itself, paracrine actions on neighbouring inflammatory cells and endothelial cells, and endocrine actions on the liver and hypothalamus.
The major histocompatibility complex
The MHC system occurs in all vertebrates and is the major system that distinguishes self from non-self. The human MHC system is known as the human leucocyte antigen (HLA) system. The HLA genes are found on the short arm of chromosome 6. They code for cell surface glycoproteins and are important in many cell-mediated immunological reactions.
The six loci that code for these cell surface glycoproteins are known as DP, DQ, DR (class II genes producing class II antigens) and A, B and C (class I) (Table 11). At each of these loci, there are several possible alleles (versions of the gene). Therefore, the number of possible combinations on both chromosomes is astronomical.
| Each of the class I and class II molecules is polymorphic (has many variants). One person can have only two variants of each molecule, one coded by each parent. | |||
| Class | Chromosome location | Number of molecules | Cell location |
|---|---|---|---|
| I | 6 | 3(HLA A, B, C) | All nucleated cells |
| II | 6 | 3(HLA DP, DQ, DR) | Antigen-presenting cells |
Class I HLA antigens are transmembrane glycoproteins and consist of two polypeptide chains which grip peptide fragments to be presented to cytotoxic T cells (CD8+). If the peptide is not a normal part of the cell, such as a viral protein, it is recognised and the cell destroyed. Almost all nucleated cells display class I antigens.
Class II HLA molecules are also composed of two polypeptide chains spanning the cell membrane. The main function of these molecules is to present foreign antigens to CD4+ T helper cells; they are potent stimulators of T cell proliferation. Generally, class II molecules are expressed only by a few specialised cells, the ‘professional’ antigen-presenting cells, namely dendritic cells, macrophages and B cells.
Since the advent of tissue typing, it has been found that some diseases are commoner in people with a particular type, or combination of types, of HLA. An example is the arthritic disease ankylosing spondylitis: the HLA B27 allele is found about 20 times more commonly in people with ankylosing spondylitis than in those without the disease.
Complement
The complement system is an enzymatic cascade which can be activated by antibody (classical pathway) or antigen (alternative and MBL pathways). Complement proteins are made in the liver and are numbered 1–9 (see Ch. 5).
Natural killer cells
This special type of non-B, non-T lymphocyte does not have immunoglobulin or T cell receptors and does not require prior exposure to antigen for its activity; therefore it is classified as part of innate immunity. The main function of NK cells is to destroy cells that are damaged or infected. NK cells can do this by recognising cells with reduced levels of MHC I expression, which occurs in neoplastic and virally infected cells. The NK cell then destroys the cell by insertion of perforin channels. In addition, NK cells express CD16, a receptor for the Fc portion of IgG, on their surface. Therefore, they can bind to IgG-coated cells and destroy them; this type of killing is called antibody-dependent cell-mediated cytotoxicity. Other ways in which NK cells recognise target cells include receptors for complement component C3b, and for heat shock proteins that are upregulated in injured cells.
Summary
All the components discussed in this chapter are interlinked in their functions and requirements for activation (Table 12 and Figure 21).
| Component | Function |
|---|---|
| B lymphocytes | Recognition of antigen Presentation of antigenic peptides to CD4 T cells |
| Secretion of antibody | |
| CD4 T helper lymphocytes | Recognition of antigens on MHC class II cells after endocytosis and processing |
| Promoting local inflammation by recruitment and activation of macrophages | |
| Helping T and/or B cell responses | |
| CD8 cytotoxic T lymphocytes | Recognition of intracellular antigens presented on MHC class I after cytosolic processing |
| Killing cells which present abnormal proteins in class I MHC | |
| Macrophages | Phagocytosis and destruction of microbes, dead cells and tissue debris, promoted by coatings of antibody and complement Walling off infection in granulomas |
| Presentation of antigens to CD4 cells | |
| Antibodies | Acting as receptors on the surface of B cells Neutralising toxins and microbial invaders |
| Marking microbes and abnormal body cells for destruction by complement and effector cells | |
| Cytokines | Control of cellular activities |
Self-assessment: questions
One best answer questions
2. Binding of a toll-like receptor on a macrophage to its ligand is most likely to cause the macrophage to:
a. become activated
b. die by apoptosis
c. die by oncosis
d. secrete immunoglobulin
e. secrete eosinophil major basic protein
3. Immunoglobulins that act as efficient opsonins are:
a. IgA and IgE
b. IgA and IgG
c. IgE and IgG
d. IgG and IgM
e. IgM and IgE
4. Which one of the following is most effective at clearing extracellular bacteria?
a. apoptosis induced by CD8+ T cells
b. apoptosis induced by NK cells
c. complement and antibody action
d. fibrin deposition
e. suppressor T cell activity
1. In the saliva, the immunoglobulin with the highest concentration is:
a. IgA
b. IgD
c. IgE
d. IgG
e. IgM
True-false questions
1. The following are correctly paired:
a. IgG – cannot cross placenta
b. CD8 – class II MHC
c. IgM – complement activation
d. B cells – cell-mediated immunity
e. Interleukin-1 – fever
2. The following statements are true:
a. dendritic cells and Langerhans’ cells can present antigen
b. Th1 cells promote cell-mediated immunity
c. CD3 is found on T cells
d. proliferation of cytotoxic (CD8+) T cells usually requires the presence of helper (CD4+) cells
e. in humans, the genes for the MHC system are found on chromosome 7
Extended matching items questions (EMIs)
EMI 1
Theme: Antigen binding and processing
A. antigen-T cell receptor binding
B. binding to IgE Fc
C. binding to IgE variable region
D. binding to IgG Fc
E. binding to IgG variable region
F. binding to IgM Fc
G. binding to IgM variable region
H. insertion of perforin channels
I. presentation of ingested antigens
J. presentation of peptides derived from the cell cytoplasm
For each of the following statements, select the most appropriate action from the list.
1. The principal function of the HLA class I molecule.
2. The principal function of the HLA class II molecule.
3. The means by which mast cells attach antibody to their membranes.
4. A means of cell execution shared by CD8+ T lymphocytes and NK cells.
5. The way IgG molecules recognise antigen.
Case history questions
Case history 1
A 10-year-old girl has a sore throat. A throat swab is taken and the culture grows Streptococcus bacteria. During the course of her illness, her general practitioner notes that her neck lymph nodes are enlarged.
1. What changes would you expect to see in this girl’s lymph nodes?
2. How would it differ if the infection was viral?
3. How might the second bout of illness relate to her sore throat?
4. Name an autoimmune condition affecting the heart that is associated with streptococcal sore throat and describe its pathogenesis.
Self-assessment: answers
One best answer
1. b. A hapten is too small to be recognised by B cells and T cells, but if it is bound to a larger carrier molecule it can be recognised and initiate an immune response. An example of a hapten is the toxic chemical in poison ivy. This chemical binds to proteins in the skin that act as carriers. As a result, the immune system becomes sensitised to the toxin and mounts an immune response on subsequent exposure to poison ivy (a type IV hypersensitivity reaction, see Ch. 8), causing the blistering rash. An epitope is defined as the smallest identifiable part of an antigenic molecule that can be recognised by a B cell or T cell receptor. An immunoglobulin is an antibody molecule. Secretory component is a molecule produced by epithelial cells that combines with IgA when it is secreted. A tolerogen is a substance that is recognised by B cells and T cells but induces tolerance (i.e. a failure to respond immunologically to the tolerogen) rather than an immune reaction.
2. a. Pattern-recognition receptors such as the toll-like receptors represent a type of host defence that is ancient in evolutionary terms. Immunoglobulin is not secreted by macrophages but by plasma cells, which are derived from B lymphocytes. Eosinophil major basic protein is highly toxic to parasites; it is stored by eosinophils in their granules and released when they are activated.
3. d. IgG and IgM are opsonins. Neutrophils and macrophages have receptors for the Fc portions of these types of antibody.
4. c. Extracellular bacteria are accessible to serum antibody and complement. IgG, IgM and complement components all act as opsonins. In addition, the complement membrane attack complex can cause lysis of bacteria. Cellular immunity, in which cytotoxic T cells and NK cells participate, is more important in defence against viruses, fungi, intracellular bacteria and protozoa. Fibrin deposition occurs in inflammation (Ch. 9) but does not clear bacteria. Suppressor T cells damp down the immune reaction.
5. a. IgA is secreted as a dimer into the fluids produced by mucous membranes, e.g. saliva, tears, gastrointestinal juice, sweat and breast milk.
True-false answers
1.
a. False. IgG can cross the placenta, the other types of immunoglobulins cannot. If IgM antibodies to a particular organism are present within the fetal circulation, this indicates that it is the fetus itself which is infected and mounting an immune response; IgG antibodies could be maternal in origin. The half-life in the circulation of IgG is about 3–4weeks, so IgG of maternal origin that enters the fetus persists after birth and provides useful defence against infection in the first few months of extrauterine life.
b. False. CD4+ (helper) T cells recognise antigen in association with class II MHC molecules, and then secrete cytokines to activate other cells of the immune system. CD8+ (cytotoxic) T cells use class I molecules, and may then kill the cell if they recognise the antigen as foreign.
c. True. IgM is very efficient at complement activation.
d. False. Cell-mediated immunity involves responses by cytotoxic T cells in association with macrophages.
e. True. Fever (pyrexia) is caused by a number of cytokines, including interleukin-1, which raise the thermoregulatory set point of the hypothalamus so that the homeostatic mechanisms of temperature control maintain body temperature higher than normal. A substance that does this is called a pyrogen. Endotoxin (bacterial lipopolysaccharide) is also a pyrogen. The function of fever is unclear, but it may assist in the defence against infection: for example, raising the body temperature seems to inhibit viral replication.
2.
a. True. Dendritic cells and Langerhans’ cells load antigen into class II HLA molecules and present it to T helper cells.
b. True. The cytokines produced by Th1 cells, e.g. interleukin-2, TNF-+ and interferon gamma, promote cell-mediated immunity to a greater extent than humoral immunity.
c. True. There are numerous CD molecules on the surface of lymphocytes, macrophages and antigen-presenting cells. The structure and function of many of these protein molecules are now well characterised. CD3 is found on all T cells and is associated with the T cell receptor. It is a useful marker of T cell differentiation in the diagnostic laboratory.
d. True. An important function of the cytotoxic T cell is to destroy viral-infected cells, recognised by the association of class I MHC molecules with viral antigens on the surface of the infected cell. In order to proliferate, the CD8+ Tc cell requires stimulation by cytokines from CD4+ Th cells.
EMI answers
EMI 1
Theme: Antigen binding and processing
1. J. Proteins degraded in proteasomes are presented on the cell surface by being loaded into MHC class I molecules. In this way, potential immunogens such as virus components that might otherwise evade immune surveillance can be shown to cells of the immune system. This procedure is different from the one used by antigen-presenting cells to ingest antigen and process it for display (see next answer).
2. I. The class II molecules are found on antigen-presenting cells, which take up antigen from outside the cell. The antigen is processed in a phagolysosome and is ultimately bound to an MHC class II molecule, which is then transported to the cell surface where it displays the antigen.
3. B. IgE binds to mast cells and basophils before it encounters its specific antigen. When antigen cross-links IgE molecules on the cell surface, the result is degranulation with release of the various mediators (histamine etc.).
4. H. Perforin creates a hole in the target cell membrane. The contents leak out and the cell dies by lysis. Alternatively, the perforin channel allows granzyme molecules to enter the target cell and initiate apoptosis.
5. E. The variable region of an immunoglobulin molecule binds to the antigen. It is the great variety of shapes exhibited by variable regions that accounts for the huge range of antigens specifically recognised by antibodies.
Case history answers
Case history 1
1. Bacterial infections tend to elicit a strong humoral (antibody) immune response. The local lymph nodes draining the site of infection will be stimulated to produce plasma cells. In a normal lymph node, B cell areas are found in the outer cortex in areas known as follicles – densely packed aggregates of cells. When B cell production is stimulated, these follicles acquire a germinal centre. Transformation of B cells occurs in germinal centres with the production of numerous memory cells. B cells differentiate into plasma cells in the cortical areas deep to and between the follicles – the paracortex. Plasma cells then enter the medullary vascular system (medullary cords) and produce antibody as they leave in the efferent lymphatics, draining the site of infection. The local lymph nodes will have large cortical follicles with prominent germinal centres. (See Ch. 24.)
2. Viral infections generally stimulate a T cell response – cytotoxic T cells (CD8+) can recognise and destroy infected ells if the viral antigen is displayed on the cell’s surface by a class I MHC molecule. The T cell areas of the lymph node are found in the paracortex and massive expansion of this region can occur during viral infections. The B cell follicles will be relatively inconspicuous.
3. Soluble antigen from the streptococci has combined with antibody to form complexes that have deposited in the girl’s kidneys causing them to function abnormally (see Section 23.3 in Ch. 23). In particular, they have lodged in the glomeruli, evoking an inflammatory reaction which causes the glomerular capillaries to become leaky. Blood can then be found in the urine (haematuria). Other organs that may be damaged by immune complex deposition are the joints and skin. Fortunately, in most cases, once the inflammatory response has died down, normal kidney function is restored. The delay in onset of the problem is a result of the time taken to produce large amounts of antibody.
4. One important autoimmune disease which occurs after a streptococcal infection is rheumatic fever. In this disease, now rare in industrialised countries, antibodies produced against streptococcal antigens cross-react with antigens in the patient’s own tissues. The heart is often affected and the inflammatory damage may lead to fibrosis and distortion of the valves. This can lead to heart failure and death if the valve is not replaced. (See Ch. 14.)
[/level-membership-for-pathology-category][not-level-membership-for-pathology-category]56
7.2 Humoral immunity58
7.3 Cellular immunity61
7.1. Natural defences and immunity
You should:
• distinguish innate from adaptive immunity
• name the cells important in innate and adaptive immunity and their main properties
• describe the morphological organisation of the immune system
• discuss the principles of specificity, memory, diversity and self-recognition.
The human body is constantly bombarded from the outside world by potentially harmful substances and microorganisms. To protect ourselves against these insults, we have evolved a sophisticated defence system composed of a complex web of cells and chemical mediators that interact to produce an integrated response. The defences can be divided into two main types:
• innate immunity – which has a general protective effect against a wide range of potential insults
• adaptive immunity – which is specific for particular foreign substances and microorganisms.
Together, the two arms of the immune system use a wide variety of different methods to distinguish between the components of the body, or ‘self’, from foreign invaders, or ‘non-self’. Non-self includes unacceptable changes within the body, such as a normal cell becoming neoplastic.
Many different cells take part in host defence; some are involved only in adaptive immune reactions whereas others play a part in innate immunity as well. The characteristics of some of these cells are discussed further in Chapter 9
Innate immunity
Natural (innate) immunity arose early in evolution. It is conferred by physical barriers and cellular responses that are not specific against any one particular insult, but provide an effective first line of defence. The most important types are give below.
Mechanical barriers
The intact skin and mucosal surfaces provide a physical barrier to microorganisms and foreign substances. Tight junctions between the cells and the continuous basement membrane are important components. The keratinised surface of the skin is particularly tough; only a minority of microorganisms can breach intact skin.
Secretion
The acidity of the gastric and vaginal secretions creates a hostile microenvironment which kills many organisms. Lysozyme, found in tears, has an antibacterial action. Paneth cells in the intestine secrete antibacterial proteins called defensins when stimulated by the presence of bacteria. Activation of the complement cascade by the alternative and mannose-binding lectin (MBL) pathways are also part of innate immunity (see Ch. 5).
Secretion and excretion currents
The flow of urine regularly flushes away any bacteria that ascend the urethra, helping to keep the bladder sterile. Bronchial mucus is constantly swept towards the pharynx, carrying with it any contaminating bacteria; when it reaches the pharynx it is coughed out or swallowed into the acid environment of the stomach. Tears have an analogous effect. Stasis of the normal flow of secretions or excretions is a potent cause of infection.
Phagocytes
Although neutrophils and macrophages participate in adaptive immune reactions and their activity is greatly enhanced when stimulated by specific immune responses, they have phagocytic and antimicrobial properties independent of the adaptive immune system. Thus, neutrophils and macrophages also contribute to innate immunity.
Mast cells and eosinophils
Like neutrophils and macrophages, mast cells and eosinophils participate in adaptive immune responses. However, they can also act independently of adaptive immunity. Mast cells (and the closely related basophils) release histamine, cytokines and other substances from their granules when stimulated, promoting inflammation (see Table 13, Ch. 8). Eosinophils secrete a range of substances that are toxic to microorganisms. The actions of mast cells and eosinophils seem to be particularly involved in defence against parasites, but they have other defensive roles and are also involved in allergic reactions.
Natural killer cells
Natural killer (NK) cells are lymphocytes that have the ability to lyse host cells, in particular virally infected cells and neoplastic cells. In this respect, they form part of innate immunity. In addition, NK cells take part in adaptive immunity by their ability to lyse cells opsonised with IgG (antibody-dependent cell-mediated cytotoxicity, see Section 7.4).
Pattern-recognition receptors
These receptors are found on many cells of the immune system and recognise a wide range of molecules found on bacteria, viruses and fungi. They represent a type of host defence found not only in animals but also in bacteria and plants. Binding of a pattern-recognition receptor triggers a response in an effector cell. An important group of pattern-recognition receptors comprises the toll-like receptors. For example, toll-like receptor 1 (TLR1) is expressed on macrophages, dendritic cells and B cells, and recognises a kind of lipopeptide found in bacteria.
Adaptive immunity
Adaptive immunity is specific to the foreign substance which invokes it and becomes quicker and more intense with subsequent exposure to it. Thus the adaptive immune response ‘remembers’ previous challenges with the same substance and can mount a heightened and more rapid response. Adaptive immunity is only seen in higher vertebrates such as mammals.
Cells of the adaptive immune system
Central to the adaptive immune response are two types of lymphocyte, called B cells and T cells. Both these cell types possess membrane receptors which can bind to foreign material (antigen). This triggers a series of intracellular events in the lymphocyte which leads to its activation, multiplication and enhanced ability to destroy or neutralise the antigen. By definition, adaptive immunity (as opposed to innate immunity) involves the antigen-specific receptors on B cells and T cells.
Both types of lymphocyte are derived from precursor cells in the bone marrow. B cell maturation occurs in the bone marrow itself, whereas T cells migrate to the thymus for maturation. B and T cells have distinct functions, but they are interdependent and rely on cooperation with each other and other cells of the immune system.
B cells
The B cell surface receptor is a type of antibody molecule, namely monomeric IgM. When the appropriate antigen binds to the receptor, the B cell is activated. It multiplies and the offspring ultimately mature into plasma cells, which synthesise and secrete large amounts of antibody into the extracellular fluid; this antibody has the same specificity as the B cell receptor, allowing disposal of the antigen that stimulated it. This process is known as humoral immunity; it is especially important in defence against extracellular bacteria.
T cells
These have a surface antigen recognition system known as the T cell receptor, which is similar to the antibody molecule. There are discrete subsets of T cells which, when triggered by antigen-receptor binding, have functions as diverse as killing cells, coordinating B and T cell responses and stopping the immune response. T cells are particularly involved with the immune response to intracellular organisms (e.g. viruses and certain bacteria such as Mycobacterium tuberculosis). This process is known as cellular immunity.
Organisation of the adaptive immune system
In order to optimise the combined functions of surveillance of the body’s tissues for antigen and production of a coordinated immune response, the cells of the adaptive immune response are organised into structured lymphoid tissues, namely lymph nodes, thymus, mucosa-associated lymphoid tissue and spleen. Lymphocytes are also found in the bone marrow.
The bone marrow and the thymus are sometimes known as primary lymphoid organs, where lymphocytes are produced and mature (B cells in bone marrow, T cells in thymus), whereas the peripheral lymph nodes and other lymphoid sites are called secondary organs, where lymphocytes live and recognise antigens and initiate the immune response. The spleen is primarily involved in clearing the blood of unwanted antigens, while the lymph nodes are responsible for clearing the lymphatic fluid. The mucosal surfaces represent potential portals of entry for microbes, and the mucosa-associated lymphoid tissue forms an important part of the body’s defences. This lymphoid tissue is often concentrated into distinct structures such as the tonsils in the throat, the Peyer patches in the intestine, and the lymphoid tissue of the appendix.
There are similarities in the structure of many of the lymphoid tissues. In general, B cells are found in the discrete nodular structures known as lymphoid follicles. These follicles are surrounded by a sea of T lymphocytes admixed with other cells of the immune response and small blood or lymphatic channels. A lymph node is shown in Figure 17. Lymphocytes can circulate via the lymphatics and bloodstream between the lymphoid organs.
|
|
| Figure 17 |
Properties of the immune response
Specificity
The response is specific. Most antigens are foreign macromolecules or microbes that are made up of large numbers of differently shaped structural components (usually proteins or carbohydrates). Some of these components will be recognised by a receptor on the membrane of a restricted number of lymphocytes. Only these few lymphocytes can, therefore, initially respond to the foreign substance. The smallest components recognisable by B cell and T cell receptors are called epitopes or determinants. Some molecules (haptens) are too small to be recognised by themselves, but will be recognised if they are bound to a larger molecule.
Memory
The immune response has memory. Exposure to a previously encountered antigen results in a quicker and larger response. This is, at least in part, through the generation of memory cells during the first challenge with antigen. Memory cells are long-lived cells that have the antigen receptor on their surface and on a subsequent occasion can proliferate quickly to mount a response.
Diversity
The response is diverse. The number of different antigen-specific lymphocytes is vast. We have millions of lymphocytes, each bearing a receptor recognising a slightly different shaped or structured antigenic determinant. When exposed to a particular antigen, e.g. a bacterium, the lymphocyte with the appropriate receptor is selected from this vast available pool and stimulated to multiply. This process is called clonal selection.
The diversity of the system results from the variability in the structure of the lymphocyte surface receptor. The basic structure of the T cell receptor and the immunoglobulin molecule are similar, both being members of the immunoglobulin ‘superfamily’. The enormous diversity of B cell and T cell receptors is due to random rearrangement of a small number of genes within the cell allowing the generation of many different receptors. The result is considerable diversity in the variable region of the antibody molecules and T cell receptors. Molecular techniques such as Southern blot analysis (see Ch. 2) have shown that the portions of DNA containing the immunoglobulin gene are of a different size in B cells and cells that do not produce antibody. This can be explained by the genes being far apart in ‘ordinary’ cells, but close together in B cells. Therefore, B cells must be able to rearrange these genes during their development. This is done in a precise order (e.g. the first rearrangement involves the heavy chains).
Recognition of self
The immune system can distinguish between self and non-self. Thus, under normal circumstances, lymphocytes recognise and respond to molecules that they perceive as foreign but apparently ignore the millions of potential antigens present in normal human tissues and organs. This non-response to our own tissues by our own lymphocytes is known as tolerance and is partly achieved by the early deletion (by apoptosis) of potentially self-reacting cells. If cells reacting with self escape from these control measures, autoimmune disease occurs and our own tissues are damaged and destroyed (see Ch. 8).
7.2. Humoral immunity
The B cell response
Antibodies are synthesised by B cells; they are then either secreted into the tissue fluids for combination with antigen or are inserted into the B cell membrane to act as receptors. If a human is exposed to an antigen, a number of B cells may each recognise a different epitope of the antigen by means of their surface antibody (immunoglobulin) receptors. The proliferation of different B cells with different antibodies being produced by the resultant plasma cells is called a polyclonal response.
Antibody structure
The basic structure of an antibody is shown in Figure 18. In its simplest form, an antibody is composed of two identical light chains and two identical heavy chains. The molecular weight is about 150 000. The light chains are joined to the heavy chains and the heavy chains to each other by covalent disulphide bonds. Overall, the antibody molecule has a ‘Y’ shape, with movement of the arms possible at the hinge. Light chains and heavy chains can be further subdivided into domains, each approximately 110 amino acid residues long. There are four domains per heavy chain and two per light chain. The amino acid sequence in the N-terminal domain of both the light and heavy chains is very variable – the so-called variable region – and it is here that antigen binding occurs. The remaining domains are relatively constant. The different types of immunoglobulin are classified according the Fc region, which is responsible for the actions of the antibody. For example, IgG and IgM opsonise bacteria to which they are bound (because phagocytes have receptors for IgG and IgM Fc regions), and IgE uses its Fc portion to attach to the surface of mast cells and basophils. There are different classes of antibody, which have different functions (Table 9), but they all are based on the simple prototype.
| Antibody class | Functions |
|---|---|
| IgM | First class of antibody, produced before IgG supervenes |
| Pentamer with 10 antigen-binding sites Intravascular: cannot cross placenta | |
| Activates complement and agglutinates foreign substances | |
| IgG | Four subclasses |
| Can attach to phagocytic cells via Fc fragment (opsonisation) | |
| Found in tissues and can cross placenta | |
| Activates complement | |
| IgA | Main antibody in secretions and excretions and in urine and gut contents |
| Two subclasses: can dimerise (molecular weight 320 000) and has an additional secretory piece – this prevents digestion |
