[level-membership-for-basic-science-category]
3 Midbrain, hindbrain, spinal cord
Box 3.1 Four decussations
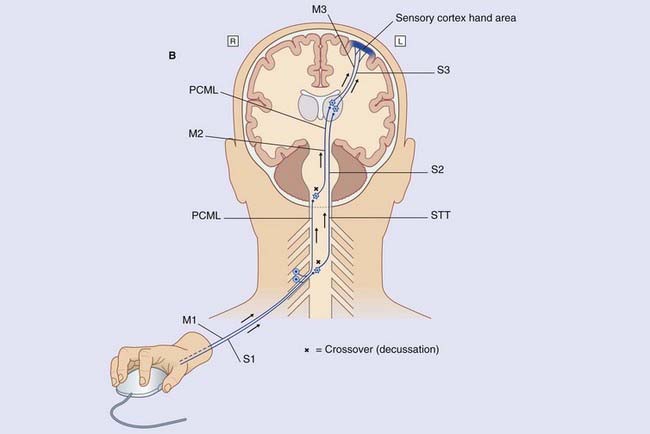
The most significant deep tissue sensory organs are neuromuscular spindles (muscle spindles) contained within skeletal muscles. The primary afferents supplying the muscle spindles of the intrinsic muscles of the hand belong to large unipolar neurons whose axons (labeled M1) ascend ipsilaterally (on the same side of the spinal cord) within the posterior funiculus, as already seen in Figure 3.5. They synapse in the nucleus cuneatus in the medulla oblongata. The multipolar second-order neurons send their axons across the midline in the sensory decussation (seen in Figure 3.6).The axons ascend (M2) through pons and midbrain before synapsing on third-order neurons (M3) projecting from thalamus to sensory cortex.
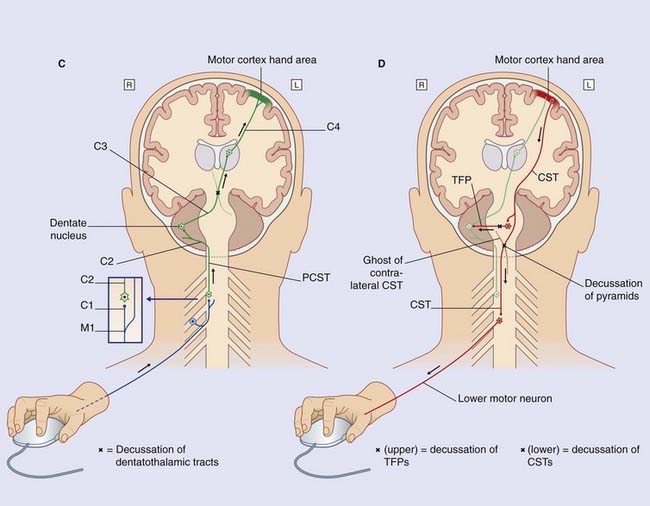
(D) Motor output. Multipolar neurons in the left motor cortex now fire impulses along the upper motor neurons that constitute the corticospinal tract (CST), which crosses to the opposite side in the motor decussation, as already noted in Figure 3.6. The CST synapses on lower motor neurons projecting from the anterior horn of the spinal gray matter to activate flexor muscles of the index finger and local stabilizing muscles.
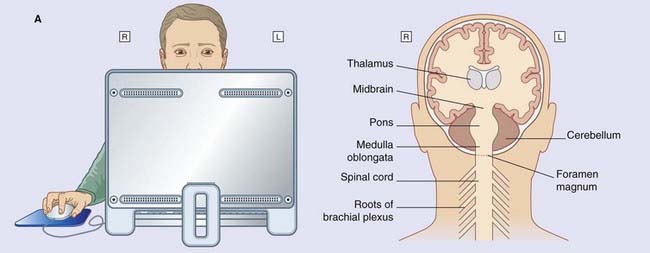
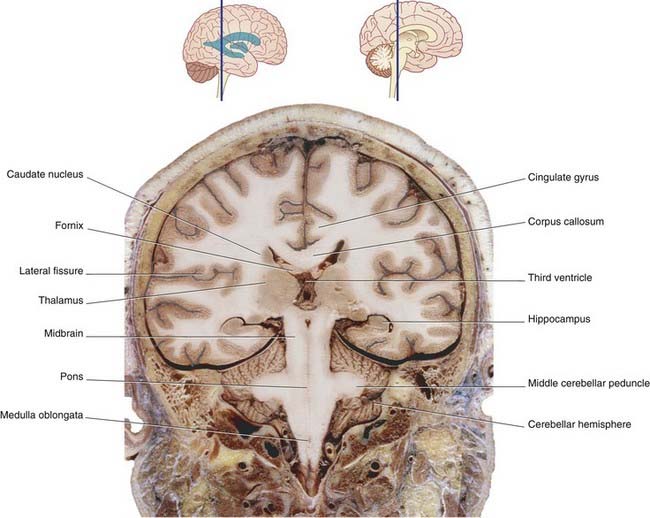
The midbrain connects the diencephalon to the hindbrain. As explained in Chapter 1, the hindbrain is made up of the pons, medulla oblongata, and cerebellum. The medulla oblongata joins the spinal cord within the foramen magnum of the skull.
Brainstem
Ventral view (Figures 3.1 and 3.2A)
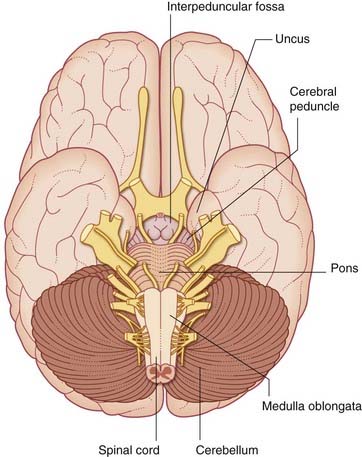
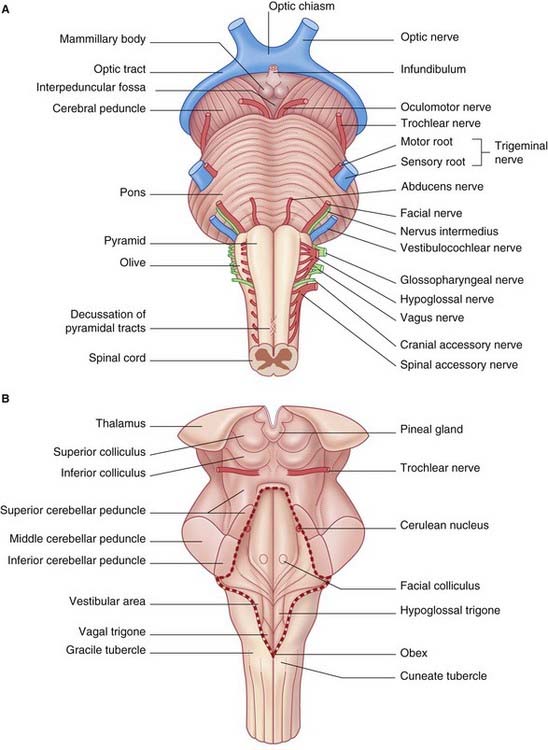
Pons
At the lower border of the pons are the attachments of the abducens (VI), facial (VII), and vestibulocochlear (VIII) nerves (see Table 3.1).
| Number | Name |
|---|---|
| I | Olfactory, enters the olfactory bulb from the nose |
| II | Optic |
| III | Oculomotor |
| IV | Trochlear |
| V | Trigeminal |
| VI | Abducens |
| VII | Facial |
| VIII | Vestibulocochlear |
| IX | Glossopharyngeal |
| X | Vagus |
| XI | Accessory |
| XII | Hypoglossal |
Sectional views
In the midbrain, the central canal of the embryonic neural tube is represented by the aqueduct. Behind the pons and upper medulla oblongata (Figure 3.3), it is represented by the fourth ventricle, which is tent-shaped in this view. The central canal resumes at midmedullary level; it is continuous with the central canal of the spinal cord, although movement of cerebrospinal fluid into the cord canal is negligible.
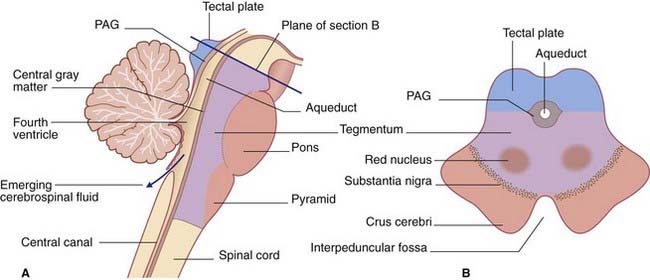
The tegmentum of the entire brainstem is permeated by an important network of neurons, the reticular formation. The tegmentum also contains ascending sensory pathways carrying general sensory information from the trunk and limbs. Illustrated in Figures 3.4–3.6 are the posterior column–medial lemniscal (PCML) pathways, which inform the brain about the position of the limbs in space. At spinal cord level, the label PCML is used here because these pathways occupy the posterior columns of white matter in the cord. In the brainstem, the label PCML is used because they continue upward as the medial lemnisci.
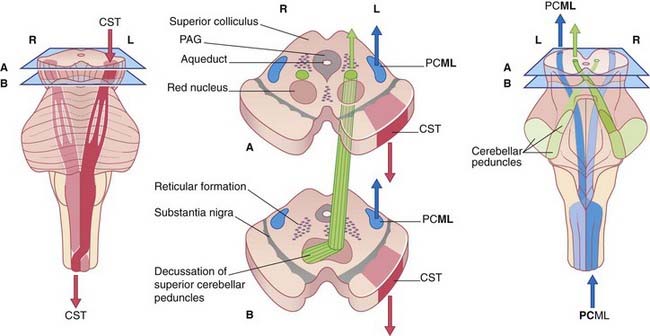
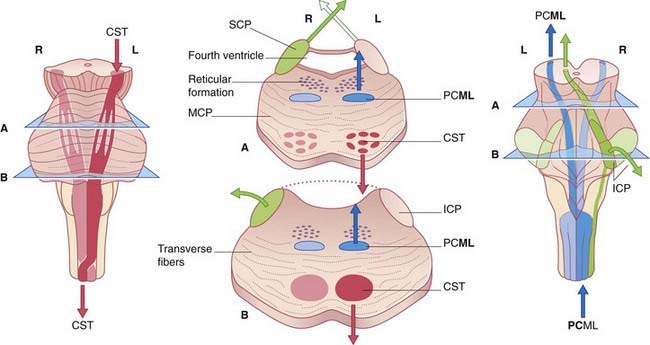
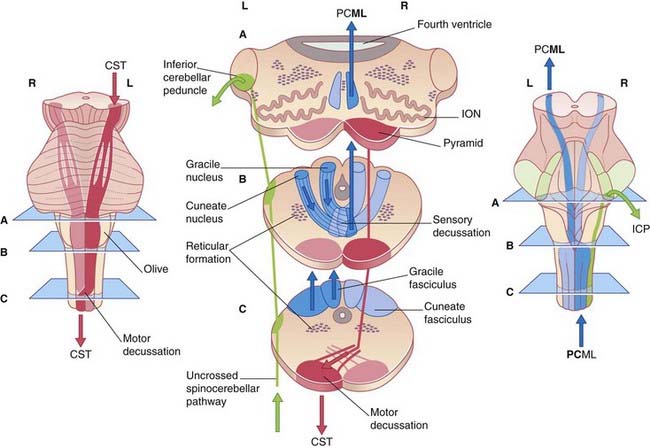
Note that, in the medulla oblongata, the PCML and CST decussate: one of each pair intersects with the other to gain the contralateral (opposite) side of the neuraxis (brainstem–spinal cord). The four most important decussations are illustrated in Box 3.1.
Midbrain (Figure 3.4)
Pons (Figure 3.5)
In the upper section, the cavity of the fourth ventricle is bordered laterally by the superior cerebellar peduncles, which are ascending (arrows) to decussate in the lower midbrain. In the floor of the ventricle is the central gray matter. The medial lemniscus occupies the ventral part of the tegmentum on each side. The basilar region contains millions of transverse fibers, some of which separate the CST into individual fascicles. The transverse fibers enter the cerebellum via the middle cerebellar peduncles and appear to form a bridge (hence, pons) connecting the cerebellar hemispheres. But the individual transverse fibers arise on one side of the pons and cross to enter the contralateral cerebellar hemisphere. The transverse fibers belong to the giant corticopontocerebellar pathway, which travels from the cerebral cortex of one side to the contralateral cerebellar hemisphere, as depicted in Box 3.1.
Medulla oblongata (Figure 3.6)
Sections of brainstem in situ are in Figures 3.7–3.11.
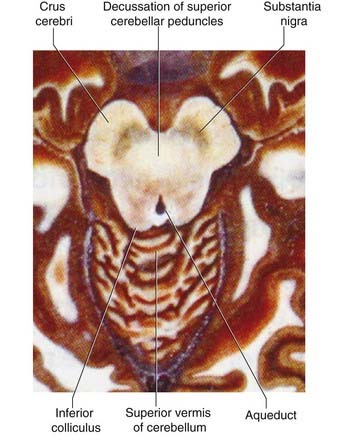
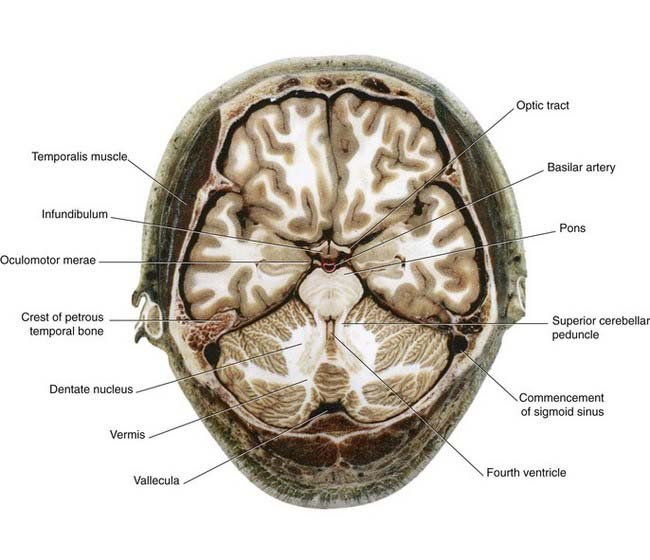
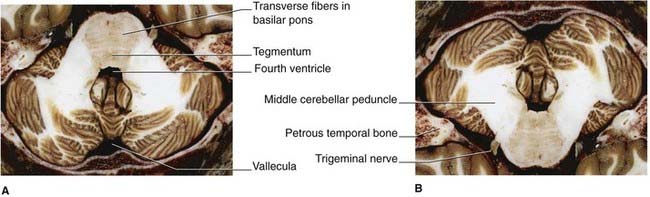
Figure 3.10 Horizontal section taken through the middle of the pons. (A) In axial brain scans, the pons would be in the position shown, i.e. in the roof of the fourth ventricle. (B) In standard anatomic descriptions including histologic sections (cf. Ch. 17), the pons occupies the floor of the fourth ventricle, as shown here. Note the massive size of the middle cerebellar peduncles.
(From Liu et al. 2003, with permission of Shantung Press of Science and Technology.)
Spinal Cord
General features
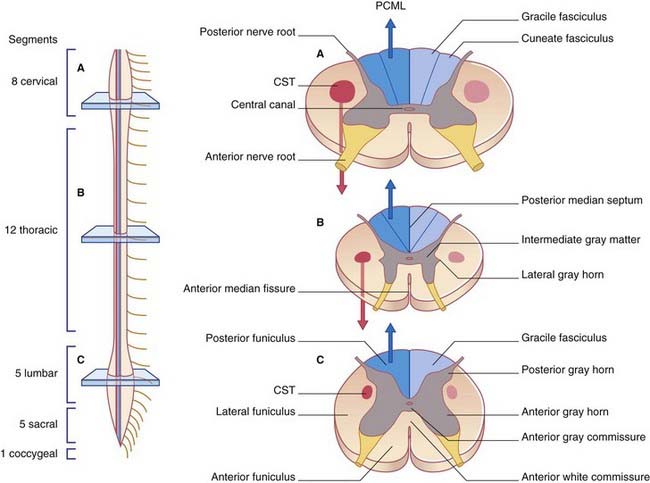
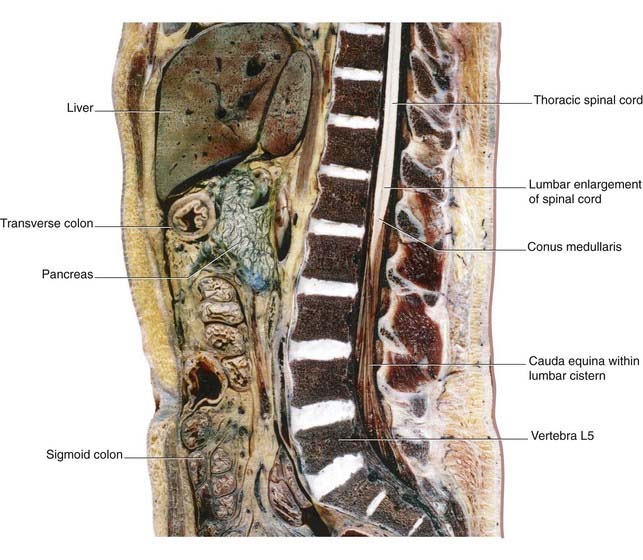
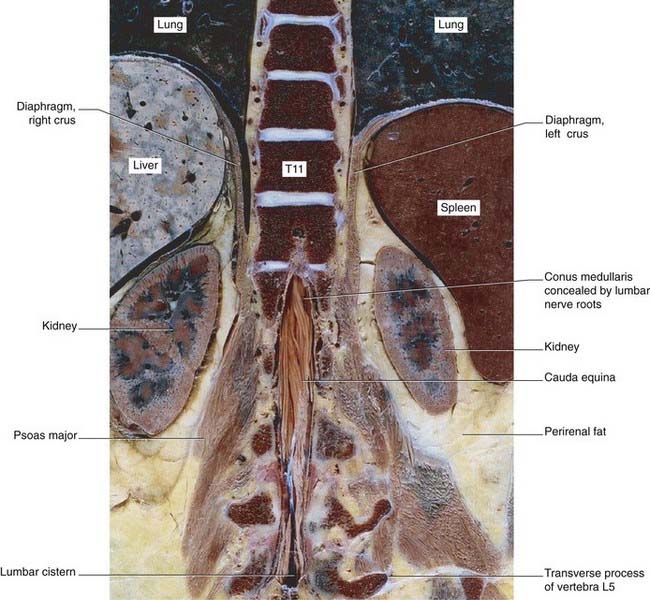
The spinal cord occupies the upper two-thirds of the vertebral canal. Thirty-one pairs of spinal nerves are attached to it by means of anterior and posterior nerve roots (Figure 3.12A). The cord shows cervical and lumbar enlargements that accommodate nerve cells supplying the upper and lower limbs.
Internal anatomy
In transverse sections, the cord shows butterfly-shaped gray matter surrounded by three columns or funiculi of white matter (Figure 3.12B): an anterior funiculus in the interval between the anterior median fissure and the emerging anterior nerve roots; a lateral funiculus between the anterior and posterior nerve roots; and a posterior funiculus between the posterior roots and the posterior median septum.
The CST descends the cord within the lateral funiculus. Its principal targets are neurons in the anterior gray horn concerned with activation of skeletal muscles. Special note: In Chapter 16, it will be seen that a small, anterior CST separates from the main bundle and descends within the anterior funiculus. Accordingly, the proper name of the bundle depicted here is the lateral CST.
In the cord, the PCML pathway is represented by the gracile and cuneate fasciculi. The fasciculi are composed of the central processes of peripheral sensory neurons supplying muscles, joints, and skin. Processes entering from the lower part of the body form the gracile (’slender’) fasciculus; those from the upper part form the cuneate (‘wedge-shaped’) fasciculus (Figures 3.13 and 3.14).
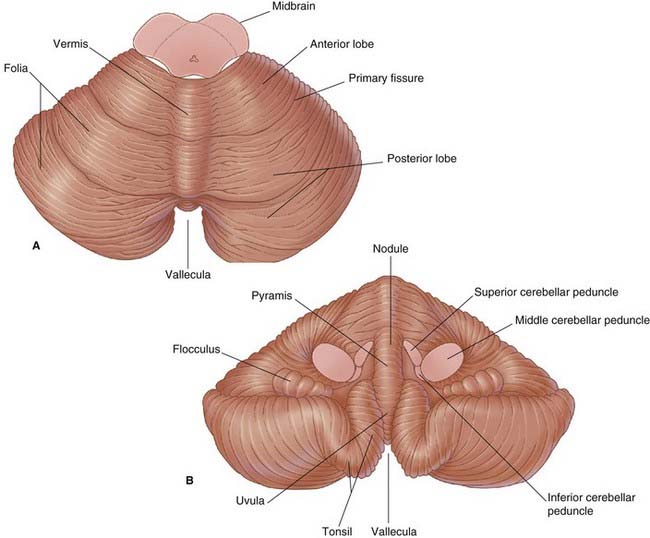
Cerebellum
The cerebellum is made up of two hemispheres connected by the vermis in the midline (Figure 3.15). The vermis is distinct only on the undersurface, where it occupies the floor of a deep groove, the vallecula. The hemispheres show numerous deep fissures, with folia between. About 80% of the cortex (surface gray matter) is hidden from view on the surfaces of the folia.
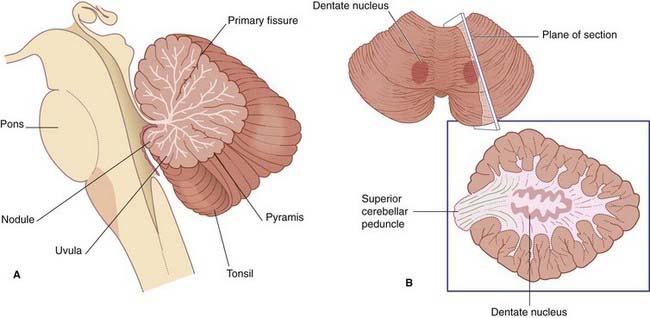
The white matter contains several deep nuclei. The largest of these is the dentate nucleus (Figure 3.16).
Clarke C, Howard R. Nervous system structure and function. In: Clarke C, Howard R, Rossor M, Shorvon S, editors. Neurology: A Queen Square Textbook. London: Wiley-Blackwell; 2009:13-74.
England MA, Wakely J. A colour atlas of the brain and spinal cord. St. Louis: Mosby; 2005.
Kretschmann H-J, Weinrich W. Cranial neuroimaging and clinical neuroanatomy. Stuttgart: Thieme; 2004.
Liu S, et al, editors. Atlas of human sectional anatomy. Jinan: Shantung Press of Science and Technology, 2003.
[/level-membership-for-basic-science-category][not-level-membership-for-basic-science-category]
3 Midbrain, hindbrain, spinal cord
Box 3.1 Four decussations
The most significant deep tissue sensory organs are neuromuscular spindles (muscle spindles) contained within skeletal muscles. The primary afferents supplying the muscle spindles of the intrinsic muscles of the hand belong to large unipolar neurons whose axons (labeled M1) ascend ipsilaterally (on the same side of the spinal cord) within the posterior funiculus, as already seen in Figure 3.5. They synapse in the nucleus cuneatus in the medulla oblongata. The multipolar second-order neurons send their axons across the midline in the sensory decussation (seen in Figure 3.6).The axons ascend (M2) through pons and midbrain before synapsing on third-order neurons (M3) projecting from thalamus to sensory cortex.
(D) Motor output. Multipolar neurons in the left motor cortex now fire impulses along the upper motor neurons that constitute the corticospinal tract (CST), which crosses to the opposite side in the motor decussation, as already noted in Figure 3.6. The CST synapses on lower motor neurons projecting from the anterior horn of the spinal gray matter to activate flexor muscles of the index finger and local stabilizing muscles.
The midbrain connects the diencephalon to the hindbrain. As explained in Chapter 1, the hindbrain is made up of the pons, medulla oblongata, and cerebellum. The medulla oblongata joins the spinal cord within the foramen magnum of the skull.
Brainstem
Ventral view (Figures 3.1 and 3.2A)
Pons
At the lower border of the pons are the attachments of the abducens (VI), facial (VII), and vestibulocochlear (VIII) nerves (see Table 3.1).
| Number | Name |
|---|---|
| I | Olfactory, enters the olfactory bulb from the nose |
| II | Optic |
| III | Oculomotor |
| IV | Trochlear |
| V | Trigeminal |
| VI | Abducens |
| VII | Facial |
| VIII | Vestibulocochlear |
| IX | Glossopharyngeal |
| X | Vagus |
| XI | Accessory |
| XII | Hypoglossal |
[/not-level-membership-for-basic-science-category]
