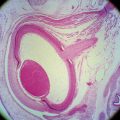
Chapter 5 Crystalline Lens
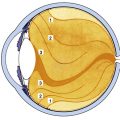
The crystalline lens is an avascular, transparent elliptic structure that aids in focusing light rays on the retina. The lens is located within the posterior chamber, anterior to the vitreous chamber and posterior to the iris (Figure 5-1). The lens is suspended from the surrounding ciliary body by zonular fibers. It is malleable, and ciliary muscle contraction can cause a change in lens shape, increasing the dioptric power of the eye. The mechanism that causes an increase in lens power is accommodation, which allows near objects to be focused on the retina.
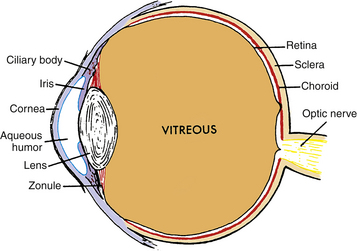
FIGURE 5-1 Diagram showing relationship of the lens and zonule to other ocular structures.
(From Paterson CA, Delamere NA: The lens. In Hart WM Jr, editor: Adler’s physiology of the eye, ed 9, St Louis, 1992, Mosby.)
The posterior lens surface is attached to the anterior vitreous face by the hyaloid capsular ligament, a circular ring adhesion. Within this ring is a potential space, the retrolental space (of Berger), an area of nonadhesion between the vitreous and the lens (see Figure 6-12).
Lens Dimensions
The lens is biconvex, with the posterior surface having the steeper curve. The anterior radius of curvature measures 8 to 14 μm, and the posterior surface radius of curvature measures 5 to 8 μm. The centers of the anterior and posterior surfaces are called the poles, and the lens thickness is the distance from the anterior to posterior pole. The thickness of the unaccommodated lens is 3.5 to 5 mm and it increases 0.02 mm each year throughout life.1–3 The lens diameter is the nasal-to-temporal measurement and in the infant is 6.5 mm. The diameter reaches 9 mm during the teenage years and does not change significantly,4–6 although some report a small age-related increase in diameter.7 The equator is the largest circumference of the lens at a location between the two poles.
The refractive power of the unaccommodated lens is approximately 20 diopters (D) and depends on the (1) surface curvatures, (2) refractive index, (3) change in index between the lens and surrounding environment, and (4) length of the optical path. The lens has a gradient refractive index because of changes in optical density throughout the lens; the index increases from the anterior to the center of the lens and decreases toward the posterior surface. The refractive index is a factor of protein concentration within the lens fibers.8 The variations in index provide additional refractive power.9,10 The power of the lens increases in accommodation, with the maximum accommodative amplitude, 14 D, reached between ages 8 and 12 years.11 Accommodative power decreases with age, approaching zero after 50 years.12
Embryologic Development
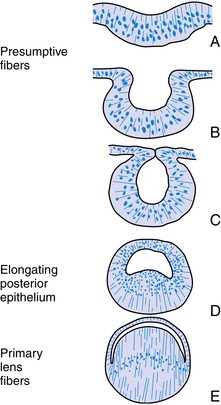
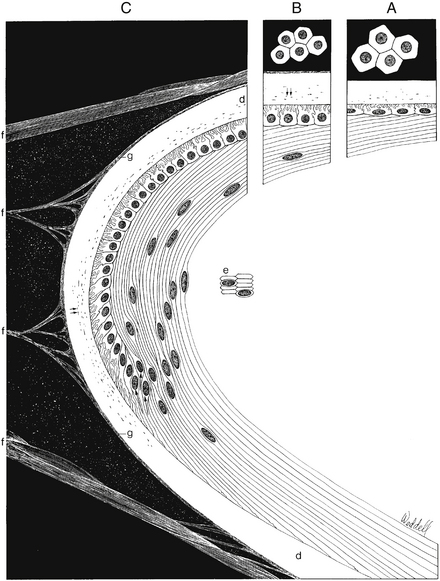
The structure of the adult lens is determined during embryologic development. The lens vesicle, the first lenslike structure observable in the developing embryo, is composed of a layer of epithelial cells that form a hollow sphere. The cells are positioned so that the apical surface lines the lumen of this sphere. The posterior cells differentiate and elongate, forming the primary lens fibers. As these fibers grow and reach the anterior cells (Figure 5-2), the center of the sphere fills. Thus the adult lens has no posterior epithelium because it was used to form these first lens fibers. During the rest of the life of the lens, cell division occurs in the germinative zone of the epithelium just anterior to the lens equator, and the cells thus formed elongate to form secondary lens fibers that are laid down outer to all earlier fibers. With age, the lens continues to grow as it forms new fibers (see Chapter 7).
Histology
Lens Capsule
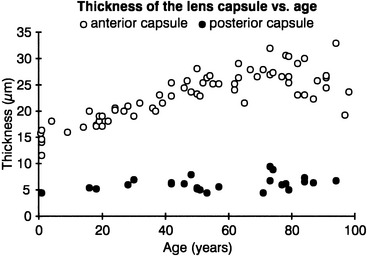
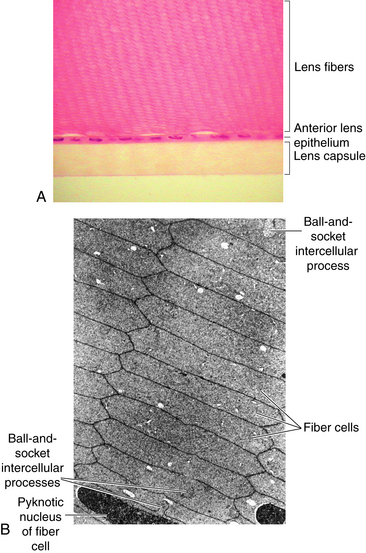
The lens capsule is a transparent envelope that surrounds the entire lens. The capsule is a basement membrane and with time becomes the thickest in the body.13 Its thickness varies with location. At the posterior pole, it is thinnest (approximately 3.5 μm) and does not appreciably increase with age; the thickness at the equator increases slightly with age, and on average is 7 μm. The capsule thickness at the anterior pole increases with age from approximately 11 to 15 μm. The annular region surrounding the anterior pole appears to be the thickest. It too increases with age from approximately 13.5 to 16 μm.14 The capsule consists primarily of collagen; it contains no elastic fibers but is highly elastic because of the lamellar arrangement of the fibers.4,15 It encloses all lens components and helps to mold the shape of the lens. The capsule would prefer to take a more spherical shape, but this tendency is counteracted by the pull from the zonular fibers. The zonular fibers insert into the capsule, merging with it in an area from the equator to near both poles. This coincides with the annular area mentioned previously. This outer superficial zone of the capsule is called the zonular lamella and consists of zonules interconnected with matrix.4 The lens capsule provides some barrier function preventing large molecules, such as albumin and hemoglobin,4 from entering the lens. The anterior lens capsule is produced by the anterior epithelium and thickens with age. The posterior lens capsule may receive some contribution from the basal membrane of lens fibers, but the thickness of the posterior capsule changes minimally throughout life (Figure 5-3).16–18
Lens Epithelium
Adjacent to the anterior lens capsule is a layer of cuboidal epithelium—the anterior lens epithelium (Figure 5-4). These cells secrete the anterior capsule throughout life and are the site of metabolic transport mechanisms. As noted, no posterior epithelium is present because it was used during embryologic development to form the primary lens fibers. The basal aspect of the epithelial cell is adjacent to the capsule, and the apical portion is oriented inward toward the center of the lens. The lateral membranes of the epithelial cells are joined by desmosomes and gap junctions.17,19–22 Although zonula occludens were once thought to join the epithelial cells,23 more recent studies report few, if any, tight junctions.18

The band of cells in the preequatorial region that lies just anterior to the equator is called the germinal zone, the location of cell mitosis. Cell division continues throughout life; as each cell divides, a daughter cell migrates posteriorly towards the equator, withdraws from the cell cycle, and differentiates into a lens fiber. Each newly formed cell elongates; the basal aspect stretches toward the posterior pole and the apical aspect toward the anterior pole (Figure 5-5). This process occurs all around the equator, with fibers stretching toward the poles from all aspects of the lens periphery. As the cells in each layer elongate, the cellular nuclei move with the cytoplasm. A line drawn to connect the dots of these nuclei would have an arcuate shape toward the anterior aspect, a configuration called the lens bow (see Figure 5-2). Eventually, as it loses all cellular organelles, the elongated cell becomes a lens fiber. The anterior end of the lens fiber (the apical surface) insinuates itself between the epithelial layer and the underlying lens fibers. The new fibers are laid down outer to the older fibers; the more superficial fibers are longer than deeper fibers, and the youngest cells lie directly below the epithelium and the capsule. All fibers formed from mitosis in the germinative zone are called secondary lens fibers. Once it loses its nucleus, the mature lens fiber has lost its attachment to the basement membrane.17
Lens Fibers
Lens fiber production continues throughout life, with the new lens fibers being laid down outer to the older fibers; growth results in concentric layers of secondary lens fibers. The structure of the lens is similar to an onion; each layer of fibers approximates a layer of an onion, but then each layer is made up of adjacent fibers. A section through the equator of the lens shows that the fibers cut in cross section are mostly hexagonal in shape and arranged in concentric rings (Figure 5-6). The cross section dimensions of a fiber are approximately 3 by 9 μm.24 Each fiber has a long crescent shape, with the broad sides parallel to the lens surface and the narrow sides at an angle to the surface and located near the lens poles.

Lens fiber cytoplasm contains a high concentration of proteins, known as crystallins, which account for approximately 40% of the net weight of the fiber. The distribution and concentration of crystallins contribute to the gradient refractive index.25 The crystallin concentration varies from approximately 15% in the cortex to 70% in the nucleus.26 A cytoskeletal network of microtubules and filaments provides structure and also provides stability by being anchored to the plasma membrane.18,25 The lateral membranes have numerous and elaborate interdigitations along the fiber length that take various shapes, such as ball-and-socket and tongue-in-groove junctions, and allow for sliding between fibers4,17,18 (Figure 5-7). The fibers also are joined by desmosomes.
Because the lens has no vascular supply and the fibers lose their cellular organelles as they age, some cell-to-fiber and fiber-to-fiber mechanism of communication is necessary. There is an extensive network of gap junctions throughout the lens along the lateral fiber membranes to account for the facility with which nutrients and ions move within the lens.17,27 These gap junctions have a different packing arrangement and different protein connexins, forming the channel than do the typical gap junctions.21 The gap junctions are not evenly distributed throughout the lens, with few near the poles, more toward the equator, and seemingly fewer junctions in deeper layers.13,21,22 Micropinocytic vesicles at apical and basal aspects of fiber membranes and significant areas of membrane fusion also allow movement of material from fiber to fiber, contributing to communication between fibers in addition to the gap type of junctions described.13,20,22,28
Epithelium-Fiber Interface
The border between the apical membrane of the anterior epithelium and the apical membrane of the elongating fiber is known as the epithelium-fiber interface (EFI). Nutrients and ions are exchanged across the EFI. It was once assumed that such movement was facilitated by gap junctions, but disagreement now exists on whether gap junctions are present.19,22,27,28 Gap junctions are usually found on the lateral cell membrane, and the EFI involves apical surfaces. Few true gap junctions have been visualized in tissue preparations. Minimal coupling occurs between the epithelium and fibers in the central zone (i.e., near the poles and sutures), but such junctions increase toward the germinative zone.21 Pinocytosis does occur at this interface, facilitating exchange.18
Divisions of Lens
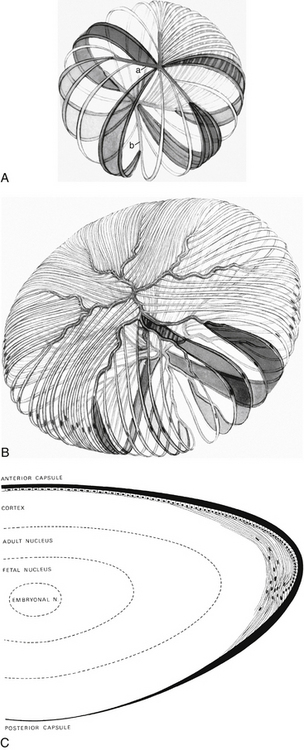
The primary lens fibers from the elongating posterior epithelium form the very center of the lens, the embryonic nucleus, and all subsequent lens fibers are laid down outer to this core. Cell mitosis then begins in the preequatorial region of the epithelium, the new cell migrates toward the equator, and then elongates, forming a lens fiber. All such fibers formed are secondary lens fibers. The fetal nucleus includes the embryonic nucleus and the fibers surrounding it that are formed before birth. The adult nucleus is considered to include the embryonic and fetal nuclei and the fibers formed from birth to sexual maturation. The lens cortex contains the fibers formed after sexual maturation (Figure 5-8, C). The cortex can be divided into superficial, internal, and deep zones,13 but these are arbitrary divisions and generally are not clinically significant. Some consider the fibers formed before sexual maturation the “juvenile nucleus,” those added before middle age the “adult nucleus,” and the remaining fibers the “cortex.”25 The lens cortex has the lowest and the embryonic nucleus has the highest index of refraction.
Lens Sutures
As the lens fibers reach the poles they meet with the other fibers in their layer, forming a junction known as a suture. The anterior suture is formed by the joining of the apical aspects of the fibers, and the posterior suture is formed by the joining of the basal aspects.4,13 The secondary fibers formed during embryologic development meet in three branches, forming Y sutures. The anterior suture is an upright-Y shape and the posterior suture an inverted-Y shape (see Figure 5-8, A).18 As growth continues and the lens becomes larger, the sutures become asymmetric and dissimilar. The limbs of the anterior and posterior sutures are offset, and the complexity of the sutures contributes to lens transparency.29 The sutures formed after birth are more stellate shaped; sutures formed through early adulthood have 6 to 9 branches; and 9 to 15 complex branched stars are formed in middle to old age29 (see Figure 5-8, B).
Zonules (of Zinn)
The lens is attached to the ciliary body by a group of threadlike fibers, the zonules (of Zinn), or the suspensory ligament of the lens (Figure 5-9). The fibers belong to a category termed microfibrils. The macromolecular composition that accounts for the remarkable extensibility of the zonules is complex and still undetermined.15,30 The zonules appear to be formed of extracellular matrix, that includes fibrillin and elastin, both of which have a role in the synthesis of elastic fibers.31 However, biomolecular analysis indicates that there are no true elastic fibers present in the zonules.31
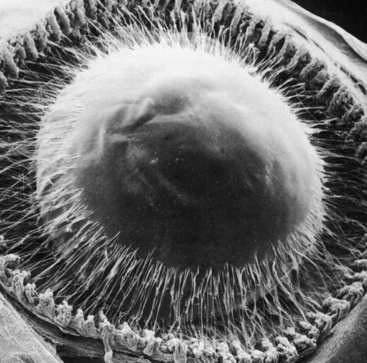
The fibers arise from the basement membrane of the nonpigmented ciliary epithelium in the pars plana and from the valleys between the ciliary processes in the pars plicata.4 They form two columnlike structures on both sides of a ciliary process and end at the lens capsule (see Figure 3-12). Most fibers attach to the lens capsule at the preequatorial and postequatorial regions; few attach directly at the equator.32 The zonules are interwoven into the components of the capsule. Those that attach to the lens are known as primary zonules. Secondary zonules join the primary zonules with each other or connect processes to one another or to the pars plana, and “tension” fibers anchor the primary zonules to the ciliary valleys to form a fulcrum.32
Accommodation
When the emmetropic eye is viewing a distant object, the ciliary muscle is relaxed, the diameter of the ciliary ring is relatively large, and the zonules are in a stretched configuration exerting tension on the lens capsule holding the lens in the unaccommodated state, such that the image lies on the retina. When a near object is to be focused on the retina, an increase in the refractive power of the eye must occur. This increase in power is called accommodation and is accomplished by a change in lens shape brought about by contraction of the ciliary muscle. Von Helmholtz33 is credited with determining that the following occur during accommodation:
These factors result in a thickened lens with more sharply curved surfaces and thus increased lens power. The stimulus that initiates the accommodative mechanism is retinal blur.34 The accommodative mechanism is dependent on cone stimulation with little influence by rods.35
The lens becomes more sharply curved, and because of the thickness variation of the capsule, a bulge is most apparent at the center of the anterior surface.36,37 The posterior surface increases in curvature only slightly; however, the steepness of the anterior surface does not become greater than that of the posterior surface.12 As the lens thickens axially, the equatorial diameter decreases, and the anterior lens pole moves toward the cornea. It was previously thought that there was no posterior lens movement, but a small amount of movement of the posterior pole in a posterior direction has been identified.8,38,39 The curvature of the internal surfaces, seen at the zones of discontinuity and the boundaries of the nuclei, mimics the changes in the surface curvatures and contributes to the increase in the total dioptric power.8 This thickening of the lens from anterior to posterior occurs in the nuclear region, but the thickness of the lens cortex remains unchanged.40 The changes occurring in accommodation described in 1855 by Helmholtz have been demonstrated using current technology.5,41
The vitreous has a passive role in accommodation, probably serving only as support for the lens.42 During ciliary muscle contraction, the choroid is pulled forward slightly, perhaps aiding in the correct orientation of the photoreceptors in relation to the entrance pupil. Contraction of the ciliary body in accommodation, by decreasing the circumference of the sclera, may lead to an elongation of the axial length of the eye.39 The ciliary muscle and trabecular meshwork are both attached to the scleral spur, and accommodation can cause a widening of the intertrabecular spaces, facilitating aqueous outflow and can result in decreased intraocular pressure (IOP).43
Clinical Comment: Presbyopia
The ability to focus at near distances decreases with age, and this loss in accommodative ability is called presbyopia. The objective measurement of accommodation nears zero by the age of 50, although subjective measurements of accommodative amplitude may be higher due to the depth of focus.44 The easiest response to patients when explaining presbyopia is that a “weakness of the muscle” causes this decrease in focusing ability. However, changes in the ciliary body, zonules, lens capsule, and the lens itself all influence the loss of accommodation; yet the precise nature of the impact each has is still unclear. Although ciliary muscle tissue is lost and replaced by connective tissue, this occurs in very old age, not when presbyopia begins.45 The force of ciliary muscle contraction does not decrease with age46; no loss of parasympathetic innervation occurs that would account for decreased muscle contraction and maximal contractile ability of the muscle decreases only slightly if at all with age.5 The diameter of the unaccommodated ciliary ring decreases in older eyes, thus the circumlental space between ciliary body and lens equator decreases with age, causing a decrease in zonular tension in the unaccommodated eye.47,48 There is some dispute as to whether the zonule-free area at the anterior lens surface decreases with age. The increase in anterior lens convexity and the increase in anterior lens capsule thickness might cause the appearance of what is called an anterior shift in the anterior zonule insertion on the capsule.48 There is no apparent increase in zonular length that presumably would accompany such an anterior shift.49 Some loss of fiber extensibility with age has been measured.31,49 The lens capsule becomes thicker, less elastic, and more brittle with age.50 Older lens fibers become more resistant to deformity, and thus the ability of the lens to change shape in response to the forces exerted by the capsule diminishes with age.10,51–54 As the lens continues to grow throughout life, the mass and volume increase, with a forward movement of the center of the lens. The increase in the bulk of the lens and the anterior displacement alter the vector relationship between the lens and the zonules, causing the zonular force to be more tangential to the lens surface and less able to change lens capsule tension.3,8 As the lens becomes more curved and little change occurs in lens power, greater force will be required to increase the power necessary for near focus.10,55
Clinical Comment: Slit-Lamp Appearance of Lens
An optic section through the lens demonstrates the biconvexity of the structure (Figure 5-10). The first bright line is convex forward and is the anterior lens capsule; posterior to this is a dark line, the anterior epithelium. The next bright line is the cortex, then various gray zones (zones of discontinuity) are seen. The anterior Y suture of the fetal nucleus may be evident. The vertical center, the embryonic nucleus, has no anterior or posterior curvature. The posterior inverted-Y suture may be seen in the posterior aspect of the fetal nucleus. Posterior to this, the zones of discontinuity are concave forward, with the final zone being the posterior capsule.56 The zones of discontinuity are apparent because of changes in light-scattering properties (Table 5-1). A diffuse view of the anterior lens surface illustrates lens shagreen, with the lens surface resembling the surface of an orange, likely caused by the conformation of the capsule to the epithelial cell undulations.57
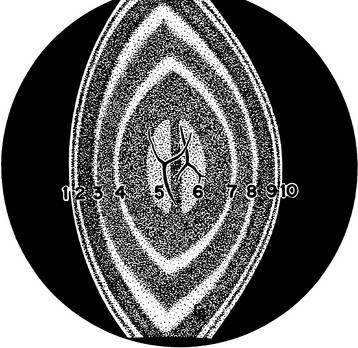
| Zones of Discontinuity | Correlative Lens Morphology |
|---|---|
| Anterior capsular zone | Anterior lens capsule |
| Anterior subcapsular zone | Anterior lens epithelium, elongating fibers, and outermost cortical fibers |
| Anterior zone of disjunction | Anterior segments of cortical fibers formed after sexual maturation, complex sutures |
| Anterior zone of the adult nucleus | Anterior half of the adult nucleus consisting of secondary fibers formed after birth to sexual maturation |
| Anterior zone of the fetal nucleus | Anterior half of the fetal nucleus consisting of secondary fibers formed before birth |
| Location of upright Y suture | |
| Zone of the embryonic nucleus | Embryonic nucleus consisting of primary lens fibers formed during first 2 months of embryonic development |
| Posterior zone of the fetal nucleus | Posterior half of the fetal nucleus consisting of secondary fibers formed before birth |
| Location of inverted Y suture | |
| Posterior zone of the adult nucleus | Posterior half of the adult nucleus consisting of secondary fibers formed after birth to sexual maturation |
| Posterior subcapsular zone | Posterior segments of elongating fibers and the outermost cortical fibers, complex sutures |
| Posterior capsular zone | Posterior lens capsule |
Adapted from Kuszak JR, Deutsch TA, Brown HG: Anatomy of aged and senile cataractous lenses. In Albert DM, Jakobiec FA, editors: Principles and practice of ophthalmology: basic sciences, Philadelphia, 1994, Saunders.
Lens Physiology
The primary function of the lens is the refraction of light, and it is imperative that the transparent lens have minimal light scatter. Transparency is a function of (1) the absence of blood vessels, (2) few cellular organelles in the light path, (3) an orderly arrangement of fibers, and (4) the short distance between components of differing indices relative to the wavelength of light.58 Since extensive metabolic activity occurs in the anterior epithelium maintaining cell and fiber function, and the preequatorial region has a high level of miotic activity, a significant amount of energy is used by these cells. Because the lens is avascular, most nutrients are obtained from the surrounding aqueous, with a small contribution from the vitreous. Thus the epithelium is rich in transport mechanisms (e.g., Na+/K+/ATPase pumps) that maintain electrolyte balance. Anaerobic glycolysis is the source of the energy required for cellular metabolism and cellular replication within the lens.
Lens Capsule
The lens capsule is first evident in early embryologic development and completely surrounds the early lens fibers. The lens is said to have immune-privilege and protection from infectious viruses and bacteria because the capsule sequesters the lens epithelium and fibers from early in prenatal development. Postnatally, the anterior lens epithelium and the posterior lens fibers continue to secrete and deposit matrix into the inner aspect of the capsule.59 As the lens itself grows throughout life, the capsule must expand as well, although the molecular mechanisms that regulate this are unknown. The capsule is permeable to water and small solutes and the proteins necessary for lens growth and function. Size and molecular charge may both influence passage through the capsule.59 A slow turnover of radio-labeled substances has been demonstrated within the capsule matrix (months to years), as compared with basement membranes elsewhere (hours).59 The capsule acts as a reservoir for the accumulation of molecules and growth factors that promote and regulate lens processes, such as proliferation, migration, and differentiation.59
Lens Fibers
Fiber Components
The lens is 65% to 70% water and 30% to 35% protein; the cortex has a higher water content (73% to 80%) than the nucleus (68%).60,61 The refractive index of the cortex is 1.38, and 1.41 is the index of the nucleus. An outer fiber can be 1 cm in length in the adult from suture to suture in the adult.62 Soluble lens proteins are known as crystallins, and insoluble proteins include those forming the cell membrane and the cytoskeleton.
The proteins manufactured during lens development must be durable because they need to last a lifetime. Eighty percent to 90% of the proteins within the lens are water soluble crystallins; this concentration is three times higher than in typical cells.62 Lens crystallins are from the alpha family or the beta/gamma super family. Interaction among crystallins, particularly the alpha crystallins, produce a phenomenon that contributes to lens transparency, and gives the lens a significantly higher index of refraction than surrounding fluids.63 Alpha crystallins are molecular chaperones and, as such, they stabilize beta and gamma proteins, preventing them from undergoing chemical changes and forming aggregates. When crystallins aggregate they undergo a change in density, become water insoluble, and when of sufficient size cause light scatter.64 Alpha crystallins also appear to be important in maintaining certain functions of lens cells and fibers. The beta/gamma crystallins are more diverse and their functions are unclear.63,64
Actin is an insoluble protein and an important component in the scaffolding of the lens fiber, its cytoskeleton. Microtubules are part of the cytoskeleton and help to stabilize the fiber membrane and may have a role in transporting vesicles to the ends of the elongating fibers.62 Numerous actin microfilaments, just inside the cell membrane, are linked to the adhesive junctions between lens fibers. Actin also helps to maintain crystallin organization.66 Lens fiber membranes have the highest cholesterol content of human cells and a high concentration of sphingomyelin. The function of sphingomyelin is unclear since it can cause rigidity in membranes and lens fibers must exhibit flexibility.62
Formation of Lens Fibers
Lens fiber formation is a complex and multistep process and various molecules influence the mechanism. Growth factors, present in the aqueous and the vitreous, accumulate in the lens capsule. The concentration and the distribution of specific factors along the lens surface direct cellular processes.65 Growth factors that influence proliferation and migration are concentrated along the anterior surface; other growth factors that influence differentiation are concentrated at the equator.66 Biomolecules that regulate interactions among actin filaments, adhering junction integrins, and extracellular matrix increase fiber mass.67 Significant protein synthesis must occur to form crystallins, aquaporin channel proteins, and gap junction components as the fibers elongate.67 As the fiber cell elongates, the cell membrane permeability increases, causing K+ and Cl− accumulation in the cytoplasm, driving water entrance and cell volume increase.68
As the cell elongates, the apical aspect slides along the apical aspect of the anterior epithelium, and the basal aspect slides along the posterior capsule. Once the elongating end reaches the end of an elongating fiber from the opposite side of the lens, they join, forming a suture. The basal end detaches from the capsule and once this detachment occurs, the membrane-bound organelles (nucleus, endoplasmic reticulum, mitochondria) degrade in an apoptosis-like process; the loss of organelles is complete within a few hours.67,69,70
Fiber Junctions
Although mature lens fibers lack cellular organelles, they still require nutrients. The fibers deep within the lens are far from the aqueous and vitreous, and fiber-to-fiber transport is important. An intracellular network of gap junctions facilitates movement of ions and small molecules between fibers.71 The lens has a higher concentration of gap junctions than other cells in the body; the lens gap junctions contain; some channel proteins that are unique to the lens.13,62
Lens Metabolism
The lens obtains glucose from the aqueous humor and because of the low oxygen concentration in the neighborhood of the lens, 70% of ATP production is via anaerobic metabolism. Aerobic glycolysis and the Krebs cycle are limited to the epithelium or superficial fibers that still have mitochondria. The thickness of the lens cortex, in which newer fibers are present and which still contains organelles, is approximately 100 microns.62 ATP activity is higher in the epithelial cells and the newer fibers of the cortex near the equator and is lower near the poles. There is no such activity in the lens nucleus, and the fibers in the nucleus are not capable of protein synthesis.69
Ionic Current
An ionic current has been identified flowing out of the lens at the equator and into the lens at the poles (Figure 5-11).24,69 It is likely that ATPase activity contributes to this current because the distribution of ATPase pumps is coincident with this pattern.71 The Na+K+/ATPase activity generates an electrochemical gradient with the interior of the lens more negative than its surrounding environment. This circulating ionic flow might help circulate solutes to the deep lens fibers and transport wastes out of the fibers and out of the lens.68 The fluid would follow the same pathway as the ionic current, facilitating water and metabolite (glucose, ascorbate, and amino acids) movement into the deeper fibers.24 Water and solutes enter the lens through extracellular spaces at the anterior and posterior polar regions, cross fiber membranes to the lens interior, and then flow through fibers back to the surface at the equator, matching the distribution of the ionic pumps and channels.68
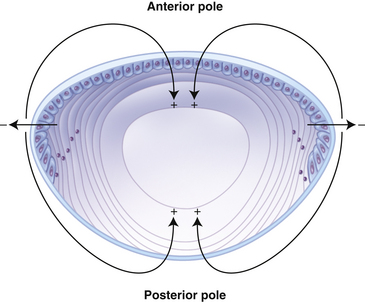
Regulation of Fluid Volume
Cl− flux appears to be the important factor in regulating fluid volume.68 The Na+K+/2Cl− and K+/2Cl− cotransporters and Cl− channels maintain ion concentration gradients at a level that keeps water in equilibrium across the cell and fiber membranes.24,68 The membranes of the epithelium and the fibers are highly permeable and aquaporins are numerous, enhancing water movement into and out of the lens.72
Ultraviolet Radiation (UVR)
The cornea absorbs wavelengths below 300 nm, the lens absorbs wavelengths between 300 and 400 nm, and wavelengths greater than 400 nm are transmitted to the retina. The lens absorbs almost all UVR to which it is exposed, and any resulting unstable free radicals cause molecular changes.73 The first active tissue of the lens that encounters UVR is the lens epithelium, which is susceptible to damage from free radicals. Morphologic changes apparent in the epithelial layer may lead to irreversible changes throughout the lens, although the mechanism by which this progresses has not been defined.74
UVR absorbed by lens fibers causes oxidative damage, leading to degradation and modification of lens proteins. An association exists between ocular exposure and increased risk of lens opacity.75,76 UVR absorption also increases chromophore concentration; yellow pigments accumulate in the center of the lens.73 The yellowing may progress to a dark-brown hue, which is called lens brunescence.25
Oxidative Stress
Free radicals are generated both by UVR absorption and by cellular metabolic processes. Oxidative stress results when the rate of free radical production is greater than the rate of their degradation. Oxidative stress can impair the structure and function of connexins (gap junction proteins), can modify lens crystallins, and can cause aggregation of proteins, all of which contribute to cataract development.77 Glutathione is a reducing agent that detoxifies free radicals, thus preventing such damage. It is found in high concentration within the lens and the aqueous humor and is transported into the lens from the aqueous. It can be synthesized and regenerated by the lens epithelial cells and young lens fibers.68 The deeper fibers rely on diffusion of glutathione from superficial fibers.78 Glutathione also has a role in maintaining membrane transport mechanisms.78
Ascorbic acid also provides some protection to lens epithelium against UV-induced damage to deoxyribonucleic acid (DNA) and is present in relatively high levels in the aqueous humor.79
Aging Changes in Crystalline Lens
The lens grows throughout life. The majority of the increase in thickness occurs between ages 8 and 40, accompanied by an increase in surface curvatures, a forward movement of the center of the lens, and a decrease in anterior chamber depth.2,35,53,80 Other physical changes that accompany age were described in the section about presbyopia. Changes occur in lens physiology as mature lens fibers lose all cellular organelles. A coincident decrease in the transport of ions, nutrients, and antioxidants may lead to damage contributing to cataract formation.81 With age there is an increase in fiber membrane permeability and the ionic pumps may not be able to compensate, disrupting ion balance. Circulation within the lens changes and restriction of the flow of water and of glutathione occurs at the cortex/nucleus border. Significant changes in aquaporins occur, causing a disruption of water flow.82
The amount of water soluble alpha crystallins decreases with age, and by age 45 there are no alpha crystallins evident in the lens nucleus. Since the alpha crystallins help to prevent other crystallins from forming aggregates, water insoluble aggregates increase with age.77 Some components of the cytoskeleton disassemble.83 Levels of UVR filters in the lens decrease approximately 12% per decade, allowing increased UVR damage.63
Clinical manifestations of aging are presbyopia and cataract formation. Both processes affect vision and are a significant concern to the patient and to the clinician, particularly because few preventive measures are available. Recommendations to patients should include the use of UVR-absorbing lenses when outdoors; the incidence of cataract is higher in those exposed to greater levels of sunlight.73
Clinical Comment: The Lens Paradox
Because the lens continues to grow, it would seem that its refractive power should change, yet it remains constant. With age the radii of curvature decrease; the anterior radius of curvature decreases to approximately 8.25 μm and the posterior radius to about 7 μm by 80 years of age.55 As the lens surface becomes more steeply curved, refractive power should increase. However, the lens thickness increases primarily in the width of the lens cortex, and as the lens becomes more optically homogenous, there is less of an affect from the gradient nature of the index of refraction; these changes apparently compensate for the increased surface curvatures, and the power of the lens remains stable.84–87
Cataracts
Clinical Comment: Clinical Cataract
Although any lens opacity is accurately called a cataract, the clinician should be aware of the impact that the word “cataract” may have on a patient. Cataracts are the greatest cause of blindness worldwide. The etiology of cataract formation is complex, and cataract development is often the result of multiple factors that influence lens metabolism. Risk factors include aging, disease, genetics, nutritional or metabolic deficiencies, trauma, congenital factors, and environmental stress (e.g., radiation), with age being the major contributor.88
Cataracts are named according to location or cause and can be graded based on severity (Figure 5-12). An opacity located in the embryonic, fetal, or adult nucleus is called a nuclear cataract (Figure 5-13). The center opacification accompanying the onset of a nuclear cataract can increase refractive power, and in the hyperopic patient this myopic shift causes a temporary improvement in vision. Brunescence accompanies nuclear cataracts caused by increased chromophore concentration. The increase in yellow coloration results in the absorption of wavelengths in the blue end of the spectrum, which may actually provide some protection for the macula.
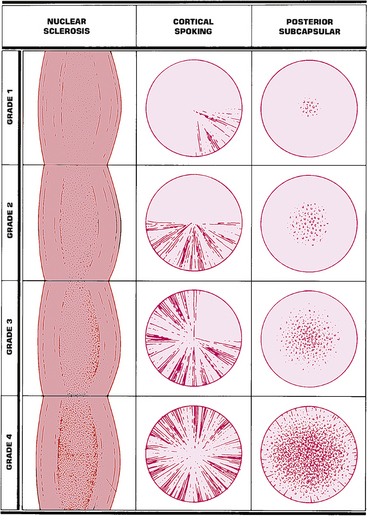
FIGURE 5-12 Grading system for age-related cataracts.
(From Fingeret M, Casser L, Woodcome HT: Atlas of primary eyecare procedures, Norwalk, Conn, 1992, Appleton & Lange.)
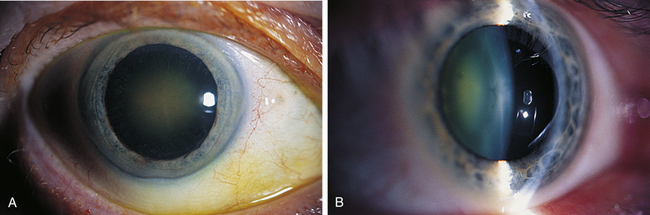
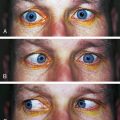
FIGURE 5-13 Nuclear cataract seen with A, diffuse illumination, and B, optic section.
(From Kanski JJ, Nischal KK: Ophthalmology: clinical signs and differential diagnosis, St Louis, 2000, Mosby.)
A cortical cataract, located in the cortex, has a spokelike shape; thicker in the periphery and tapering toward the lens center, it follows the shape of the fiber (Figure 5-14). Cortical cataracts generally progress slowly; with time the spoke width expands as the opacity spreads to adjacent fibers.89,90 Fluid accumulates and membrane rupture in the equatorial area can occur.91 Cortical cataracts affect vision only when they spread into the center of the lens and cause light scatter in the pupillary region.
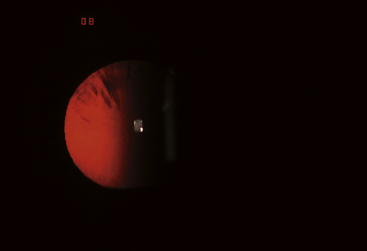
FIGURE 5-14 Spokes of a cortical cataract visible against the red reflex.
(Courtesy Pacific University Family Vision Center, Forest Grove, Ore.)
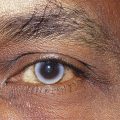
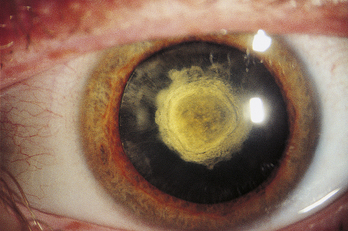
A posterior subcapsular cataract is a disturbance located just beneath the posterior capsule (Figure 5-15). This type of cataract impacts vision early and significantly given its location along the visual axis and near the nodal point of the eye. A significant risk factor for posterior subcapsular cataracts is long-term, high-dose steroid use.
The Physiology of Cataract Formation
Numerous mechanisms are presumed causative for cataracts, including fluid and ion imbalance, oxidative damage, protein modification, and metabolic disruption.66 A disturbance in fluid regulation can be caused by ionic pump dysfunction and/or membrane permeability increase that allows water accumulation. If Na+/K+ ATPase pump activity decreases significantly, a rise in Na+ in the cytoplasm is accompanied by an influx of water, the lens fibers swell and transparency diminishes.69 An increased level of cytoplasmic Ca++ is also associated with a loss of transparency.69 Water accumulation between fibers can form vacuoles causing a disruption of fiber arrangement and increased light scatter. UVR and oxidative damage as a result of free radical accumulation affects cellular function, damages lens DNA, causes protein modification, and high-molecular-weight crystallin aggregations, any of which can increase light scatter.25 Alpha crystallins, as molecular chaperones, help to stabilize beta/gamma crystallin configuration but by age 40 have disappeared from the lens nucleus, although the normal lens usually remains fairly transparent for years past that age.92 But as the concentration of alpha crystallins is reduced, aggregates accumulate and with time form light-scattering opacities.
Glutathione and ascorbate maintain a reducing environment providing some protection from free radical damage and preventing protein modification. Reduced levels of glutathione allow oxidative damage to membranes and proteins.77 A decrease in glutathione concentration is associated with cataract development.92,93 A barrier, speculated to develop in middle age and located at the interface of the cortex and nucleus, seems to impede the flow of small molecules from the cortex into the nucleus and might account for the reduction in glutathione in the nucleus.91 A modification of the connexins in gap junctions causes a disruption in communication between fibers and might be one cause of this barrier.94,95 Changes occur in aquaporin channel proteins in the innermost nuclear regions of the lens as early as age 5 and by middle age (age 40 to 50), half of such channels are lost in the region of the speculated barrier.82 These changes can lead to the occlusion of the water channels and contribute to the barrier function.
A diabetic cataract results from elevated glucose levels and can develop rapidly. With increased blood glucose, excess glucose present in the aqueous enters the lens. As this excess glucose is metabolized, sorbitol accumulates faster than it is converted to fructose. Sorbitol concentration increases within the lens fiber, because sorbitol does not readily pass through the fiber membrane, and thus water is drawn into the fiber. The fibers swell, the lens loses transparency, and the fibers may eventually rupture.
Age-Related Cortical Cataract
High lifetime exposure to UVR is associated with increased incidence of cortical cataracts; the paradox is that the most severe damage in cortical cataracts occurs near the equator initially, the area most protected from sunlight by the iris. Cortical cataracts are associated with increased membrane permeability and ion transporters, pumps, and exchangers are not able to maintain the homeostatic concentration.71 An increased concentration of Ca++ in the fiber cytoplasm also drives fluid accumulation.69 Affected regions of the fiber show disruption of structure and can include membrane rupture. The changes first occur in the center of the elongated fiber (that is at the equatorial region), with the apical and basal ends remaining transparent. Generally the tapered fiber ends, located at the sutures in the optical axis, are only affected very late in the life of the cortical cataract.
Age-Related Nuclear Cataract
Age-related nuclear cataracts are associated with a decline of glutathione, making the fibers susceptible to oxidative damage. Levels can be significantly reduced in the nucleus while levels in the cortex remain within the normal range.78,94 Oxidative protein modification increases significantly after age 50, contributing to the damage seen in senile nuclear sclerosis.92 The color changes that often accompany nuclear cataracts are usually seen as various hues of yellow or brown; this pigmentation is primarily protein-bound.91
Posterior Subcapsular Cataract (PSC)
The opacity in the posterior subcapsular region is formed by epithelial-like cells that migrate from the equatorial region. These cells accumulate at the posterior pole forming an opacity.96 It is speculated that radiation damage is one causative factor as patients undergoing radiation therapy for cancer treatments also develop PSCs and/or cortical cataracts.62 There are some sutural congenital cataracts that occur at this location, but they are caused by swelling of the basal ends of cortical fibers.62
Steroid-Induced Cataract
Steroid-induced cataracts are also located in the posterior subcapsular region. Dosage and the duration of steroid use appear to be controlling factors, although individuals may have varying levels of susceptibility. Children develop such cataracts at a faster rate than do adults. Reversal of the cataract can occur but this is rare.5,37,66 The opacity appears to be formed of undifferentiated epithelial cells at the interface of the posterior cortex and capsule. These misplaced cells (which should only be present in the lens epithelium) display aberrant behavior. The undifferentiated cells may have migrated from the preequatorial area, influenced by a change in the concentration of growth factors.66 The growth factors governing mitosis, migration, and differentiation are obtained from aqueous and reside in the lens capsule. If steroids influence production of these growth factors that are present in the aqueous, and the concentration and location in the capsule is altered, cellular processes can be affected.66
1. Patterson C.A., Delamere N.A. The lens. In: Hart W.M., editor. Adler’s physiology of the eye. ed 9. St Louis: Mosby; 1992:348.
2. Dubbelman M., van der Heijde G.L., Weeber H.A. The thickness of the aging human lens obtained from corrected Scheimpflug images. Optom Vis Sci. 2001;78(6):411.
3. Koretz J.E., Strenk S.A., Strenk L.M., et al. Scheimpflug and high resolution magnetic resonance imaging of the anterior segment: a comparative study. J Opt Soc Am A Opt Image Sci Vis. 2004;21:346-354.
4. Hogan M.J., Alvarado J.A., Weddell J.E. Histology of the human eye. Philadelphia: Saunders; 1971. p 638
5. Strenk S.A., Semmlow J.L., Strenk L.M., et al. Age-related changes in ciliary muscle and lens: a magnetic resonance imaging study. Invest Ophthalmol Vis Sci. 1999;40:1162.
6. Jones C.E., Atchison D.A., Pope J.M. Changes in lens dimensions and refractive index with age and accommodation. Optom Vis Sci. 2007;84:990-995.
7. Rosen A.M., Denham D.B., Fernandez V., et al. in vitro dimensions and curvatures of human lenses. Vision Res. 2006;46:1002-1009.
8. Glasser A., Kaufman P.L. Accommodation and presbyopia. In: Kaufman P.L., Alm A., editors. Adler’s physiology of the eye. ed 10. St Louis: Mosby; 2003:197.
9. Smith G., Pierscionek B.K. The optical structure of the lens and its contribution to the refractive status of the eye. Ophthalmic Physiol Opt. 1998;18(1):21.
10. Pierscionek B.K. Refractive index contours in the human lens. Exp Eye Res. 1997;64:887.
11. Borish I.M. Clinical refraction, ed 3. Chicago: Professional Press; 1975. p 169
12. Koretz J.F., Handelman G.H., Brown N.P. Analysis of human crystalline lens curvature as a function of accommodative state and age. Vision Res. 1984;24:1141.
13. Kuszak J.R., Brown H.G. Embryology and anatomy of the lens. In: Albert D.M., Jakobiec F.A., editors. Principles and practice of ophthalmology. Philadelphia: Saunders; 1994:82.
14. Barraquer R.I., Michael R., Abreu R., et al. Human lens capsule thickness as a function of age and location along the sagittal lens perimeter. Invest Ophthalmol Vis Sci. 2006;47:2053-2060.
15. Alexander R.A., Garner A. Elastic and precursor fibres in the normal human eye. Exp Eye Res. 1983;36:305.
16. Krag S., Olsen T., Andreassen T.T. Biomechanical characteristics of the human anterior lens capsule in relation to age. Invest Ophthalmol Vis Sci. 1997;38(2):357.
17. Kuwabara T. The maturation of the lens cell: a morphologic study. Exp Eye Res. 1975;20:427.
18. Kuszak J.R., Peterson K.L., Brown H.G. Electron microscopic observations of the crystalline lens. Microsc Res Tech. 1996;33:441.
19. Bassnett S., Kuszak J.R., Reinisch L., et al. Intercellular communication between epithelial and fiber cells of the eye lens. J Cell Sci. 1994;107:799.
20. Rae J. Physiology of the lens. In: Albert D.M., Jakobiec F.A., editors. Principles and practice of ophthalmology. Philadelphia: Saunders; 1994:123.
21. Kuszak J.R., Novak L.A., Brown H.G. An ultrastructural analysis of the epithelial-fiber interface (EFI) in primate lenses. Exp Eye Res. 1995;61:579.
22. Lo W.K., Harding C.V. Structure and distribution of gap junctions in lens epithelium and fiber cells. Cell Tissue Res. 1986;244(2):253.
23. Lo W.K., Harding C.V. Tight junctions in the lens epithelia of human and frog: freeze-fracture and protein tracer studies. Invest Ophthalmol Vis Sci. 1983;24(4):396.
24. Mathias R.T., Kistler J., Donaldson P. The lens circulation. J Membr Biol. 2007;216:1-16.
25. Vavvas D., Azar N.F., Azar D.T. Mechanisms of disease: cataracts. Ophthalmol Clin North Am. 2002;15:49.
26. Clark J.I. Development and maintenance of lens transparency. In: Albert D.M., Jakobiec F.A., editors. Principles and practice of ophthalmology. Philadelphia: Saunders; 1994:114.
27. Mathias R.T., Rae J.L. Transport properties of the lens. Am J Physiol. 1985;249(3):181.
28. Dahm R., van Marle J., Prescott A.R., et al. Gap junctions containing alpha8-connexin (MP70) in the adult mammalian lens epithelium suggests a re-evaluation of its role in the lens. Exp Eye Res. 1999;69:45.
29. Kuszak J.R., Peterson K.L., Sivak J.G., et al. The interrelationship of lens anatomy and optical quality. II. Primate lenses. Exp Eye Res. 1994;59(5):521.
30. Streeten B.W., Licari P.A. The zonules and the elastic microfibrillar system in the ciliary body. Invest Ophthalmol Vis Sci. 1983;24(6):667.
31. Bourge J.L., Robert A.M., Renard G. Zonular fibers, multimolecular composition as related to function (elasticity) and pathology. Pathol Biol (Paris). 2007;55:347-359.
32. Rohen J.W. Scanning electron microscopic studies of the zonular apparatus in human and monkey eyes. Invest Ophthalmol Vis Sci. 1979;18:133.
33. Von Helmholtz HH: Treatise on physiologic optics, Mineola, NY, 1962, Dover (Translated by JPC Southhall), p 143.
34. Von Noorden G. Burien and von Noorden’s binocular vision and ocular motility, ed 5. St Louis: Mosby; 1996. p 85
35. Johnson C.A. Effects of luminance and stimulus distance on accommodation and visual resolution. J Opt Soc Am. 1976;66:138-142.
36. Garner L.F., Smith G. Changes in equivalent and gradient refractive index of the crystalline lens with accommodation. Optom Vis Sci. 1997;74(2):114.
37. Garner L.F., Yap M.K. Changes in ocular dimensions and refraction with accommodation. Ophthalmic Physiol Opt. 1997;17(1):12.
38. Drexler W., Baumgartner A., Findl O., et al. Biometric investigation of changes in the anterior eye segment during accommodation. Vision Res. 1997;37:2789.
39. Drexler W., Findl O., Schmetterer L., et al. Eye elongation during accommodation in humans: differences between emmetropes and myopes. Invest Ophthalmol Vis Sci. 1998;39(11):2140.
40. Koretz J.F., Bertasso A.M., Neider M.W., et al. Slit-lamp studies of the rhesus monkey eye. II. Changes in crystalline lens shape, thickness and position during accommodation and aging. Exp Eye Res. 1987;45:317.
41. Ludwig K., Wegscheider E., Hoops J.P., et al. in vivo imaging of the human zonular apparatus with high-resolution ultrasound biomicroscopy. Graefes Arch Clin Exp Ophthalmol. 1999;237:361-371.
42. Fisher R.F. The vitreous and lens in accommodation. Trans Ophthalmol Soc U K. 1982;102:318.
43. Mauger R.R., Likens C.P., Applebaum M. Effects of accommodation and repeated applanation tonometry on intraocular pressure. Am J Optom Physiol Opt. 1984;6(1):28.
44. Hamasaki D., Ong J., Marg E. The amplitude of accommodation in presbyopia. Am J Optom Arch Am Acad Optom. 1956;33:3-14.
45. Pardue M.T., Sivak J.G. Age-related changes in human ciliary muscle. Optom Vis Sci. 2000;77:204.
46. Hermans E.A., Dubbelman M., van der Heijde G.L., et al. Change in the accommodative force on the lens of the human eye with age. Vision Res. 2008;48:119-126.
47. Strenk S.A., Strenk L.M., Guo S. Magnetic resonance imaging of aging, accommodating, phakic, and pseudophakic ciliary muscle diameters. J Cataract Refract Surg. 2006;32:1792-1798.
48. Sakabe I., Oshika T., Lim S.J., et al. Anterior shift of zonular insertion onto the anterior surface of human crystalline lens with age. Ophthalmology. 1998;105(2):295.
49. Assia E.I., Apple D.J., Morgan R.C., et al. The relationship between stretching capability of the anterior capsule and zonules. Invest Ophthalmol Vis Sci. 1991;32:2835-2839.
50. Fisher R.F. The influence of age on some ocular basement membranes. Eye. 1987;1:184-189.
51. Krag S., Andreassen T.T. Mechanical properties of the human posterior lens capsule. Invest Ophthalmol Vis Sci. 2003;44:691.
52. Seland J.H. Ultrastructural changes in the normal human lens capsule from birth to old age. Acta Ophthalmol. 1974;52:688.
53. Glasser A., Campbell M.C. Biometric, optical and physical changes in the isolated human crystalline lens with age in relation to presbyopia. Vision Res. 1999;39:1991.
54. Beers A.P., van der Heijde G.L. Age-related changes in the accommodation mechanism. Optom Vis Sci. 1996;73(4):235.
55. Brown N. The change in lens curvature with age. Exp Eye Res. 1974;19:175.
56. Brandreth R.H. Clinical slit lamp biomicroscopy. Berkeley, Calif: Brandreth; 1978.
57. Sasaki K., Kojirna M., Hara T. in vivo observation of the crystalline lens capsule. Ophthalmic Res. 1988;20(3):154.
58. Trokel S. The physical basis for transparency of the crystalline lens. Invest Ophthalmol. 1962;1:493.
59. Danysh B.P., Duncan M.K. The lens capsule. Exp Eye Res. 2009;88:151-164.
60. Hejtmancik J.F. Congenital cataracts and their molecular genetics. Semin Cell Dev Biol. 2008;19:134-149.
61. Chong H.N.V. Clinical ocular physiology, Butterworth Heinemann. In Linacre House. Oxford, UK: Jordan Hill; 1996. p 41
62. Beebe D.C. The lens. In: Kaufman P.L., Alm A., editors. Adler’s physiology of the eye. ed 10. St Louis: Mosby; 2003:117.
63. Truscott R.J. Presbyopia. Emerging from a blur towards an understanding of the molecular basis for this most common eye condition. Exp Eye Res. 2009;88:241-247.
64. Takemoto L., Sorensen C.M. Protein-protein interactions and lens transparency. Exp Eye Res. 2008;87:496-501.
65. Zelenka P.S., Arpitha P. Coordinating cell proliferation and migration in the lens and cornea. Sem Cell Develop Bio. 2008;19:113-124.
66. Jobling A.I., Augusteyn R.C. What induces steroid cataracts? A review of steroid-induced posterior subcapsular cataracts. Clin Exp Optom. 2002;85:61-75.
67. Rao P.V., Maddala R. The role of the lens actin cytoskeleton in fiber cell elongation and differentiation. Sem Cell Dev Bio. 2006;17:698-711.
68. Donaldson P.J., Chee K.S., Lim J.C., et al. Regulation of lens volume: Implication for lens transparency. Exp Eye Res. 2009;88:144-150.
69. Delamere N.A., Tamiya S. Lens Na+, K+-ATPase. In: Tombran-Tink J., Barnstable C.J., editors. Ophthalmology research: ocular transporters in ophthalmic diseases and drug delivery. Totowa, NJ: Humana Press; 2008:111-123.
70. Bassnett S., Beebe D.C. Coincident loss of mitochondria and nuclei during lens fiber cell differentiation. Dev Dyn. 1992;194:85-92.
71. Delamere N.A., Tamiya S. Lens ion transport: from basic concepts to regulation of Na+, K-ATPase activity. Exp Eye Res. 2009;88:140-143.
72. Zampighi G.A. The lens. In: Fischbarg J., editor. The biology of the eye. Amsterdam: Elsevier; 2006:149-179.
73. Young R.W. The family of sunlight-related eye diseases. Optom Vis Sci. 1994;71(2):125.
74. Hightower K.R., Reddan J.R., McCready J.P., et al. Lens epithelium: a primary target of UVB irradiation. Exp Eye Res. 1994;59:557.
75. Hightower K.R. The role of the lens epithelium in development of UV cataract. Curr Eye Res. 1995;14:71.
76. West S.K., Duncan D.D., Muñoz B., et al. Sunlight exposure and risk of lens opacities in a population-based study. Arch Ophthalmol. 1998;116:1666.
77. Berthoud V.M., Beyer E.C. Oxidative stress, lens gap junctions, and cataracts. Antioxid Redox Signal. 2009;11:339-353.
78. Reddy V.N. Glutathione and its function in the lens—an overview. Exp Eye Res. 1990;50:771-778.
79. Reddy V.N., Giblin F.J., Lin L.R., et al. The effect of aqueous humor ascorbate on ultraviolet-B-induced DNA damage in lens epithelium. Invest Ophthalmol Vis Sci. 1998;39(2):344.
80. Alió J.L., Schimchak P., Negri H.P., et al. Crystalline lens optical dysfunction through aging. Ophthalmology. 2005;112:2022-2029.
81. Moffat B.A., Landman K.A., Truscott R.J., et al. Age-related changes in the kinetics of water transport in normal human lenses. Exp Eye Res. 1999;69(6):663.
82. Korlimbinis A., Berry Y., Thibault D., et al. Protein aging: truncation of aquaporin 0 in human lens regions is a continuous age-dependent process. Exp Eye Res. 2009;88:966-973.
83. Friedrich M.G., Truscott R.J. Membrane association of proteins in the aging human lens: Profound changes take place in the fifth decade of life. Invest Ophthalmol Vis Sci. 2009;50:4786-4793.
84. Pierscionek B.K. Age-related response of human lenses to stretching forces. Exp Eye Res. 1995;60:325.
85. Moffat B.A., Atchison D.A., Pope J.M. Age-related changes in refractive index distribution and power of the human lens as measured by magnetic resonance micro-imaging in vitro. Vis Res. 2002;42:1683.
86. Dubbelman M., van der Heijde G.L. The shape of the aging human lens: curvature, equivalent refractive index and the lens paradox. Vision Res. 2001;41:1867.
87. Charman W.N. The eye in focus: accommodation and presbyopia. Clin Exp Optom. 2008;91:207-225.
88. Cruickshanks K.J., Klein B.E., Klein R. Ultraviolet light exposure and lens opacities: the Beaver Dam Eye Study. Am J Public Health. 1992;82(12):1658.
89. Brown N.P., Harris M.L., Shun-Shin G.A., et al. Is cortical spoke cataract due to lens fibre breaks? The relationship between fibre folds, fibre breaks, waterclefts and spoke cataract. Eye. 1993;7:672.
90. Vrensen G., Willekens B. Biomicroscopy and scanning electron microscopy of early opacities in the aging human lens. Invest Ophthalmol Vis Sci. 1990;31(8):1582.
91. Truscott R.J. Age-related nuclear cataract-oxidation is the key. Exp Eye Res. 2005;80:709-725.
92. Sweeney M.H., Truscott R.J. An impediment to glutathione diffusion in older normal human lenses: a possible precondition for nuclear cataract. Exp Eye Res. 1998;67:587.
93. Truscott R.J. Age-related nuclear cataract: a lens transport problem. Ophthalmol Res. 2000;32:185.
94. Sweeney M.H., Truscott R.J. An impediment to glutathione diffusion in older normal human lenses: a possible precondition for nuclear cataract. Exp Eye Res. 1998;67:587-595.
95. Moffat B.A., Landman K.A., Truscott R.J., et al. Age-related changes in the kinetics of water transport in normal human lenses. Exp Eye Res. 1999;69:663-669.
96. Streeten B.W., Eshaghian J. Human posterior subcapsular cataract. A gross and flat preparation study. Arch Ophthalmol. 1978;96:1653-1658.