CHAPTER 50 Wrist and hand
SKIN AND SOFT TISSUE
SKIN
Skin creases and fingerprints
Flexure lines commonly crease the skin across the flexor surfaces of the wrist and hand (Fig. 50.1). Though not all directly over their functionally related subjacent skeletal joints, they are produced by adhesion of the skin to subjacent deep fascia and are sites of folding of the skin during movement. These flexures are useful landmarks. Less regular, but quite prominent, crease-line complexes are centred over the dorsal (extensor) aspects of the radiocarpal, carpal, metacarpophalangeal and interphalangeal joints. They are mainly transverse but display varying curvatures. During flexion the dorsal skin is stretched and the lines become less prominent (but can still be identified). During extension the now redundant skin becomes increasingly puckered and the lines are finally maximally prominent. (For a general review of ‘skin lines’ see p. 160.)
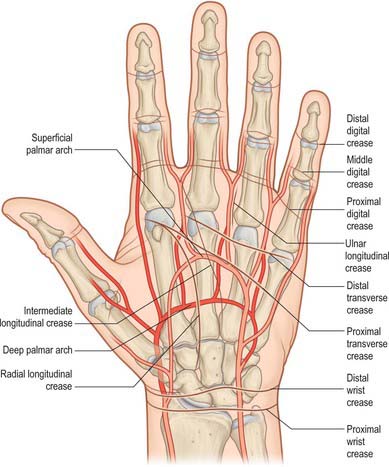
Fig. 50.1 Relation of the skin flexure lines and palmar arterial arches to the bones of the left hand.
(Adapted from Drake, Vogl and Mitchell 2005.)
The second to fifth digits show proximal, middle and distal sets of transverse lines. The proximal, often double, are at the digital roots, approximately 2 cm distal to the metacarpophalangeal joints. The middle lines are typically double, the proximal line lying directly over the proximal interphalangeal joint. The distal lines are usually single, and lie proximal to the distal interphalangeal joints: their levels are sometimes marked by a fainter, more distal line. The free pollicial base is partly encircled by a line which starts on the radial side and crosses distally over the metacarpophalangeal joint to end between the thumb and index finger level with the base of the proximal pollicial phalanx. There is a second, shorter crease usually 1 cm distal to this line. There are two lines comparable to the middle digital lines in other digits opposite the interphalangeal joint of the thumb (see also p. 160).
Cutaneous vascular supply
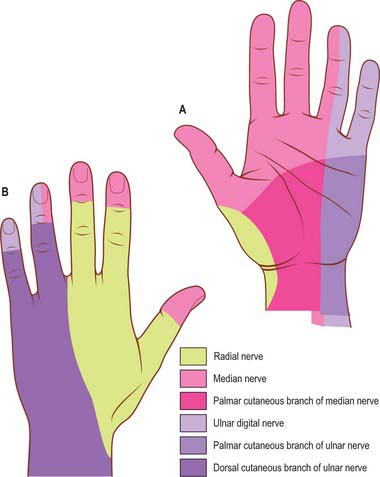
The skin of the volar aspect of the wrist is supplied directly by cutaneous branches from the superficial palmar branch of the radial artery, the ulnar artery and occasionally the median artery if it is large enough (Fig. 50.2). The skin over the thenar eminence is supplied by small perforating branches from the superficial palmar branch of the radial artery and the princeps pollicis. The skin over the hypothenar eminence is supplied by perforating branches from the ulnar artery, some of which pass through palmaris brevis. The remainder of the palm is supplied by small perforating branches from the common palmar digital arteries which pierce the palmar aponeurosis, and small branches from the radialis indicis artery. The blood supply to the volar aspect of the digital skin comes from small branches from each digital artery. At the level of the distal phalanx the two digital arteries typically form an H-shaped anastomosis from which cutaneous perforators fan out within the pulp. Deep digital veins accompanying the digital arteries are usually very small and frequently absent. More commonly, superficial palmar veins tend to pass dorsally and drain into the larger superficial dorsal venous system.
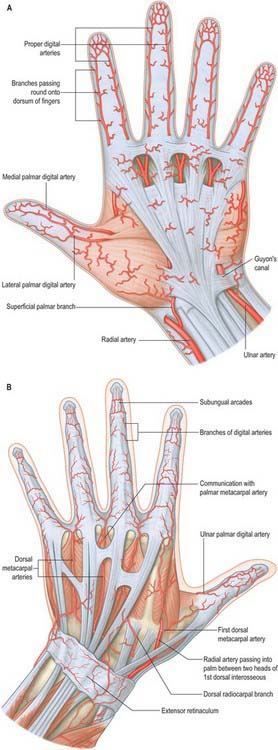
The blood supply to the dorsum of the fingers comes proximally from the terminal branches of the dorsal metacarpal arteries – supplying a region as far distally as the proximal interphalangeal joint – as well as from dorsal branches of the palmar digital arteries which are given off at each phalangeal level. At the level of the distal phalanx the cutaneous supply comes from three dorsal arcades: a superficial arcade over the base of the distal phalanx, and two distal subungual arcades. The skin of the dorsum of the thumb is supplied by longitudinal axial branches of the princeps pollicis and dorsal branches from the palmar digital arteries.
Cutaneous innervation
The skin of the volar aspect of the wrist is innervated by the terminal branches of the lateral and medial cutaneous nerves of the forearm. The skin of the palm is innervated by the palmar branches of the ulnar nerve and the palmar branch of the median nerve (Fig. 50.3; see also Fig. 50.45. The skin of the volar aspect of the thumb, index, middle and radial aspect of the ring fingers is supplied by cutaneous branches of the median nerve, while that of the little finger and ulnar side of the ring finger is supplied by the ulnar nerve.
NAIL APPARATUS
The nail apparatus consists of the nail plate, proximal and lateral nail folds, nail matrix, nail bed and hyponychium. It is described on page 154.
SOFT TISSUE
Palmar fascial complex
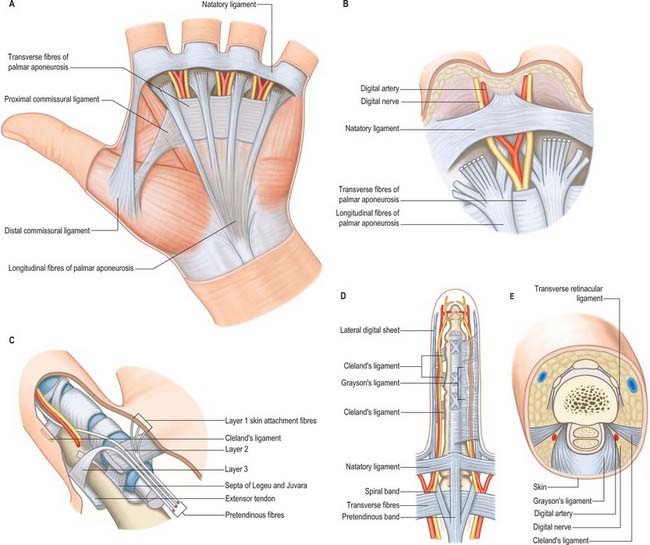
The palmar fascia is a three-dimensional ligamentous system composed of longitudinal, transverse and vertical fibres (Fig. 50.4).
Longitudinal fibre system
The longitudinal fibres represent the phylogenetically degenerated metacarpophalangeal joint flexor. They run distally from the palmaris longus tendon or the flexor retinaculum of the wrist across the whole width of the central third of the palm, producing four well-defined longitudinal bundles to the index, middle, ring and little fingers. A less well-defined bundle passes to the thumb. Distal to the transverse fibres of the palmar aponeurosis the longitudinal fibres pass in three layers (McGrouther 1982). The most superficial longitudinal fibres (layer 1) are inserted superficially into the skin of the distal palm between the distal palmar crease and the proximal digital crease. Some superficial fibres pass distally into the palmar midline of the digit. Deeper longitudinal fibres (layer 2) pass deep to the natatory ligament and neurovascular bundles into the apex of the web space skin and into the fingers themselves where they are continuous with Cleland’s ligaments and the lateral digital sheet. These are known as the spiral bands of Gosset. Deeper still, the longitudinal fibres in layer 3 perforate the deep transverse metacarpal ligament to pass around the sides of the metacarpophalangeal joint and attach to the metacarpal bone and proximal phalanx, and extensor tendon.
Transverse fibre system
Digital fascial complex
The superficial fascia within the finger is fibrofatty in the palmar and dorsal aspects, but more sheet-like laterally, where it is termed the lateral digital sheet. Within the core of the finger the fascia is thickened in areas, forming the flexor sheath, Cleland’s, Grayson’s and Landsmeer’s ligaments (Fig. 50.4). The flexor sheath is discussed in detail on page 879. Cleland’s ligaments extend from the sides of the phalanges, pass dorsal to the neurovascular bundles and insert into the lateral digital sheet. Grayson’s ligaments are more delicate, may even be discontinuous and pass from the lateral sides of the phalanges volar to the neurovascular bundles to insert into the lateral digital sheet. Landsmeer’s ligaments are inconsistent anatomical structures made up of transverse and oblique retinacular ligaments (see Fig. 50.30). The transverse retinacular ligament passes from the A3 pulley of the fibrous flexor sheath at the level of the proximal interphalangeal joint to the lateral border of the lateral extensor band. The oblique retinacular ligament lies deep to the transverse retinacular ligament. It originates from the lateral aspect of the proximal phalanx and flexor sheath (A2 pulley) and passes volar to the axis of rotation of the proximal interphalangeal joint in a dorsal and distal direction to insert into the terminal extensor tendon.
Functions of the fascia of the hand
Channelling of structures in transit between forearm and digits
The vertical septa act as spacers between the tendons and neurovascular bundles of the individual digital rays. Where tendons change direction around a concave surface the channels are thickened. They perform a retinacular role, forming sheaths with specialized pulleys to prevent the tendon springing away from the underlying skeleton (see flexor tendon sheaths, p. 879).
Digital and palmar spaces
There are many potential spaces within the hand, often with ill-defined margins.
The nail fold is a ‘U-shaped’ space made up of the eponychium and the lateral nail fold. The apical spaces at the tip of the finger are formed by the fibrous attachments of the distal phalanx to the tip of the digital pulp skin. The digital pulp spaces are confined compartments bounded by the digital creases which overlie the joints, and are attached to the underlying pulleys. The synovial flexor tendon sheaths are described on page 879. The web space is bounded distally by the skin and natatory ligament, by the deep transverse metacarpal ligament posteriorly, and by the deep attachments of the palmar fascia, together with their lateral attachments to the tendon sheaths proximally. The deep palmar space is a complex three-dimensional space limited proximally by the carpal tunnel. It lies deep to the palmar aponeurosis, between the radial and ulnar condensations of vertical fibres which connect the palmar aponeurosis to the thenar and hypothenar eminences. Partitions that pass deeply from the longitudinal bands of the palmar aponeurosis form eight narrow compartments: four contain the digital flexor tendons and four contain the lumbricals and the neurovascular bundles.
BONE
CARPAL BONES
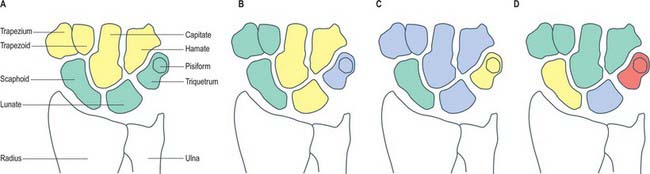
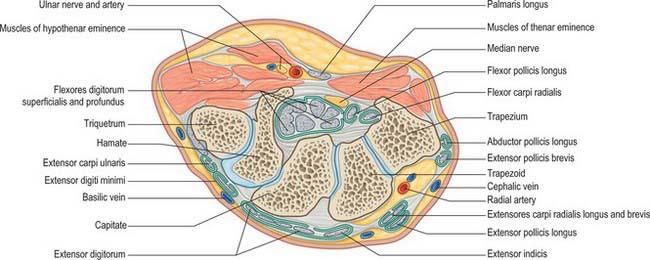
The carpus contains eight bones: four each in proximal and distal rows (Fig. 50.5 and Fig. 50.6). In radial (lateral) to ulnar (medial) order, the scaphoid, lunate, triquetrum and pisiform make up the proximal row, and the trapezium, trapezoid, capitate and hamate make up the distal row. The pisiform articulates with the palmar surface of the triquetrum, and is thus separated from the other carpal bones, all of which articulate with their neighbours. The other three proximal bones form an arch which is proximally convex, and which articulates with the radius and articular disc of the distal radio-ulnar joint. The concavity of the arch is a distal recess embracing, proximally, the projecting aspects of the capitate and hamate. The two rows of carpal bones are thus mutually and firmly adapted without any loss of movement.
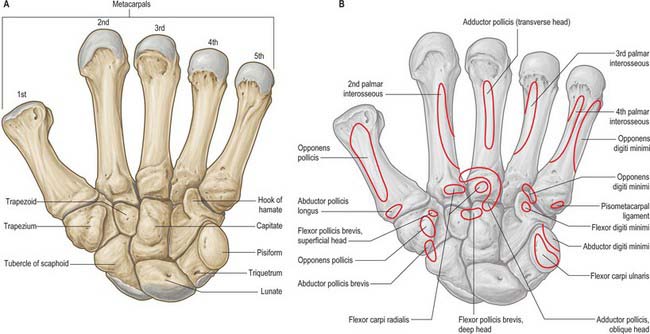
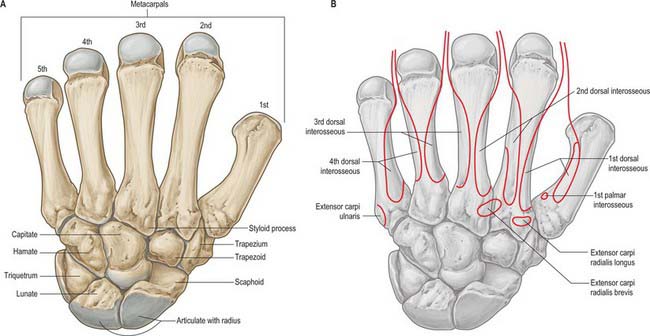
The dorsal carpal surface is convex. The palmar surface forms a deeply concave carpal groove, accentuated by the palmar projection of the radial (lateral) and ulnar (medial) borders. The ulnar projection is formed by the pisiform and the hamulus (hook), an unciform palmar process of the hamate. The pisiform is at the proximal border of the hypothenar eminence, on the ulnar side of the palm, and it is easily felt in front of the triquetrum. The hamulus is concave in a radial direction, its tip is palpable 2.5 cm distal to the pisiform, in line with the radial border of the ring finger. The superficial division of the ulnar nerve can be rolled on it. The radial border of the carpal groove is formed by the tubercles of the scaphoid and trapezium. The former is distal on the anterior scaphoid surface and palpable (sometimes also visible) as a small medial knob at the proximal border of the palmar thenar eminence, radial to the tendon of flexor carpi radialis. The tubercle of the trapezium is a vertically rounded ridge on the anterior surface of the bone, slightly hollow medially and just distal and radial to the scaphoid tubercle: it is difficult to palpate. (Both the scaphoid and trapezium may be grasped individually, and moved passively, by firm pressure between an opposed index finger and thumb applied to the palmar surface and ‘anatomical snuff-box’ simultaneously.) The carpal groove is made into an osseofibrous carpal tunnel by a fibrous retinaculum attached to its margins. The tunnel carries flexor tendons and the median nerve into the hand. The retinaculum strengthens the carpus and augments flexor efficiency. Radiocarpal, intercarpal and carpometacarpal ligaments are attached to the palmar and dorsal surfaces of all of the carpal bones, except the triquetrum and pisiform.
Individual carpal bones
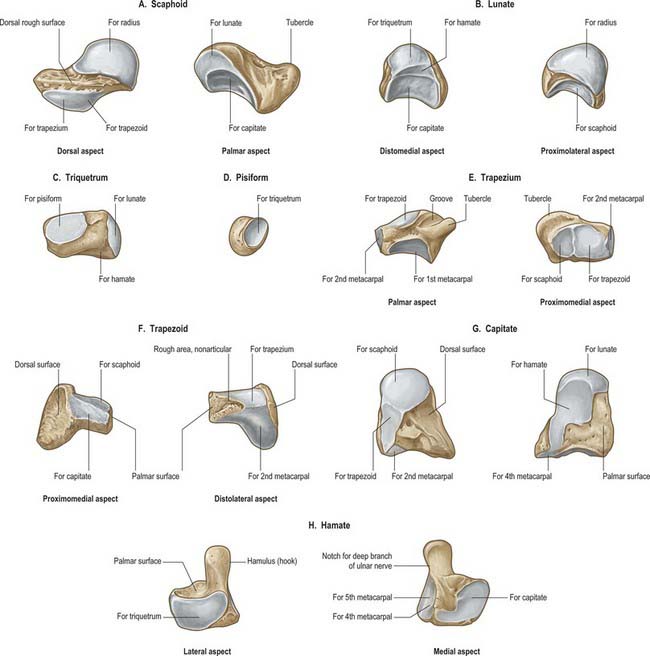
Scaphoid
The scaphoid is the largest element in the proximal carpal row (Fig. 50.7A). It has a long axis which is distal, radial and slightly palmar in direction. A round tubercle on the distolateral part of its palmar surface is directed anterolaterally (Fig. 50.5), and provides an attachment for the flexor retinaculum and abductor pollicis brevis: it is crossed by the tendon of flexor carpi radialis. The rough dorsal surface is slightly grooved, narrower than the palmar, and pierced by small nutrient foramina, which are often restricted to the distal half (13%). The radial collateral ligament is attached to the lateral surface, which is also narrow and rough. The remaining surfaces are all articular. The radial (proximal) surface is convex, proximal and directed proximolaterally; the lunate surface is flat, semilunar, and faces medially; the capitate surface is large, concave and distal, and directed distomedially. The surface for the trapezium and trapezoid is continuous, convex and distal.
Lunate
The lunate is approximately semilunar and articulates between the scaphoid and triquetrum in the proximal carpal row (Fig. 50.7B). Its rough palmar surface, almost triangular, is larger and wider than the rough dorsal surface. Its smooth convex proximal surface articulates with the radius and the articular disc of the distal radio-ulnar joint. Its narrow lateral surface bears a flat semilunar facet for the scaphoid. The medial surface, almost square, articulates with the triquetrum and is separated from the distal surface by a curved ridge, usually somewhat concave for articulation with the edge of the hamate in adduction (Fig. 50.7B, left). The distal surface is deeply concave to fit the medial part of the head of the capitate.
Triquetrum
The triquetrum is somewhat pyramidal and bears an oval isolated facet for articulation with the pisiform on its distal palmar surface (Fig. 50.7C). Its medial and dorsal surfaces are confluent, and marked distally by the attachment of the ulnar collateral ligament, but smooth proximally to receive the articular disc of the distal radio-ulnar joint in full adduction. The hamate surface, lateral and distal, is concavoconvex, broad proximally, narrow distally. The lunate surface, almost square, is proximal and lateral.
Pisiform
The pisiform is shaped like a pea, with a distolateral long axis (Fig. 50.7D). It bears a dorsal flat articular facet for the triquetrum. The tendon of flexor carpi ulnaris and the distal continuations of the tendon, the pisometacarpal and pisohamate ligaments, are all attached to the palmar non-articular area, which surrounds and projects distal to the articular surface: the pisiform therefore has attributes of a sesamoid bone.
Trapezium
The trapezium has a tubercle and groove on its rough palmar surface (Fig. 50.7E). The groove, which is medial, contains the tendon of flexor carpi radialis, and two layers of the flexor retinaculum are attached to its margins. The tubercle is obscured by the thenar muscles which are attached to it (opponens pollicis, flexor pollicis brevis and abductor pollicis brevis) (Fig. 50.5B). The elongated, rough dorsal surface is related to the radial artery. The large lateral surface is rough for attachment of the radial collateral ligament and capsular ligament of the thumb carpometacarpal joint. A large sellar surface faces distolaterally and articulates with the base of the first metacarpal. Most distally it projects between the bases of the first and second metacarpal bones and carries a small, quadrilateral, distomedially directed facet which articulates with the base of the second metacarpal. The large medial surface is gently concave for articulation with the trapezoid. The proximal surface is a small, slightly concave facet for articulation with the scaphoid. Its ridge, or ‘summit’, fits the concavity of the first metacarpal base, and extends in a palmar and lateral direction, at an angle of approximately 60° with the plane of the second and third metacarpals. Abduction and adduction occur in the plane of the ridge, which is shorter than the corresponding metacarpal groove. Their contours vary reciprocally: they are more curved near the second metacarpal base, whereas the radius of curvature is longer further away from this site. The two surfaces are not completely congruent, and the area of close contact probably moves towards the palm in adduction and dorsally in abduction. While the axis of flexion/extension passes through the trapezium, that for adduction/abduction is in the metacarpal base. Flexion is accompanied by medial rotation, and extension by lateral rotation (p. 875).
Trapezoid
The trapezoid is small and irregular. It has a rough palmar surface which is narrower and smaller than its rough dorsal surface: the former invades the lateral aspect (Fig. 50.7F). The distal surface, which articulates with the grooved base of the second metacarpal, is triangular, convex transversely and concave at right angles to this. The medial surface articulates by a concave facet with the distal part of the capitate, the lateral surface articulates with the trapezium, and the proximal surface articulates with the scaphoid.
Capitate
The capitate is the central and largest carpal bone. It articulates with the base of the third metacarpal via its triangular distal concavoconvex surface (Fig. 50.7G). Its lateral border is a concave strip for articulation with the medial side of the base of the second metacarpal. Its dorsomedial angle usually bears a facet for articulation with the base of the fourth metacarpal. The head projects into the concavity formed by the lunate and scaphoid: the proximal surface articulates with the lunate, and the lateral surface with the scaphoid. The facets for the scaphoid and trapezoid, though usually continuous on the distolateral surface, may be separated by a rough interval. The medial surface bears a large facet for articulation with the hamate, which is deeper proximally where it is partly non-articular. Palmar and dorsal surfaces are roughened for carpal ligaments, the dorsal being the larger.
Hamate
The hamate is cuneiform and bears an unciform hamulus (hook) which projects from the distal part of its rough palmar surface. The hamulus is curved with a lateral concavity and its tip inclines laterally, contributing to the medial wall of the carpal tunnel (Fig. 50.7H). The flexor retinaculum is attached to the apex of the hamulus. Distally, on the hamular base, a slight transverse groove may be in contact with the terminal deep branch of the ulnar nerve. The remaining palmar surface, like the dorsal, is roughened for attachment of ligaments. A faint ridge divides the distal surface into a smaller lateral facet which articulates with the base of the fourth metacarpal base, and a medial facet for articulation with the base of the fifth. The proximal surface, the thin margin of the wedge, usually bears a narrow facet which contacts the lunate in adduction. The medial surface is a broad strip, convex proximally, concave distally, which articulates with the triquetrum: distally a narrow medial strip is non-articular. The lateral surface articulates with the capitate by a facet covering all but its distal palmar angle.
Ossification
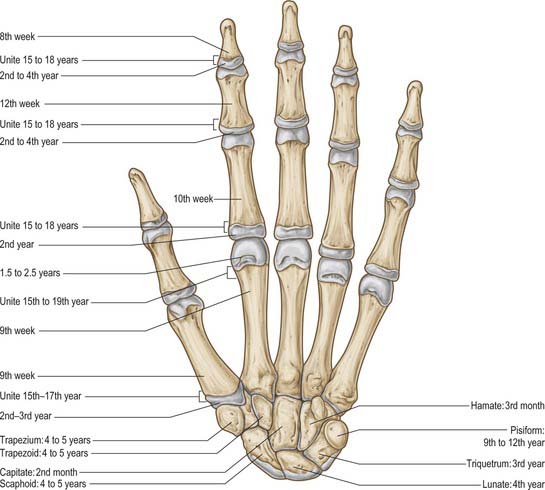
Carpal bones are cartilaginous at birth, although ossification may have started in the capitate and hamate. Each carpal bone is ossified from one centre, capitate first, and pisiform last: the order in the others varies (Fig. 50.8, Fig. 5.9, Fig. 5.10, Fig. 5.11, Fig. 50.12). The capitate begins to ossify in the second month, the hamate at the end of the third month, the triquetrum in the third year, the lunate, scaphoid, trapezium and trapezoid in the fourth year in females and fifth year in males. The pisiform begins to ossify in the ninth or tenth year in females, and the twelfth in males. The order varies according to sex, nutrition and, possibly, race. Occasionally an os centrale occurs between the scaphoid, trapezoid and capitate bones: during the second prenatal month it is a cartilaginous nodule which usually fuses with the scaphoid. Occasionally, lunate and triquetral elements may fuse. Other fusions and accessory ossicles have also been described.
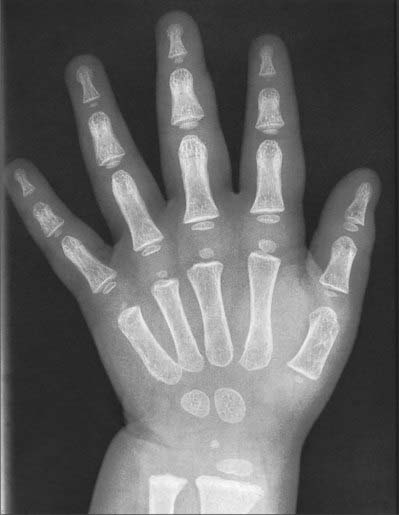
Fig. 50.8 Radiograph of a hand at 2½ years (male), dorsopalmar projection. Note early stages of ossification in the epiphyses at the proximal ends of the phalanges and first metacarpal; at the distal ends of the remaining metacarpals and radius; in the capitate, hamate and lunate. Typically, the centre for the lunate is preceded by the centre for the triquetrum. Compare with Figs 50.9 and 50.10.
METACARPALS
Individual metacarpal bones
First metacarpal
The first metacarpal is short and thick (Fig. 50.13A). Its dorsal (lateral) surface can be felt to face laterally; its long axis diverges distolaterally from its neighbour. The shaft is flattened, dorsally broad and transversely convex. The palmar (medial) surface is longitudinally concave and divided by a ridge into a larger lateral (anterior) and smaller medial (posterior) part. Opponens pollicis is attached to the radial border and adjoining palmar surface; the first dorsal interosseous muscle (radial head) is attached to its ulnar border and adjacent palmar surface. The base is concavoconvex and articulates with the trapezium. Abductor pollicis longus is attached on its lateral (palmar) side, the first palmar interosseous muscle to its ulnar side. The head is less convex than in other metacarpals and is transversely broad. Sesamoid bones glide on radial and ulnar articular eminences on its palmar aspect.
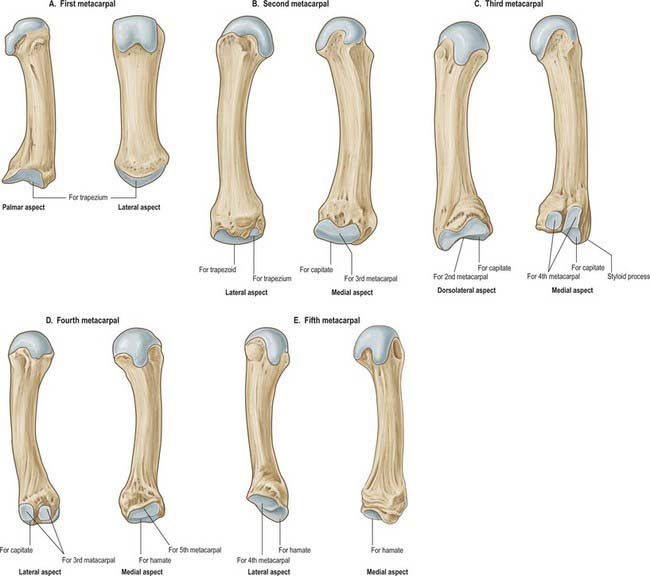
Second metacarpal
The second metacarpal has the longest shaft and largest base (Fig. 50.13B). The latter is grooved in a dorsopalmar direction for articulation with the trapezoid. Medial to the groove a deep ridge articulates with the capitate; laterally, nearer the dorsal surface of the base, there is a quadrilateral facet for articulation with the trapezium, and just dorsal to this facet a rough impression marks the attachment of extensor carpi radialis longus. On the palmar surface a small tubercle or ridge receives flexor carpi radialis. The medial side of the base articulates with that of the third metacarpal by a long facet, centrally narrowed. The shaft is prismatic in section and longitudinally curved, convex dorsally, concave towards the palm. Its dorsal surface is distally broad but proximally narrows to a ridge which is covered by extensor tendons of the index finger. Its converging borders begin at the tubercles, one on each side of its head for the attachment of collateral ligaments. Proximally the lateral surface inclines dorsally for the ulnar head of the first dorsal interosseous. The medial surface inclines similarly, and is divided by a faint ridge into a palmar strip for attachment of the second palmar interosseous, and a dorsal strip for attachment of the radial head of the second dorsal interosseous.
Third metacarpal
The third metacarpal has a short styloid process, projecting proximally from the radial side of the dorsal surface (Fig. 50.13C). Its base articulates with the capitate by a facet anteriorly convex but dorsally concave where it invades the styloid process on the lateral aspect of its base. A strip-like facet, constricted centrally, articulates with the bases of the second metacarpal (laterally) and the fourth metacarpal (medially), the latter by two oval facets. The palmar facet may be absent; less frequently the facets are connected proximally by a narrow bridge. The palmar surface of the base receives a slip from the tendon of flexor carpi radialis; extensor carpi radialis brevis is attached to its dorsal surface, beyond the styloid process. The shaft resembles that of the second metacarpal. The ulnar head of the second dorsal interosseous is attached to its lateral surface; the radial head of the third dorsal interosseous is attached to its medial surface, and the transverse head of adductor pollicis is attached to the intervening palmar ridge in its distal two-thirds. Its dorsal surface is covered by the extensor tendon.
Fourth metacarpal
The fourth metacarpal is shorter and thinner than the second and third (Fig. 50.13D). On its base it displays two lateral oval facets for articulation with the base of the third metacarpal; the dorsal is usually larger and proximally in contact with the capitate. A single medial elongated facet is for articulation with the base of the fifth metacarpal. The quadrangular proximal surface articulates with the hamate, and is anteriorly convex, dorsally concave. The shaft is like the second, but a faint ridge on its lateral surface separates the attachments of the third palmar interosseous and the ulnar head of the third dorsal interosseous. The radial head of the fourth dorsal interosseous is attached to the medial surface.
Fifth metacarpal
The fifth metacarpal (Fig. 50.13E) differs in its medial basal surface, which is non-articular and bears a tubercle for extensor carpi ulnaris. The lateral basal surface is a facet, transversely concave, convex from palm to dorsum, for articulation with the hamate. A lateral strip articulates with the base of the fourth metacarpal. The shaft bears a triangular dorsal area which almost reaches the base; the lateral surface inclines dorsally only at its proximal end. Opponens digiti minimi is attached to the medial surface. The lateral surface is divided by a ridge, which is sometimes sharp, into a palmar strip for the attachment of the fourth palmar interosseous and a dorsal strip for the ulnar part of the fourth dorsal interosseous.
Ossification
Each metacarpal ossifies from a primary centre for the shaft and a secondary centre which is in the base of the first metacarpal and in the heads of the other four (Figs 50.8–50.11). Ossification begins in the midshaft about the ninth week. Centres for the second to fifth metacarpal heads appear in that order in the second year in females, and between 1½ to 2½ years in males. They unite with the shafts about the 15th or 16th year in females, 18th or 19th in males. The first metacarpal base begins to ossify late in the second year in females, early in the third year in males, uniting before the 15th year in females and 17th in males. Sometimes the styloid process of the third metacarpal is a separate ossicle. The thumb metacarpal ossifies like a phalanx, and some authorities therefore consider that the thumb skeleton consists of three phalanges. Others believe that the distal phalanx represents fused middle and distal phalanges, a condition occasionally observed in the fifth toe. When the thumb has three phalanges, the metacarpal has a distal and a proximal epiphysis. It occasionally bifurcates distally. When it does, the medial branch has no distal epiphysis and bears two phalanges, while the lateral branch shows a distal epiphysis, and three phalanges. The existence of only a distal metacarpal epiphysis may be associated with a greater range of movement at the metacarpophalangeal joint. In the thumb, the carpometacarpal joint has the wider range, and the first metacarpal has a basal epiphysis. A distal epiphysis may appear in the first, and a proximal epiphysis in the second, metacarpal.
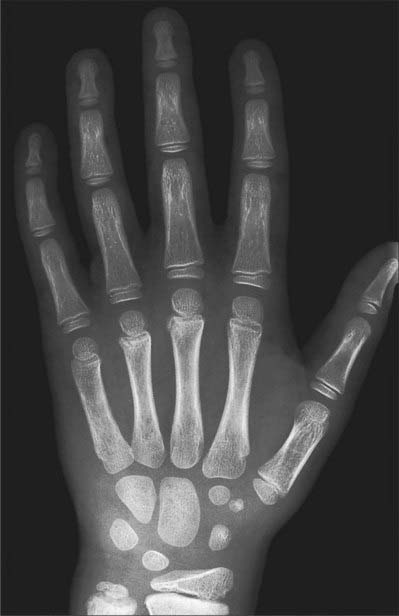
Fig. 50.9 Radiograph of a hand at 6½ years (male), dorsopalmar projection. Note the more advanced state of the centres of ossification which were already visible in Fig. 50.8, and the appearance of additional centres in the distal ulnar epiphysis and in the triquetrum, scaphoid, trapezium and trapezoid.
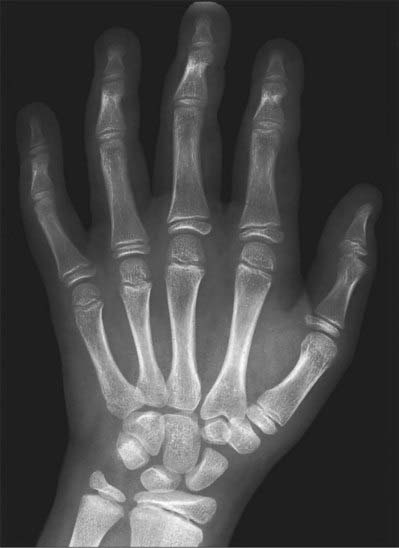
Fig. 50.10 Radiograph of a hand at 11 years (female), dorsopalmar projection. Note the maturing shapes of all the ossifications previously seen in Figs 50.8 and 50.9, with the addition of the pisiform.
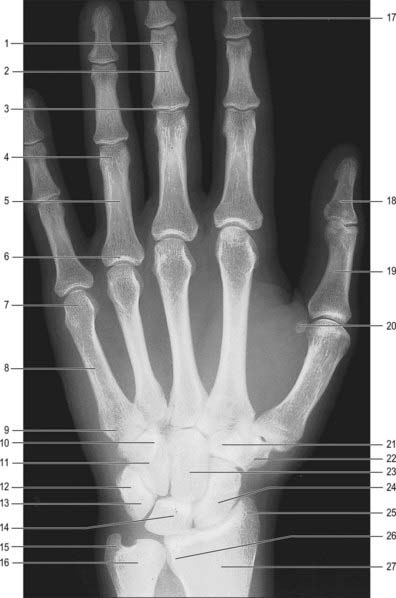
PHALANGES
The phalanges of the little finger and the thumb differ. Abductor and flexor digiti minimi are attached to the medial side of the base of the proximal phalanx of the little finger. The tendon of extensor pollicis brevis and the oblique head of adductor pollicis (dorsally), and the oblique and transverse heads of adductor pollicis, sometimes conjoined with the first palmar interosseous (medially), are attached to the base of the proximal pollicial phalanx.
Ossification
Phalanges are ossified from a primary centre for the shaft and a proximal epiphysial centre (Figs 50.8–50.12). Ossification begins prenatally in shafts as follows: distal phalanges in the eighth or ninth week, proximal phalanges in the tenth, middle phalanges in the 11th week or later. Epiphysial centres appear in proximal phalanges early in the second year (females), and later in the same year (males), and in middle and distal phalanges in the second year (females), or third or fourth year (males). All epiphyses unite about the 15th to 16th year in females, and 17th to 18th year in males.
JOINTS
DISTAL RADIO-ULNAR JOINT
The distal radio-ulnar joint is a uniaxial pivot joint (Fig. 50.14).
RADIOCARPAL (WRIST) JOINT
The radiocarpal joint is a synovial biaxial and ellipsoid joint formed by articulation of the distal end of the radius and the triangular fibrocartilage with the scaphoid, lunate and triquetrum (Fig. 50.15 and Fig. 50.16). In the neutral position of the wrist, only the scaphoid and lunate are in contact with the radius and articular disc: the triquetrum comes into apposition with the disc only in full adduction of the wrist joint. The radial articular surface and distal discal surface form an almost elliptical, concave surface with a transverse long axis. The radial surface is bisected by a low ridge into two concavities. A similar ridge usually appears between the medial radial concavity and the concave distal discal surface. The proximal articular surfaces of the scaphoid, lunate and triquetrum, and their interosseous ligaments, form a smooth convex surface which is received into the proximal concavity.
CARPAL JOINTS
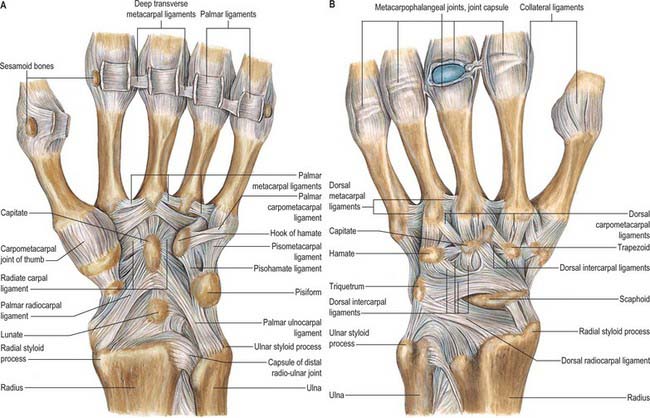
Wrist ligaments
Extrinsic ligaments
Extrinsic palmar carpal ligaments
When the synovial lining of the carpal tunnel is dissected away, two V-shaped ligamentous bands are visible with their apices lying distally (Fig. 50.15A). The limbs of the ‘V’ take origin from the radius and ulna respectively: the apex of one ‘V’ attaches to the distal row and that of the second ‘V’ to the proximal row.
The long radiolunate ligament takes origin adjacent to the radioscaphocapitate ligament on the palmar lip of the radius. It passes over and supports the proximal pole of the scaphoid before inserting into the palmar horn of the lunate. This ligament is discrete from the radioscaphocapitate ligament and the visible separation is known as the interligamentous sulcus (continuous with the space of Poirier).
Extrinsic dorsal carpal ligaments
The dorsal wrist ligaments are comparatively thin. They are reinforced by the floor and septa of the fibrous tunnels for the six dorsal compartments. The extrinsic dorsal carpal ligaments and the intrinsic dorsal intercarpal ligaments have a ‘Z-shaped’ configuration (Fig. 50.15B). The pattern and shape of these ligaments is utilized in one surgical approach to the dorsum of the wrist joint where incisions are oriented parallel with the ligaments: this reduces scarring and restriction of subsequent motion caused by the arthrotomy.
Triangular fibrocartilage complex (TFCC) and distal radio-ulnar ligaments
The triangular fibrocartilage complex (TFCC) is a ligamentous and cartilaginous structure which suspends the distal radius and ulnar carpus from the distal ulna. The TFCC stabilizes the ulnocarpal and radio-ulnar joints, transmits and distributes load from the carpus to the ulna, and facilitates complex movements at the wrist (Fig. 50.17). By definition, it is made up of the cartilaginous disc, the meniscus homologue (an embryological remnant of the ‘ulnar’ wrist that is only occasionally present), volar and dorsal distal radio-ulnar ligaments, ulnar collateral ligament, floor of extensor carpi ulnaris subsheath, ulnolunate and ulnotriquetral ligaments. The triangular fibrocartilage proper (TFC) is a biconcave body composed of chondroid fibrocartilage. It extends across the dome of the ulnar head and varies between 2 and 5 mm in thickness.
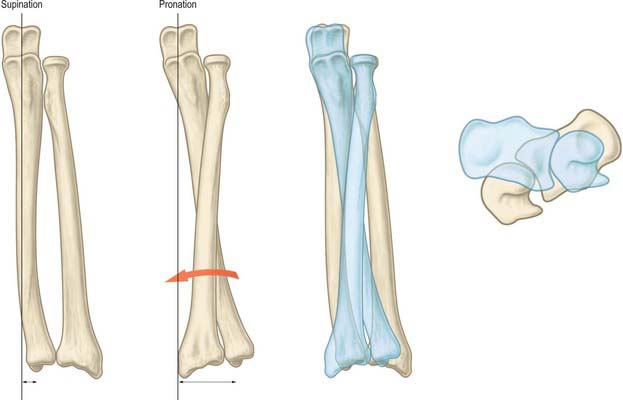
COORDINATED MOVEMENTS AND LOAD-BEARING AT THE WRIST JOINT
Wrist movements
The range of flexion is greater at the radiocarpal joint, while in extension there is more movement at the midcarpal joint (Fig. 50.18). Hence the proximal surfaces extend further posteriorly on the lunate and scaphoid bones. These movements are limited chiefly by antagonistic muscles, and therefore the range of flexion is perceptibly diminished when the fingers are flexed, due to increased tension in the extensors. Only when the joints are forced to the limits of flexion or extension are the dorsal or palmar ligaments fully stretched (but see below).
Adduction of the hand is considerably greater than abduction, perhaps due to the more proximal site of the ulnar styloid process. Most adduction occurs at the radiocarpal joint. The lunate articulates with both the radius and articular disc when the hand is in the midposition, but in adduction it articulates solely with the radius, and the triquetrum now comes into contact with the articular disc (Fig. 50.19A). Much of the proximal articular surface of the scaphoid becomes subcapsular beneath the radial collateral ligament and forms a smooth, convex, palpable prominence in the floor of the ‘anatomical snuff-box’.
Abduction from the neutral position occurs at the midcarpal joint, the proximal carpal row not moving. Radiographs of abducted hands show that the capitate rotates round an anteroposterior axis so that its head passes medially and the hamate conforms to this: the distance between the lunate and the apex of the hamate is increased (Fig. 50.19B). The scaphoid rotates around a transverse axis, and its proximal articular surface moves away from the capsule to articulate solely with the radius. Movements are limited by antagonistic muscles and, at extremes, by the carpal collateral ligaments.
Integrated model of wrist movement (carpal kinematics)
The proximal row (scaphoid, lunate and triquetrum) is an intercalated segment: no tendons insert onto the bones of the row. It is inherently unstable and controlled by specific retaining and gliding ligaments. Its relative position is determined by the spatial configurations of the radius, triangular fibrocartilage complex (TFCC) and ulna on one side, and the rigid distal carpal row on the other. The proximal carpal row is subject to two opposing moments: the scaphoid straddles the proximal and distal rows and tends to rotate the proximal row into flexion under axial load and radial deviation. At the same time there is a force tending to extend the proximal row which is initiated by the distal row and transmitted via the midcarpal ligaments to the triquetrum (Fig. 50.20). Stability of the midcarpal joint is thus ensured during both movement and loading.
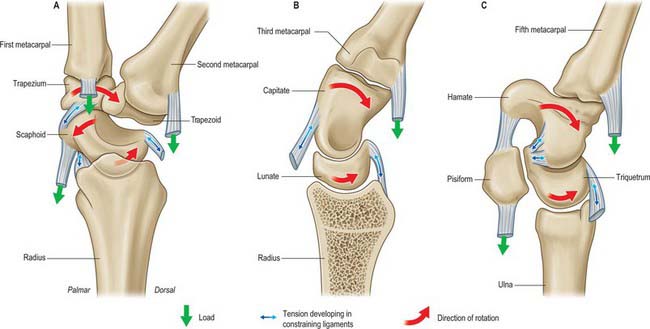
The carpus was originally thought to move simply as proximal and distal rows (row or rigid body theory). According to this view, during the composite movement of wrist flexion and extension, approximately two-thirds of movement occurs at the radiocarpal joint and one-third at the midcarpal joint. The carpus was later judged to move in lateral central and medial columns more than it did in rows, and the radius–lunate–capitate was described as a three-bar linkage system (column theory). This theory was modified to incorporate the specific stabilizing role of the scaphoid as it bridges the proximal and distal rows. A further theory proposed that the bones were linked by their ligaments in a ring configuration, so that any breakage of the key links leads to instability (ring theory). Most recently the ‘four unit’ theory suggests that the distal carpal row moves as a single unit, and the scaphoid, lunate and triquetrum move in complex but characteristic relationships which are dependent on the given movement (Fig. 50.21). Clinical observation provides some support for each of these theories.
Wrist loading
Force transmission
Radiocarpal joint
The articulation of the carpus with the radius (radiocarpal) and ulna (ulnocarpal) can be described in terms of specific fossae, i.e. the scaphoid and lunate fossae of the radius and the facet formed by the distal aspect of the TFCC. The scaphoid fossa contributes 43%, the lunate fossa 46%, and the TFCC 11%, of the total area of this articulating surface. Under physiological conditions, the contact area between this surface and the proximal carpal row is of the order of 20% of the whole. It varies consistently with position: greater contact is seen in forearm supination, radial deviation and dorsiflexion of the wrist, and lesser contact in forearm pronation, ulnar deviation and palmar flexion of the wrist. Further increase in contact area is seen with axial loading. Maximum contact areas are recorded at 40%. Force across the joint also varies with the position of the forearm and the degree of wrist flexion. For a given load in wrist neutral position, 50% of force passes across the scaphoid fossa, 35% through the lunate fossa, and 15% across the TFCC. With a power grip there is slight ulnar deviation, and the proportion of force passing across the scaphoid and lunate fossae reverses.
Specific actions
Hammer action
The action exemplified by hammering, where the wrist alternates between the dorsoradial and the palmar-ulnar position, is an important movement. The reciprocal movement is more constrained and awkward to perform. The former movement is important in a wide variety of activities of daily living. In primates this movement is needed (along with forearm rotation at the distal radio-ulnar joint) for brachiation.
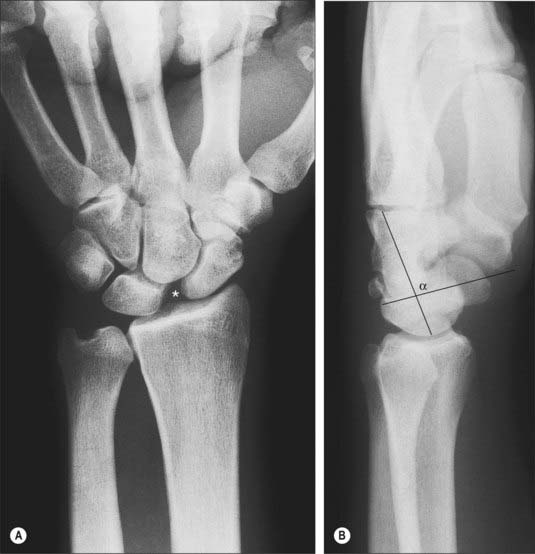
Wrist instability
Some degree of carpal instability may occur as part of inherent hypermobility in affected individuals, but usually it occurs only after rupture or attenuation of intrinsic and extrinsic carpal ligaments and may be detected either clinically or evidenced on radiographs as malalignment of the carpal bones (Fig. 50.22).
CARPOMETACARPAL JOINTS
Carpometacarpal joint of the thumb
Except at initiation, flexion is accompanied by medial rotation; conversely, medial rotation involves flexion. Linkage of movements is due largely to the shape of the articular surfaces (which impose some conjunct rotation), and to the obliquity of the dorsal ligament (which, when taut, anchors the ulnar side of the metacarpal base while its radial side continues to move). Contraction of flexor pollicis brevis, assisted by opponens pollicis, thus produces medial rotation with flexion; combined with abduction this brings the thumb pulp into contact with the pulps of the slightly flexed fingers, a movement termed opposition. (The flexed fingers have varying degrees of lateral metacarpophalangeal rotation, which is minimal in the index, and maximal in the minimus.) Conversely, full extension of the thumb metacarpal entails slight lateral rotation, attributable to the sellar form of the joint and to the action of the palmar ligament (which is similar to that of the dorsal ligament in flexion). (See also p. 888.)
INTERMETACARPAL JOINTS
METACARPOPHALANGEAL JOINTS
The metacarpophalangeal joints are usually considered ellipsoid. However, the metacarpal heads are adapted to shallow concavities on the phalangeal bases: they are not regularly convex but partially divided on their palmar aspects and thus almost bicondylar (Fig. 50.23).
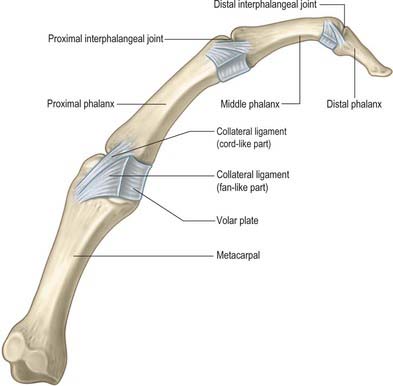
Each metacarpophalangeal joint has a palmar and two collateral ligaments.
Deep transverse metacarpal ligaments
The deep transverse metacarpal ligaments are three short, wide, flat bands which connect the palmar ligaments of the second to fifth metacarpophalangeal joints. They are related anteriorly to the lumbricals and digital vessels and nerves, and posteriorly to the interossei. Bands from the digital slips of the central palmar aponeurosis join their palmar surfaces. On both sides of the third and fourth metacarpophalangeal joints, but only the ulnar side of the second and radial side of the fifth, transverse bands of the dorsal digital expansions join the deep transverse metacarpal ligaments. The lumbricals and the phalangeal attachments of the dorsal interossei lie anterior to this band, and the remaining attachments of dorsal interossei and palmar interossei are posterior to it (see Fig. 50.30).
INTERPHALANGEAL JOINTS
The interphalangeal joints are uniaxial hinge joints (Fig. 50.23).
The volar plate constitutes the floor of the interphalangeal joint. In the proximal interphalangeal joint, the distal end of the volar plate is thickened laterally where it is firmly attached to the base of the middle phalanx at the position of the true collateral ligament attachment. Centrally, it is more delicate and blends with the volar periosteum of the middle phalanx. Proximally, the volar plate is also very delicate in its central portion, but it is thickened laterally to form the so-called check rein ligaments which attach to the periosteum of the proximal phalanx just within the walls of the A2 pulley component of the fibrous flexor sheath. Nutrient branches of the digital arteries pass underneath these check ligaments to reach the vinculae (Fig. 50.24). A detailed account of the anatomy of the volar plate can be found in Bowers et al (1980).
MUSCLE
EXTRINSIC LONG FLEXORS AND EXTENSORS
The extrinsic long flexors and extensors are described on page 845.
Flexor retinaculum
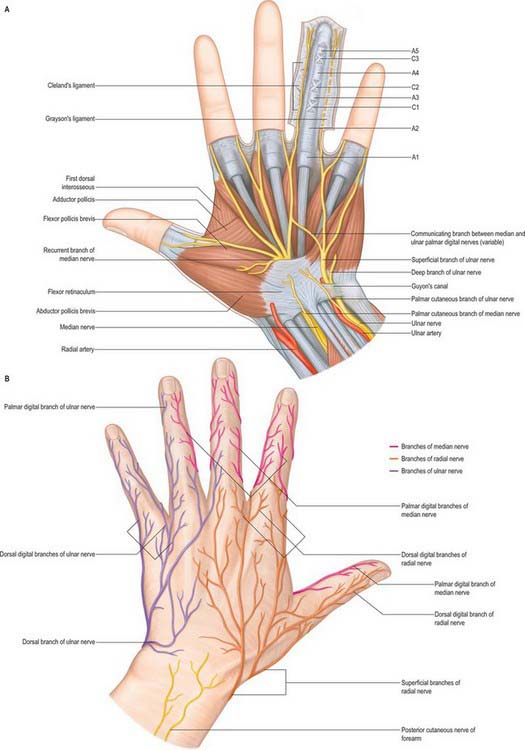
The flexor retinaculum is a strong, fibrous band (Fig. 50.25). It crosses the front of the carpus and converts its anterior concavity into the carpal tunnel which transmits the flexor tendons of the digits and the median nerve. The retinaculum is short and broad, measuring 2.5–3 cm both transversely and proximodistally. It is attached medially to the pisiform and the hook of the hamate. Laterally, it splits into superficial and deep laminae. The superficial lamina is attached to the tubercles of the scaphoid and trapezium. The deep lamina is attached to the medial lip of the groove on the trapezium. Together with this groove, the two laminae form a tunnel, lined by a synovial sheath, which contains the tendon of flexor carpi radialis. The retinaculum is crossed superficially by the ulnar vessels and nerve – immediately radial to the pisiform – and by the palmar cutaneous branches of the median and ulnar nerves. A slender band of fascia, the superficial part of the flexor retinaculum, bridges over the ulnar neurovascular bundle and attaches to the radial side of the pisiform, forming a tunnel (Guyon’s canal) which is an occasional site of ulnar nerve entrapment. The tendons of palmaris longus and flexor carpi ulnaris are partly attached to the anterior surface of the retinaculum. Distally some of the intrinsic muscles of the thumb and little finger are attached to the retinaculum.
Long flexor tendon apparatus
Flexor tendon sheaths
The fibrous sheaths of the flexor tendons are specialized parts of the palmar fascial continuum. Each finger has an osseoaponeurotic tunnel which extends from midpalm to the distal phalanx. The thumb has a tunnel for flexor pollicis longus which extends from the metacarpal to the distal phalanx. The proximal border is to some extent a matter of definition, because the transverse fibres of the palmar aponeurosis may be considered to be a part of the pulley system. The sheath consists of arcuate fibres which arch anteriorly over bone, tendons (where the sheath is required to be stiff), and the centres of joints (where a bucket-handle of arcuate fibres is a mechanically favourable arrangement). In contrast, where the sheath is required to fold to permit joint flexion, it consists of cruciate fibres. These fibrous sheaths are lined by a thin synovial membrane which provides a sealed lubrication system containing synovial fluid. The synovial membrane extends from the distal phalanx to midpalm in the case of the index, middle and ring fingers, and further proximally in the case of the little finger (Fig. 50.26). The sheaths around the thumb and little finger are continuous with the flexor sheaths in front of the wrist. The parietal synovial membrane is reflected onto the surface of the flexor tendon, forming a visceral synovium.
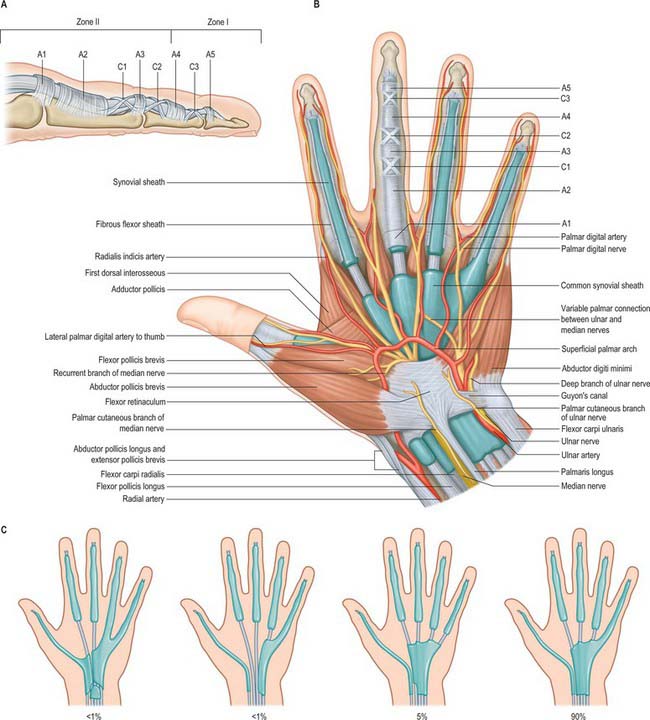
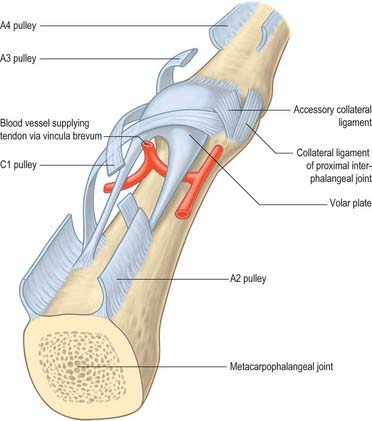
A standard nomenclature for the anular and cruciform pulleys that make up the sheath has been adopted by the American Society for Surgery of the Hand: the letters A and C respectively are used (Doyle & Blythe 1975).
The usual pattern is as follows (Fig. 50.26). The A1 pulley is situated anterior to the palmar cartilaginous plate of the metacarpophalangeal joint and may extend over the proximal part of the proximal phalanx. The A2 overlies the middle third of the proximal phalanx. It is the strongest pulley and arises from well-defined longitudinal ridges on the palmar aspect of the phalanx. Its distal edge is well developed. A pouch or recess of synovium extends superficial to the free edge of the pulley fibres so that the free edge forms a lip protruding into the synovial space. A3 is a narrow pulley lying palmar to the proximal interphalangeal joint. A4 overlies the middle third of the middle phalanx, and A5 overlies the distal interphalangeal joint. The cruciate fibres are numbered in a slightly different manner. C0 is palmar to the metacarpophalangeal joint. There are two cruciate zones, C1 and C2, at the proximal interphalangeal joint and they lie just proximal and distal respectively to A3. At the distal interphalangeal joint there is one pronounced cruciate system, C3, which lies between A4 and A5. Variations occur frequently. During flexion, the cruciate fibres become orientated more transversely in the digits and the edges of adjacent anular pulleys approximate so that they form, in full flexion, a continuous tunnel of transversely orientated fibres. Surgically the most important pulleys which prevent bowstringing of the flexor tendons are the A2 and A4 pulleys.
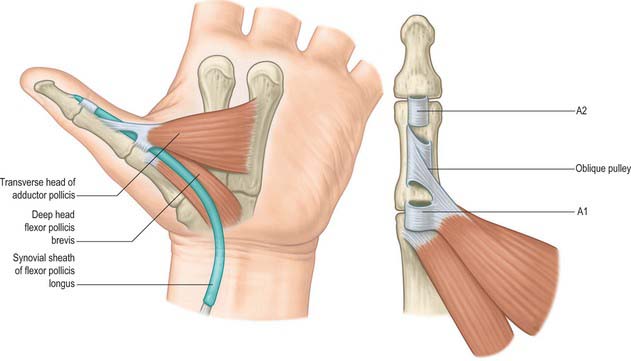
The pulley arrangement in the thumb is different from that in the other digits. There are three constant pulleys – two anular and one oblique (Fig. 50.27). The A1 pulley is located at the metacarpophalangeal joint. The oblique pulley is located over the mid-portion of the proximal phalanx, and its fibres pass from the ulnar aspect proximally to the radial aspect dorsally. The A2 pulley is thinner than the A1 pulley and is situated just proximal to the interphalangeal joint. The oblique pulley is the most important pulley in the thumb for maintaining the action of flexor pollicis longus.
Synovial sheaths of the carpal flexor tendons
Two synovial sheaths envelop the flexor tendons as they traverse the carpal tunnel, one for the flexores digitorum superficialis and profundus, the other for flexor pollicis longus (Fig. 50.25). These sheaths typically extend into the forearm for 2.5 cm proximal to the flexor retinaculum, and occasionally communicate with each other deep to it. The sheath of the flexores digitorum tendons reaches about halfway along the metacarpal bones, where it ends in blind diverticula around the tendons to the index, middle and ring fingers (Fig. 50.26). The sheath is prolonged around the tendons to the little finger and is usually continuous with their digital synovial sheath. A transverse section through the carpus shows that the tendons are invaginated into the sheath from the lateral side (Fig. 50.25). The parietal layer lines the flexor retinaculum and the floor of the carpal tunnel. It is reflected laterally as the visceral layer over the tendons of flexor digitorum superficialis ventrally and flexor digitorum profundus dorsally. Medially a recess formed by the visceral layer of the sheath insinuates between the two groups of tendons and passes laterally for a variable distance. The sheath of flexor pollicis longus, which is usually separate, is continued along the thumb as far as the insertion of the tendon.
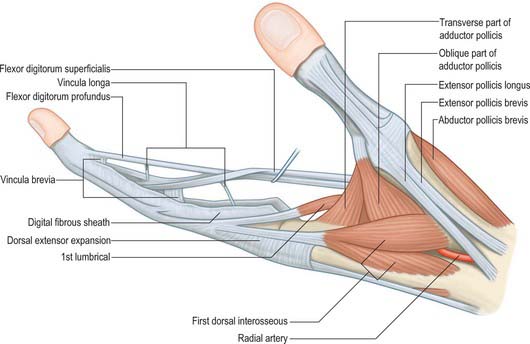
Vincula
The phenomenon of tendon gliding within a fibrous sheath requires a very specialized arrangement of the vascular supply. Folds of synovial membrane containing a loose plexus of fascial fibres carry blood vessels to the tendons at certain defined points (Fig. 50.28). These folds, vincula tendinum, are of two kinds. Vincula brevia, of which there are two in each finger, are attached to the deep surfaces of the tendons near to their insertions. There is thus one vinculum brevium attaching flexor digitorum profundus to the region of the distal interphalangeal joint, and a more proximal vinculum deep to flexor digitorum superficialis at the proximal interphalangeal joint. Vincula longa are filiform: usually two are attached to each superficial tendon, one to each deep tendon.
Extensor retinaculum
The extensor retinaculum is a strong, fibrous band which extends obliquely across the back of the wrist (Fig. 50.29). It is attached laterally to the anterior border of the radius, medially to the triquetral and pisiform bones, and, in passing across the wrist, to the ridges on the dorsal aspect of the distal end of the radius. It prevents bowstringing of the tendons across the wrist joint.
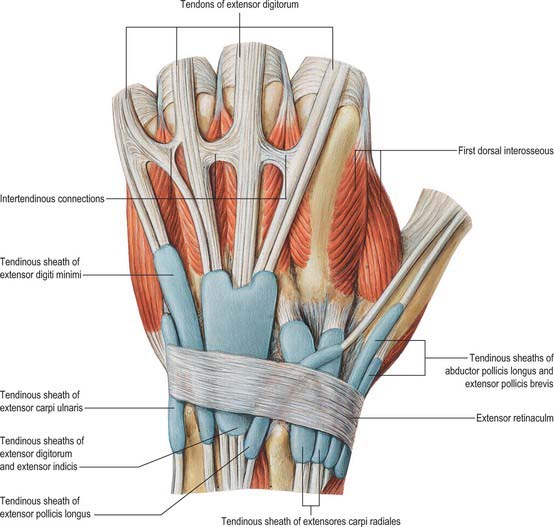
Fig. 50.29 The synovial sheaths of the tendons on the extensor aspect of the left wrist.
(From Sobotta 2006.)
Synovial sheaths of the carpal extensor tendons
De Quervain’s tenovaginitis is a stenosing tenovaginitis of unknown aetiology which occurs in the first dorsal compartment at the level of the radial styloid. It involves the common extensor sheath containing the tendons of abductor pollicis longus and extensor pollicis brevis. There is palpable thickening of the tendon sheath with painful limitation of extension of the thumb. Treatment frequently requires division of the thickened sheath, care being taken to avoid the adjacent superficial radial nerve. Division of the sheath produces no functional impairment.
Extensor tendon apparatus
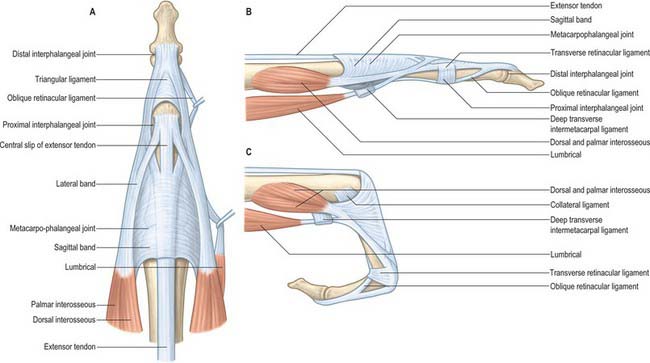
At the level of the metacarpophalangeal joint, each extensor tendon is held in a central position over the dorsum of the joint by a flat fibrous extensor expansion (Fig. 50.30). The expansion extends onto the dorsum of the proximal phalanx of each digit. It forms a movable hood, which moves distally when the metacarpophalangeal joint is flexed, and proximally when it is extended (in which position it is most closely applied to the joint).
The margins of the extensor expansions are thickened on the radial side by the tendons of lumbrical and interosseous muscles and on the ulnar side by the tendon of an interosseous alone or, in the case of the fifth digit, by abductor digiti minimi.
INTRINSIC MUSCLES OF THE HAND
Flexor pollicis brevis
Flexor pollicis brevis lies medial to abductor pollicis brevis (Fig. 50.31). It has superficial and deep parts. The superficial head arises from the distal border of the flexor retinaculum and the distal part of the tubercle of the trapezium, and passes along the radial side of the tendon of flexor pollicis longus. It is attached, by a tendon that contains a sesamoid bone, to the radial side of the base of the proximal phalanx of the thumb. The deep part arises from the trapezoid and capitate bones and from the palmar ligaments of the distal row of carpal bones, and passes deep to the tendon of flexor pollicis longus. It unites with the superficial head on the sesamoid bone and base of the first phalanx.
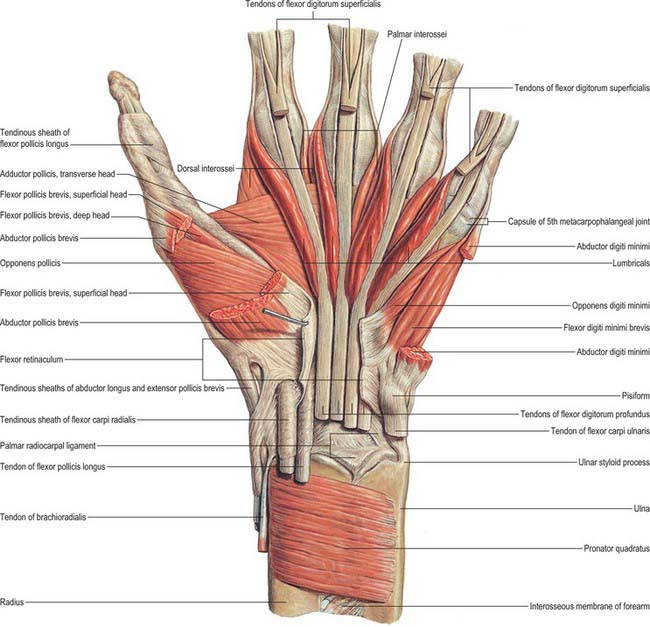
Abductor pollicis brevis
Abductor pollicis brevis is a thin, subcutaneous muscle in the proximolateral part of the thenar eminence (Fig. 50.31). It arises mainly from the flexor retinaculum, but a few fibres spring from the tubercles of the scaphoid bone and trapezium, and from the tendon of abductor pollicis longus. Its medial fibres are attached by a thin, flat tendon to the radial side of the base of the proximal phalanx of the thumb, and its lateral fibres join the dorsal digital expansion of the thumb. The muscle may receive accessory slips from the long and short extensors of the thumb, opponens pollicis, or the styloid process of the radius.
Opponens pollicis
Adductor pollicis
Adductor pollicis arises by oblique and transverse heads (Fig. 50.31). The oblique head is attached to the capitate bone, the bases of the second and third metacarpal bones, the palmar ligaments of the carpus and the sheath of the tendon of flexor carpi radialis. Most of the fibres converge into a tendon (which contains a sesamoid bone), which unites with the tendon of the transverse head, and is attached to the ulnar side of the base of the proximal phalanx of the thumb. The deepest fibres may pass into the medial side of the dorsal digital expansion of the thumb. On the lateral side of the oblique head a considerable fasciculus passes deep to the tendon of flexor pollicis longus to join flexor pollicis brevis; this has been described as the ‘deep head’ of flexor pollicis brevis. The transverse head is the deepest of the pollicial muscles, it is triangular, and arises from the distal two-thirds of the palmar surface of the third metacarpal. The fibres converge to be attached, with the oblique head and the first palmar interosseous, to the base of the proximal phalanx of the thumb. The two parts of the adductor vary in relative size and degree of connection.
Abductor digiti minimi
Abductor digiti minimi arises from the pisiform bone, the tendon of flexor carpi ulnaris and the pisohamate ligament (Fig. 50.31). It ends in a flat tendon which divides into two slips: one is attached to the ulnar side of the base of the proximal phalanx of the little finger, and the other to the ulnar border of the dorsal digital expansion of extensor digiti minimi.
Flexor digiti minimi brevis
Opponens digiti minimi
Opponens digiti minimi flexes the fifth metacarpal bone, drawing it forwards and rotating it laterally at the carpometacarpal joint: this deepens the hollow of the palm. These actions, together with flexion and some lateral rotation at the metacarpophalangeal and interphalangeal joints, bring the digit into opposition with the thumb.
Palmaris brevis
Palmaris brevis is a thin, quadrilateral muscle, lying beneath the skin of the ulnar side of the palm (see Fig. 50.4). It arises from the flexor retinaculum and the medial border of the central part of the palmar aponeurosis, and is attached to the dermis on the ulnar border of the hand.
Interossei
Palmar interossei
Palmar interossei are smaller than dorsal interossei and lie on the palmar surfaces of the metacarpal bones rather than between them (Fig. 50.32). With the exception of the first, each of the three arises from the entire length of the metacarpal bone of one finger, and passes to the appropriate (adductor) side of the dorsal digital expansion.
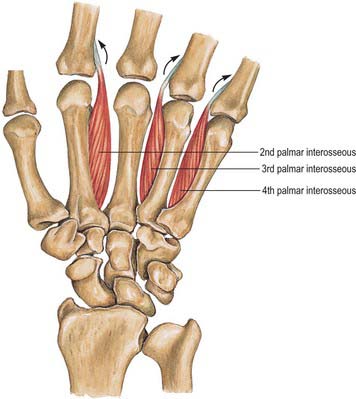
The middle finger has no palmar interosseus. The remaining digits have palmar interossei on their aspects which face the middle finger. The first arises from the ulnar side of the palmar surface of the base of the first metacarpal bone, and is inserted into a sesamoid bone on the ulnar side of the proximal phalanx and from there passes to the phalanx and usually also into the dorsal digital expansion. It lies in front of the lateral head of the first dorsal interosseous, and is overlapped anteriorly by the oblique head of adductor pollicis (Fig. 50.31, Fig. 50.32). It is often very rudimentary because the thumb has its own powerful adductor. The second arises from the ulnar side of the second metacarpal bone, and is inserted into the same side of the digital expansion of the index finger. The third arises from the radial side of the fourth metacarpal bone, and is inserted together with the third lumbrical. The fourth arises from the radial side of the fifth metacarpal bone, and is attached with the fourth lumbrical and also to the base of the proximal phalanx. The attachment of these muscles to the dorsal digital expansions (Fig. 50.30) stabilizes the extensor tendons on the convex heads of the metacarpal bones during flexion and extension at the metacarpophalangeal joints.
The interossei show little variation in their arrangement. They are occasionally reduplicated.
Dorsal interossei
Dorsal interossei consist of four bipennate muscles, each arising from the adjacent sides of two metacarpal bones, but more extensively from the metacarpal bone of the finger into which the muscle passes (Fig. 50.33). They insert on the bases of the proximal phalanges and separately into the dorsal digital expansions. Between the double origin of each of these muscles there is a narrow triangular interval. The radial artery passes through the first of these intervals, and a perforating branch from the deep palmar arch passes through each of the others. The first and largest muscle is sometimes named abductor indicis. It is attached to the radial side of the proximal phalanx of the index finger and to the capsule of the adjoining metacarpophalangeal joint. The second and third are attached to the radial and ulnar sides of the middle finger, respectively. Whereas the second generally reaches the digital expansion and the proximal phalanx, the third usually extends only to the digital expansion (Fig. 50.30). The fourth may be wholly attached to the digital expansion, but it often sends an additional slip to the proximal phalanx.
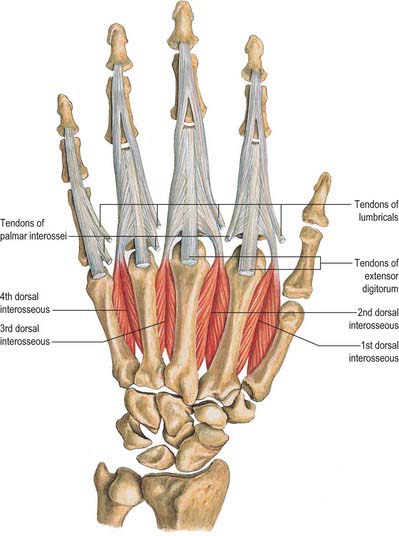
Dorsal interossei are supplied by the dorsal metacarpal arteries (1st–4th), palmar metacarpal arteries (2nd–4th), radial artery (1st); princeps pollicis artery, radialis indicis artery, three perforating branches from the deep palmar arch (proximal perforating arteries), and three distal perforating branches. The tendons are supplied by branches from the common and proper palmar digital arteries and the dorsal digital arteries.
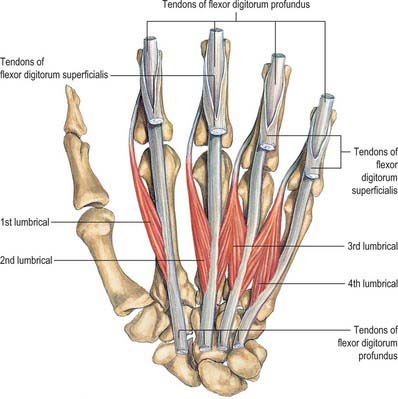
Lumbricals
The lumbricals are four small fasciculi which arise from the tendons of flexor digitorum profundus (Fig. 50.34). The first and second arise from the radial sides and palmar surfaces of the tendons of the index and middle fingers respectively. The third arises from the adjacent sides of the tendons of the middle and ring fingers, and the fourth from the adjoining sides of the tendons of the ring and little fingers. Each passes to the radial side of the corresponding finger, and is attached to the lateral margin of the dorsal digital expansion of extensor digitorum which covers the dorsal surface of the finger.
COORDINATED MOVEMENTS OF THE HAND
The apparently simple human functions of closing the hand to grasp an object, or opening the palm to release it, are in reality tasks of considerable mechanical complexity, requiring the simultaneous contraction of many individual muscles. The isolated action of a single muscle may be inferred from the positions of its origin and insertion, and the estimated line of action (usually the centre line of the muscle) in relation to the axes of all the joints traversed by the muscle and its tendon. The limb can be regarded as a chain of joints crossed by muscles. If it is known which muscles are active, then the reason why one joint moves and others do not is a matter of simple mechanical relationships.
SPECIAL FUNCTIONS OF THE HAND
Closing the hand
Role of the long digital flexors
Flexor digitorum profundus reaches further (to the distal phalanx), and is therefore the only muscle available for flexion of the distal interphalangeal joint. It also contributes, together with superficialis, to flexion at the proximal interphalangeal and metacarpophalangeal joints. These two long flexors (sometimes called extrinsic flexors, because the muscle bellies are outside the hand) can be considered to act together to flex the finger. However, their action alone would wind up the interphalangeal before the metacarpophalangeal joints and the finger would not move in a normal arc of flexion. This is precisely what happens in an ulnar nerve paralysis, in which the interossei and lumbricals are not functioning. These small (intrinsic) muscles have been described earlier in terms of their individual actions. For their role in coordinated activity it is sufficient to appreciate that their contribution changes the arc produced by the long flexors, increasing flexion at the metacarpophalangeal joint and reducing flexion at the proximal interphalangeal joint. All three joints are then angulated to the same degree and the fingers form a normal arc of flexion. As the finger flexes, the long extensor tendons (extensor digitorum, extensor indicis and extensor digiti minimi) aid the process by relaxing and allowing the extensor apparatus to glide distally on the dorsum of the phalanges. (See also p. 879.)
Role of the wrist
For movement about major axes, the wrist tendons can be considered to act in pairs: flexores carpi radialis and carpi ulnaris produce wrist flexion; extensores carpi radialis longus and brevis, and extensor carpi ulnaris produce wrist extension; extensor carpi ulnaris and flexor carpi ulnaris produce ulnar deviation; flexor carpi radialis, extensores carpi radialis longus and brevis, extensor policis and abductor policis longus produce radial deviations. (See also p. 871.)
Opening the hand
One final component of the extensor apparatus provides an additional automatic function. This is a fibrous anchorage system, Landsmeer’s oblique retinacular ligament, which anchors the distal expansion to the middle phalanx. The role of the oblique retinacular ligament is controversial (reviewed by Bendz 1985). Some argue it may act in a dynamic tenodesis effect to synchronize the movements of the interphalangeal joints, i.e. it may initiate extension of the distal interphalangeal joint as the proximal interphalangeal joint is extended from a fully flexed position, and relax with proximal interphalangeal joint flexion to allow full distal interphalangeal joint flexion. Others argue that it only becomes taut when the proximal interphalangeal joint is fully extended and the distal interphalangeal joint is flexed, so that it functions as a restraining force to stabilize the fingertip when it is flexed against resistance, e.g. in the hook grip. A further suggestion is that the ligament is merely a secondary lateral stabilizer of the proximal interphalangeal joint and that it acts to centralize the extensor components over the dorsum of the middle phalanx.
Movements of the thumb
Flexion and extension should be confined to motion at the interphalangeal or metacarpophalangeal joints (Fig. 50.35A–C). Palmar abduction (Fig. 50.35D,E), in which the first metacarpal moves away from the second at right angles to the plane of the palm, and radial abduction (Fig. 50.35D,F), in which the first metacarpal moves away from the second with the thumb in the plane of the palm, occur at the carpometacarpal joint. The opposite of radial abduction is ulnar adduction, or transpalmar adduction, in which the thumb crosses the palm towards its ulnar border. In clinical practice the term adduction is generally used without qualification. Circumduction describes the angular motion of the first metacarpal, solely at the carpometacarpal joint, from a position of maximal radial abduction in the plane of the palm towards the ulnar border of the hand, maintaining the widest possible angle between the first and second metacarpals (Fig. 50.35G). Lateral inclinations of the first phalanx maximize the extent of excursion of the circumduction arc. Opposition is a composite position of the thumb achieved by circumduction of the first metacarpal, internal rotation of the thumb ray and maximal extension of the metacarpophalangeal and interphalangeal joints (Fig. 50.35H). Retroposition is the opposite to opposition (Fig. 50.35I). Flexion and adduction is the position of maximal transpalmar adduction of the first metacarpal: the metacarpophalangeal and interphalangeal joints are flexed and the thumb is in contact with the palm (Fig. 50.35J).
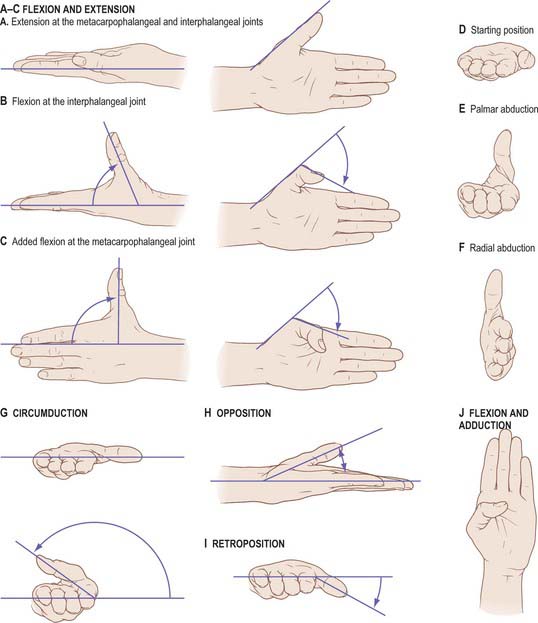
Rotary movements occur during circumduction. The simple angular movements described above combine with rotation about the long axis of the metacarpal shaft. In opposition, the shaft must rotate medially into pronation. In retroposition, the thumb must rotate laterally into supination. Axial rotation of the thumb metacarpal is produced by muscle activity (which moves the thumb through its arc of circumduction); the geometry of the articular surfaces of the trapeziometacarpal joint; tensile forces in the ligaments (which combine with forces exerted by the muscles of opposition and retroposition to produce axial rotation). The stability of the first metacarpal is greatest after complete pronation in the position of full opposition, when ligamentary tension, muscular contraction and joint congruence combine to maximal effect. (See also p. 875.)
Grips
From different positions of the arc of circumduction, numerous different types of pinch grip are possible (Fig. 50.36). In clinical practice these have been classified into two main types: tip pinch and lateral (or key) pinch. Many forces contribute to these configurations.
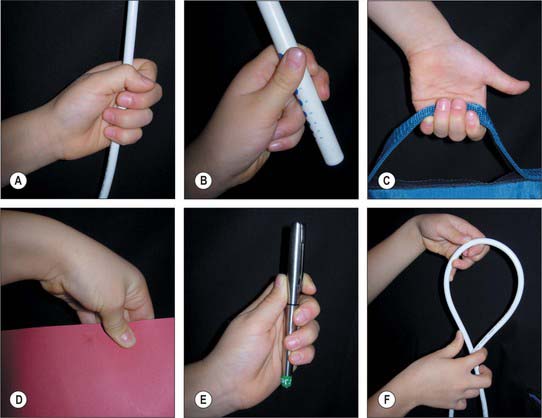
The thumb muscles can be classified into those used for retroposition, opposition and pinch grip.
Pinch grip muscles
The muscles of pinch grip can be divided into lateral, medial and intermediate subgroups. The lateral subgroup (opposition muscles) moves the first metacarpal into palmar abduction. The metacarpal shaft rotates medially into pronation. Radial angulation at the metacarpophalangeal joint increases the span of the hand. The metacarpophalangeal joint is stabilized principally by extensor pollicis brevis and flexor pollicis brevis. Flexion of the proximal and distal phalanges is controlled. Muscles of the medial subgroup (abductor pollicis brevis and first dorsal interosseous) produce an approach of the first metacarpal towards the palm. Since they act with the lateral group they have a strong controlling effect on the position and rotation of the first metacarpal. The intermediate subgroup consists simply of flexor pollicis longus, which flexes the interphalangeal or metacarpophalangeal joint, as described earlier. Palpating the thenar eminence during tip and lateral pinch provides some appreciation of the action of the pinch grip muscles.
VASCULAR SUPPLY AND LYMPHATIC DRAINAGE
ARTERIES
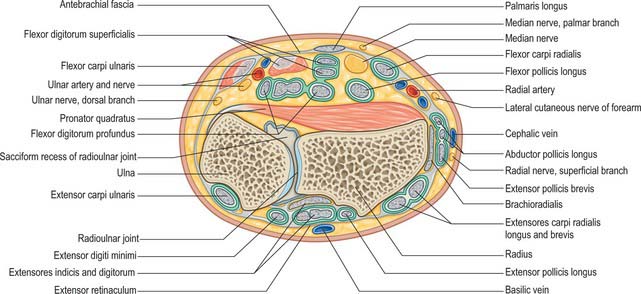
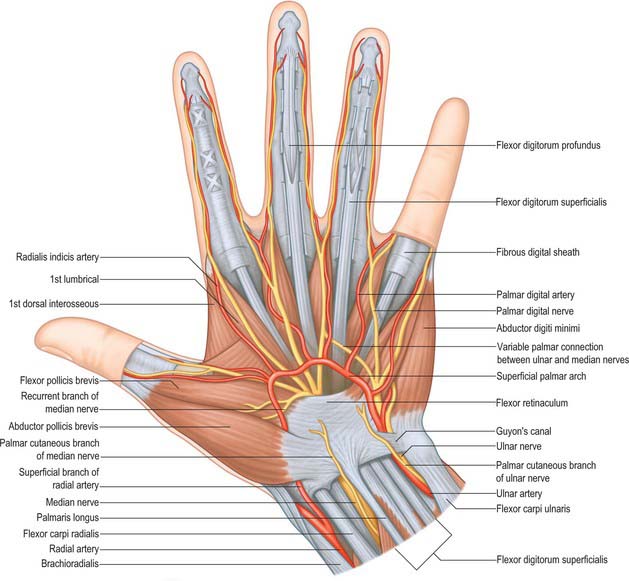
Radial artery
At the wrist the radial artery passes on to the dorsal aspect of the carpus between the lateral carpal ligament and the tendons of abductor pollicis longus and extensor pollicis brevis (Fig. 50.37). It crosses the scaphoid bone and trapezium (in the ‘anatomical snuff-box’), where again its pulsation is obvious, and as it passes between the heads of the first dorsal interosseous it is crossed by the tendon of extensor pollicis longus. Between the thumb extensors it is crossed by the start of the cephalic vein and the digital branches of the radial nerve which supply the thumb and index finger (Fig. 50.38). Occasionally it gives off a distal superficial dorsal branch which crosses the radial extensor tendons at the wrist together with the superficial radial nerve. Filaments of the lateral cutaneous nerve of the forearm run along its distal part as it curves round the carpus.
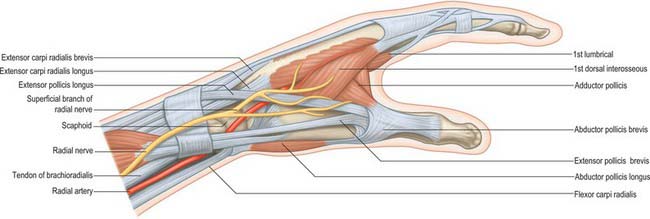
In the hand the radial artery passes through the first interosseous space between the heads of the first dorsal interosseous and crosses the palm. At first it lies deep to the oblique head of adductor pollicis and then passes between its oblique and transverse heads or through the transverse head. At the fifth metacarpal base it anastomoses with the deep branch of the ulnar artery, completing the deep palmar arch (Fig. 50.39).
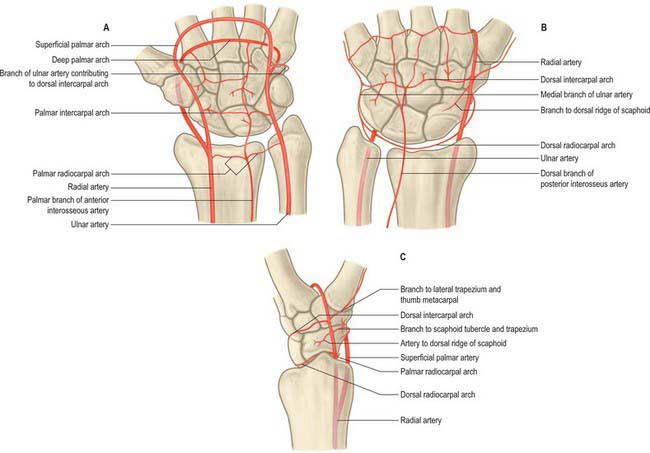
The palmar carpal branch is a small vessel which arises from the radial artery near the distal border of pronator quadratus (Fig. 50.40). It crosses the anterior surface of the distal end of the radius, near the palmar carpal surface, and passes medially to anastomose behind the long flexor tendons with the palmar carpal branch of the ulnar artery. This transverse anastomosis is joined by longitudinal branches from the anterior interosseous artery and recurrent branches from the deep palmar arch, to form a cruciate palmar carpal arch which supplies the carpal articulations and bones by descending branches. (Although so named, this anastomosis is usually sited near the wrist joint on the distal forearm bones.)
The superficial palmar branch arises from the radial artery just before it curves round the carpus. It passes through, and occasionally over, the thenar muscles, which it supplies, sometimes anastomosing with the end of the ulnar artery to complete a superficial palmar arch (Fig. 50.39).
First dorsal metacarpal artery
The first dorsal metacarpal artery arises from the radial artery just before it passes between the heads of the first dorsal interosseous (Fig. 50.2). It divides almost at once into two branches which supply the adjacent sides of the thumb and index finger as far distally as the proximal interphalangeal joint. The radial side of the thumb receives a branch direct from the radial artery proper.
The usual anatomical arrangement allows a flap of skin over the dorsum of the proximal phalanx of the index finger to be raised on this artery and its accompanying venae comitantes. This is particularly useful undercertain circumstances for reconstruction of the thumb following injury.
Second, third and fourth dorsal metacarpal arteries
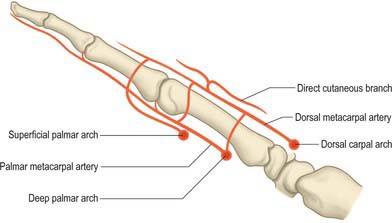
Cutaneous branches from the dorsal metacarpal arteries supply the dorsal skin as far distally as the proximal interphalangeal joint. At the level of the neck of the second, third and fourth metacarpals, a direct cutaneous branch is given off which passes proximally and supplies an area of skin between the two adjacent metacarpals (Fig. 50.41).
The arteria princeps pollicis arises from the radial artery as it turns into the palm to form the deep palmar arch. It descends on the palmar aspect of the first metacarpal under the oblique head of adductor pollicis lateral to the first palmar interosseous. At the base of the proximal phalanx, deep to the tendon of flexor pollicis longus, the artery divides into two branches. It appears between the medial and lateral attachments of the oblique head of adductor pollicis to run along both sides of the thumb. On the palmar surface of its distal phalanx it forms a pollicial arch which supplies the skin and subcutaneous tissue. The arteria princeps pollicis is the usual nutrient of supply to the first metacarpal bone.
The arteria radialis indicis is often a proximal branch of the arteria princeps pollicis. It descends between the first dorsal interosseous and transverse head of adductor pollicis, and along the lateral side of the index finger to its end (Fig. 50.37). It anastomoses with the indicial medial digital artery. At the distal border of the transverse head of adductor pollicis it anastomoses with the arteria princeps pollicis and links with the superficial palmar arch. It may arise from the superficial arch or from the first dorsal metacarpal artery.
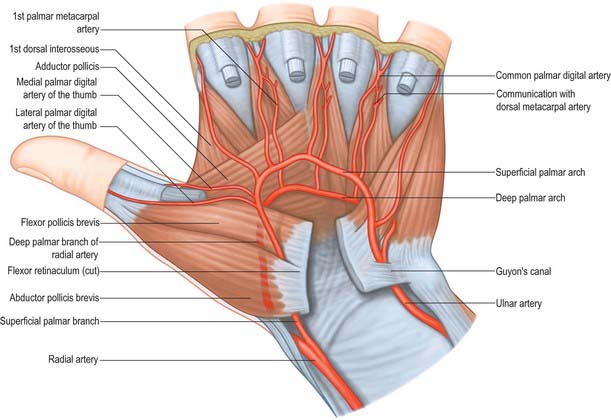
The deep palmar arch is formed by anastomosis of the end of the radial artery with the deep palmar branch of the ulnar artery (Fig. 50.39 and Fig. 50.42A). It crosses the bases of the metacarpal bones and interossei, covered by the oblique head of adductor pollicis, the digital flexor tendons and lumbricals. In its concavity, running laterally, is the deep branch of the ulnar nerve. Rarely the arch is incomplete. There are variations in the size of contribution from the ulnar artery.
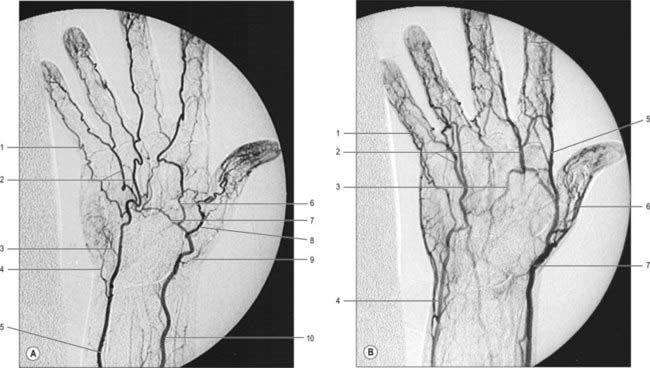
The three palmar metacarpal arteries run distally from the convexity of the deep palmar arch on the interossei of the second to fourth spaces, and join the common digital branches of the superficial arch at the digital clefts (Fig. 50.39). They supply nutrient branches to the medial four metacarpals.
Ulnar artery
At the wrist the ulnar artery (Figs 50.37 and 50.39) is covered by skin, fasciae and palmaris brevis. It lies between the superficial and main parts of the flexor retinaculum, lateral to the ulnar nerve and pisiform.
A small palmar carpal branch crosses the distal ulna deep to the tendons of flexor digitorum profundus (Fig. 50.40), and anastomoses with a palmar carpal branch of the radial artery to make a palmar carpal arch.
A dorsal carpal branch arises just proximal to the pisiform (Fig. 50.40). It curves deep to the tendon of flexor carpi ulnaris to reach the carpal dorsum, which it crosses laterally beneath the extensor tendons. It anastomoses with the dorsal carpal branch of the radial artery to complete the dorsal carpal arch. Near its origin it sends a small digital branch along the ulnar side of the fifth metacarpal to supply the medial side of the dorsal surface of the fifth finger.
Superficial palmar arch
The superficial palmar arch is an anastomosis fed mainly by the ulnar artery (Figs 50.2 and 50.39). The latter enters the palm with the ulnar nerve, anterior to the flexor retinaculum and lateral to the pisiform. It passes medial to the hook of the hamate, then curves laterally to form an arch that is convex distally and level with a transverse line through the distal border of the fully extended pollicial base. About a third of the superficial palmar arches are formed by the ulnar artery alone; a further third are completed by the superficial palmar branch of the radial artery; and a third by the arteria radialis indicis, a branch of either arteria princeps pollicis or the median artery. The superficial palmar arch is covered by palmaris brevis and the palmar aponeurosis and it is superficial to flexor digiti minimi, branches of the median nerve and the long flexor tendons and lumbricals.
Common and proper palmar digital arteries
Three common palmar digital arteries arise from the convexity of the superficial palmar arch (Fig. 50.39). They pass distally on the second to fourth lumbricals, each joined by a corresponding palmar metacarpal artery from the deep palmar arch, and divide into two proper palmar digital arteries. These run along the contiguous sides of all four fingers, dorsal to the digital nerves, between Grayson’s and Cleland’s ligaments, anastomosing in the subcutaneous tissue of the finger tips and near the interphalangeal joints. Each digital artery has two dorsal branches which anastomose with the dorsal digital arteries and supply the soft parts dorsal to the middle and distal phalanges, including the matrices of the nails. The palmar digital artery for the medial side of the little finger leaves the arch under palmaris brevis. Palmar digital arteries supply the metacarpophalangeal and interphalangeal joints and nutrient rami to the phalanges. They are the main digital supply, because the dorsal digital arteries are minute.
The terminal branches of the digital arteries contribute to a number of vascular arcades which provide a rich vascular supply for the distal elements of each digit (Fig. 50.43). Three distal phalangeal dorsal arterial arcades anastomose with each other and with those from the other side of the digit. The superficial arcade occurs at the level of the proximal nailfold and is supplied primarily by a dorsal branch from the palmar digital artery, which is given off at the level of the middle phalanx. The proximal subungual arcade is at the level of the lunula and is supplied by a terminal branch of the digital artery, which passes dorsally. The distal subungual arcade occurs more distally in the nail bed. It is supplied by a dorsal vessel which emerges from the point of confluence of the ‘H’-shaped anastomosis between the terminal portions of both digital arteries, and passes from volar to dorsal aspect of the digit under the interosseous ligament which connects the proximal and distal parts of the distal phalanx (Fig. 50.43).
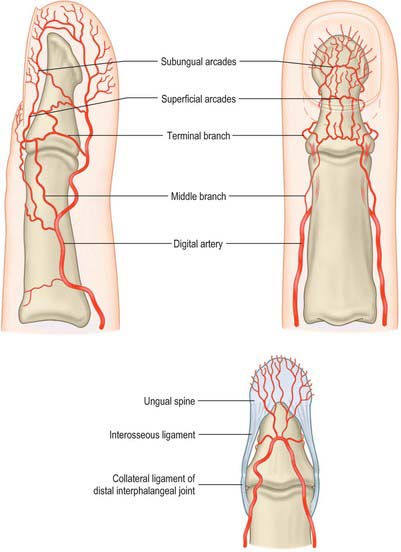
Glomus tumours are very painful (often tiny and sometimes difficult to identify) tumours of the glomus bodies (small AV anastomoses involved in the regulation of peripheral skin temperature control; see Fig. 6.16). These typically occur in the proximal nailfold/subungual regions of the finger tips in association with the dorsal digital arterial arcades, although they can occur anywhere. They are diagnosed by exquisite point tenderness over the swelling with reduction of tenderness when the finger is exsanguinated. If the tumour is not visible (as is often the case) then the best way surgically to explore the dorsal fingertip region to look for the tumour is to incise the lateral nailfold and hyponychium and raise this as a flap along with the underlying periosteum pedicled on the blood supply from the opposite nailfold. The dorsal distal phalangeal digital arterial arcades are then identified from their deep surface and any glomus tumour identified.
VEINS
Superficial veins of the hand
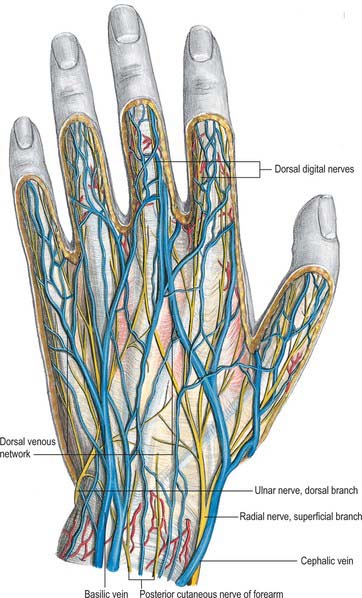
Dorsal and palmar digital veins
Dorsal digital veins pass along the sides of the fingers, joined by oblique branches. They unite from the adjacent sides of the digits into three dorsal metacarpal veins (Fig. 50.44), which form a dorsal venous network over the metacarpus. This is joined laterally by a dorsal digital vein from the radial side of the index finger and both dorsal digital veins of the thumb, and is prolonged proximally as the cephalic vein (Fig. 50.42). Medially a dorsal digital vein from the ulnar side of minimus joins the network, which ultimately drains proximally into the basilic vein. A vein often connects the central parts of the network to the cephalic vein near the midforearm.
INNERVATION
Median nerve
The median nerve proximal to the flexor retinaculum is lateral to the tendons of flexor digitorum superficialis and lies between the tendons of flexor carpi radialis and palmaris longus (Fig. 50.45A). It passes under the retinaculum in the ‘carpal tunnel’ where it may be compressed in the carpal tunnel syndrome (see p. 897). Distal to the retinaculum the nerve enlarges and flattens, and usually divides into five or six branches: the mode and level of division are variable.
Muscular branch (motor or recurrent branch)
The muscular branch is short and thick, and arises from the lateral side of the nerve; it may be the first palmar branch or a terminal branch which arises level with the digital branches. It runs laterally, just distal to the flexor retinaculum, with a slight recurrent curve beneath the part of the palmar aponeurosis covering the thenar muscles (Fig. 50.45A). It turns round the distal border of the retinaculum to lie superficial to flexor pollicis brevis, which it usually supplies, and continues either superficial to the muscle or traverses it. It gives a branch to abductor pollicis brevis, which enters the medial edge of the muscle, and then passes deep to it to supply opponens pollicis, entering its medial edge. Its terminal part occasionally gives a branch to the first dorsal interosseous, and may be its sole or partial supply. The muscular branch may arise in the carpal tunnel and pierce the flexor retinaculum, a point of surgical importance.
Palmar digital branches
The median nerve usually divides into four or five digital branches (Fig. 50.45A). It often divides first into a lateral ramus which provides digital branches to the thumb and the radial side of the index finger, and a medial ramus, which supplies digital branches to adjacent sides of the index, middle and ring fingers. Other modes of termination can occur.
Ulnar nerve
Dorsal branch
The dorsal branch arises approximately 5 cm proximal to the wrist. It passes distally and dorsally, deep to flexor carpi ulnaris, perforates the deep fascia, descends along the medial side of the back of the wrist and hand and then divides into two, or often three, dorsal digital nerves. The first supplies the medial side of the little finger, the second, the adjacent sides of the little and ring fingers, while the third, when present, supplies adjoining sides of the ring and middle fingers. The latter may be replaced, wholly or partially, by a branch of the radial nerve, which always communicates with it on the dorsum of the hand (Fig. 50.45A). In the little finger, the dorsal digital nerves extend only to the base of the distal phalanx, and in the ring finger they extend only to the base of the middle phalanx. The most distal parts of the little finger and of the ulnar side of the ring finger are supplied by dorsal branches of the proper palmar digital branches of the ulnar nerve. The most distal part of the lateral side of the ring finger is supplied by dorsal branches of the proper palmar digital branch of the median nerve.
Superficial and deep terminal branches
Deep terminal branch
The deep terminal branch accompanies the deep branch of the ulnar artery as it passes between abductor digiti minimi and flexor digiti minimi and then perforates opponens digiti minimi to follow the deep palmar arch dorsal to the flexor tendons (Fig. 50.46). At its origin it supplies the three short muscles of the little finger. As it crosses the hand, it supplies the interossei and the third and fourth lumbricals. It ends by supplying adductor pollicis, the first palmar interosseous and usually flexor pollicis brevis. It sends articular filaments to the wrist joint.
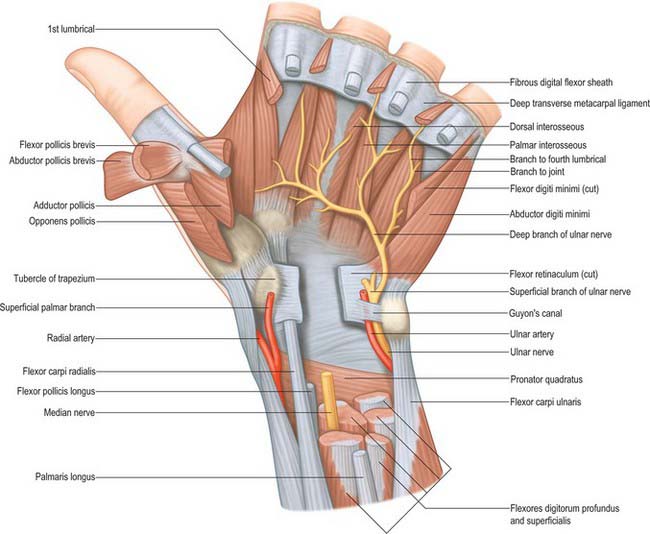
The medial part of flexor digitorum profundus is supplied by the ulnar nerve, as are the third and fourth lumbricals which are connected with the tendons of this part of the muscle. Similarly, the lateral part of flexor digitorum profundus and the first and second lumbricals are supplied by the median nerve. The third lumbrical is often supplied by both nerves. The deep terminal branch is said to give branches to some intercarpal, carpometacarpal and intermetacarpal joints: precise details are uncertain. Vasomotor branches, arising in the forearm and hand, supply the ulnar and palmar arteries.
Ulnar nerve division at the wrist
Division of the ulnar nerve at the wrist paralyses all the intrinsic muscles of the hand (apart from the radial two lumbricals). The intrinsic muscle action of flexing the metacarpophalangeal joint and extending the interphalangeal joints is therefore lost. The unopposed action of the long extensors and flexors of the fingers cause the hand to assume a clawed appearance with extension of the metacarpophalangeal joints and flexion of the interphalangeal joints. The clawing is less intense in the index and middle fingers because of their intact lumbricals, supplied by the median nerve. (For a detailed account of the hand posture adopted in ulnar nerve lesions, see Smith 2002.)
An K-N, Berger RA, Cooney WP. Biomechanics of the Wrist Joint. New York: Springer-Verlag, 1991.
Bendz P. The functional significance of the oblique retinacular ligament of Landsmeer. A review and new proposals. J Hand Surg. 1985;10:25-29.
Bowers WH, Wolf JWJr, Nehil JL, Bittinger S. The proximal interphalangeal joint volar plate. I. An anatomical and biochemical study. J Hand Surg [Am]. 1980;5:79-88.
Cormack GC, Lamberty BGH. The Arterial Anatomy of Skin Flaps, 2nd edn. Edinburgh: Churchill Livingstone, 1994.
Doyle JR, Blythe WF. The finger flexor tendon sheath and pulleys: anatomy and reconstruction. In: AAOS Symposium on Tendon Surgery in the Hand. St Louis: Mosby; 1975:81-88.
Garcia-Elias M. Carpal kinetics. In: Büchler V, editor. Wrist Instability. London: Martin Dunitz, 1996. Chapter 2.
Green D, Hotchkiss R, Pederson W. Green’s Operative Hand Surgery, 4th edn. New York: Churchill Livingstone, 1999.
Kauer JMG. The functional anatomy of the carpal joint: the whole and its components. In: Wrist Instability. London: Martin Dunitz; 1996.
Kirk Watson H, Weinzweig J. The Wrist. Philadelphia: Lippincott, Williams & Wilkins, 2001.
McGrouther D. The microanatomy of Dupuytren’s contracture. The Hand. 1982;14:215-236.
Salmon M. Anatomic studies. In: Taylor GI, Razaboni RM. Book 1: Arteries of the muscles of the extremities and the trunk. Book 2: Arterial anastomotic pathways of the extremities. St Louis: Quality Medical Publishing, 1994.
Smith PJ. Lister’s The Hand. Diagnosis and Indications. 4th edn.. Edinburgh: Churchill Livingstone; 2002.
Spinner MJB. Kaplan’s Functional and Surgical Anatomy of the Hand, 3rd edn. Philadelphia: Lippincott, Williams & Wilkins, 1984.
Stanley J, Saffar P. Anatomy. In: Wrist Arthroscopy. London: Martin Dunitz; 1994.
Watson HK, Weinzweig J. The Wrist. Philadelphia: Lippincott Williams and Wilkins, 2001.