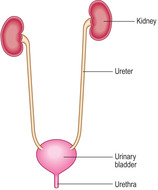
[level-membership-for-basic-science-category]The urinary system consists of paired kidneys and the urinary tract, which comprises paired ureters, a urinary bladder and a urethra (Fig. 13.1). Urine is produced by the kidneys and passes along the ureters to the urinary bladder, where it is stored until it is voided via the urethra. The route taken by urine along the urethra to the exterior is an independent closed system in females, but is shared with the reproductive system in males (Chapter 15).
The urinary system is essential in maintaining the homeostasis of the body. It does this by regulating the water and mineral salts in, and the acid–base balance of, blood. It is particularly important in excreting toxic molecules containing nitrogen (e.g. urea and creatinine) produced by the breakdown of endogenous proteins. The urinary system also ensures that useful molecules in blood, e.g. proteins and carbohydrates, are not lost during the formation of urine. The toxic molecules and excess ions, dissolved in water, leave the kidneys as urine, which passes along the urinary tract before being voided at micturition. In addition, the kidneys produce and secrete into blood two molecules, renin and erythropoietin. The former is important in regulating blood pressure and the latter in stimulating the formation of red blood cells.
Kidneys
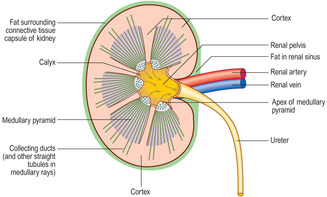
The gross structure of a kidney is best described as seen in a longitudinal hemi-section (
Fig. 13.2). Facing towards the mid-line of the body, each kidney has an indentation forming a hilum. At the hilum blood enters and leaves each kidney in, respectively, a renal artery and a renal vein, and urine drains into a ureter.
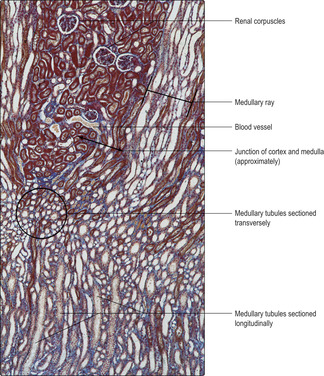
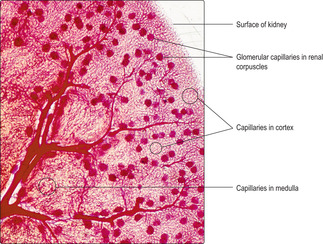
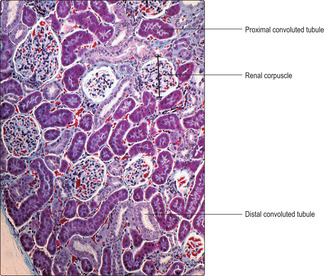
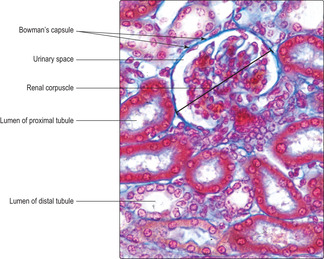
The gross appearance of the kidney displays an outer and an inner region, the cortex and the medulla. The cortex has a granular appearance due to the presence of spherical structures about 200
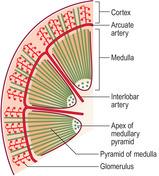
μm in diameter known as renal corpuscles. These corpuscles filter blood in the initial stage in the formation of urine. The cortex also contains convoluted tubules involved in forming urine. Renal corpuscles are not present in the medulla and the medulla appears smooth or may show striations. Straight and arching tubules are present in the medulla and they also are involved in forming urine. Renal corpuscles and tubules may be distinguished in histological sections even at low magnification (Fig. 13.3). In humans, the medulla projects centrally as several pyramids and the cortex extends as columns between the pyramids (Fig. 13.2). The apices of the pyramids project as renal papillae into urine-filled spaces (minor calyces). Urine drains from tubules in the medulla into the calyces. Each minor calyx drains urine into major calyces that together form the renal pelvis. The relatively large, fluid-filled space of each renal pelvis drains into a ureter (Fig. 13.2). Fat cells surround the renal pelvis, the ureter and vessels at the hilum of the kidney and pack a space known as the renal sinus. A dense layer of fat surrounds each kidney.The blood supply to kidneys ensures that the whole volume of blood in the body passes through the kidneys every 5 minutes or so. The arrangement of blood vessels is based on supplying (and draining) each medullary pyramid and its associated cortical tissues separately, a unit described as a renal lobe. (The human kidney is multilobar, but uni-lobar kidneys occur in many species.) Each renal artery branches and forms interlobar arteries. In turn, interlobar arteries branch further and then form arcuate arteries which arch between the cortex and medulla (
Fig. 13.4). Branches from arcuate arteries eventually supply each renal corpuscle with an afferent arteriole. This arrangement is essential in filtering the blood (see below) as well as supplying oxygen and nutrients.
Each afferent arteriole divides within a renal corpuscle into capillaries, known as glomerular capillaries (Fig. 13.5), and these drain from the corpuscle into an efferent arteriole (
Fig. 13.6). This arrangement of capillaries draining to an arteriole is unique to the kidney and results in a relatively high blood pressure in glomerular capillaries (
Fig. 13.5) which aids the filtration of blood. Efferent arterioles divide and form a second network of capillaries. Some of these capillaries are straight vessels, vasa recta, which lie between straight tubules in the kidney whereas others form an extensive meshwork surrounding convoluted tubules in the cortex (
Fig. 13.6). The second set of capillaries supplies oxygen and nutrients to the cells of the kidney and is also involved in modifying the filtrate and forming urine. Veins drain capillaries (other than glomerular capillaries) in the kidney and the routes they take closely follow the arterial pattern.
Nephrons and collecting ducts
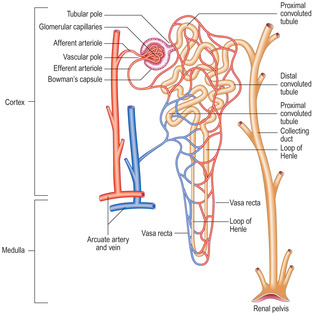
A nephron is the functional unit of the kidney (
Fig. 13.6) and there are about a million in each kidney in humans. A nephron consists of a renal corpuscle, which filters blood, and the uriniferous tubule attached to it which drains and modifies the filtrate. Eventually, the modified filtrate becomes urine and drains, via collecting ducts, into the renal pelvis.
Renal corpuscles
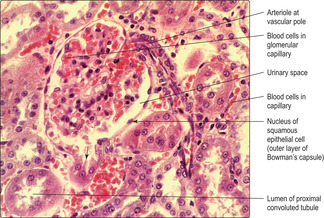
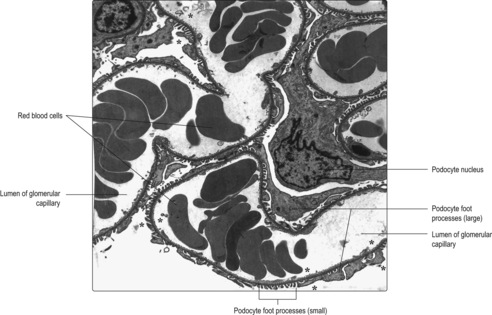
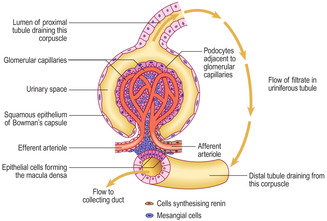
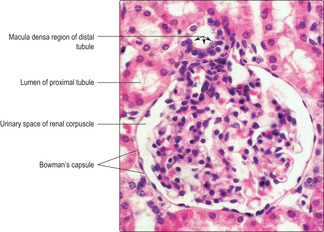
Each renal corpuscle consists of glomerular capillaries (a renal glomerulus) (Fig. 13.5) within a Bowman’s capsule. The capsule is shaped like a hollow, double-walled cup (
Fig. 13.6) and it is lined by epithelial cells. The outer parietal layer of Bowman’s capsule is lined by squamous epithelial cells (
Fig. 13.7) and it is continuous with the inner visceral layer of epithelial cells. The cells of the inner layer, known as podocytes, have numerous cytoplasmic processes which abut the capillaries of the glomerulus. The endothelial cells lining the capillaries and the podocytes (and their fused basement membranes) form the filtration barrier between the blood and the filtrate. The space between the inner and outer layers of the capsule receives the filtrate and is known as the urinary space (
Fig. 13.7) even though the filtrate is not yet urine.
Each Bowman’s capsule has two poles, and it is extremely rare to see all the features of each pole in any one (histological) section of a corpuscle.
■
Vascular pole (
Figs 13.6 and
13.7). At this pole an afferent arteriole enters each renal corpuscle, branches and forms glomerular capillaries. These capillaries rejoin and drain into an efferent arteriole (not a venule), which leaves the corpuscle at the vascular pole.
■
Tubular pole (
Fig. 13.6,
Fig. 13.7 and
Fig. 13.8). At this pole the filtrate drains from the urinary space of the renal corpuscle. It enters the proximal part of the uriniferous tubule, which extends from the parietal layer of Bowman’s capsule at this pole.
Filtration
Blood in glomerular capillaries is filtered through the endothelium lining the capillaries. It then traverses the fused basement membranes of the endothelium and of the adjacent podocytes (the visceral epithelial cells of Bowman’s capsule) (
Fig. 13.9).
Filtration is affected by:
■ the pressure of blood in the afferent arteriole
■ the pressure of blood in the efferent arteriole; this provides resistance to the outflow of blood from the glomerular capillaries causing a relatively high pressure in the glomerular capillaries
■ fenestrations in the endothelium of the glomerular capillaries; these fenestrations retain molecules over about 69 000 daltons in the blood (e.g. albumin)
■ the fused basement membranes; these are charged and repel many protein molecules
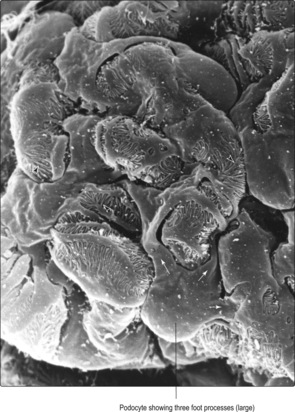
■ the structure of the podocytes. Podocytes have numerous large and small cytoplasmic foot processes (
Figs 13.9 and
13.10). The smaller foot processes abut the basement membrane and there are small spaces between them. It is probable that this arrangement of the foot processes is involved in preventing some proteins from passing from blood into the filtrate.
Uriniferous tubule
Each uriniferous tubule drains the filtrate from the urinary space of a Bowman’s capsule (
Fig. 13.6). The filtrate enters the proximal convoluted tubule, the first part of the uriniferous tubule, and this takes a coiled course in the cortex before straightening. This straight tubule continues and forms a loop (the loop of Henle), which extends from the cortex towards the medulla; many loops of Henle extend deep into the medulla. Each loop of Henle returns to the cortex and continues as a distal convoluted tubule which eventually drains into a straight collecting duct in the cortex (
Fig. 13.6). (Collecting ducts are not usually described as part of the nephron.)
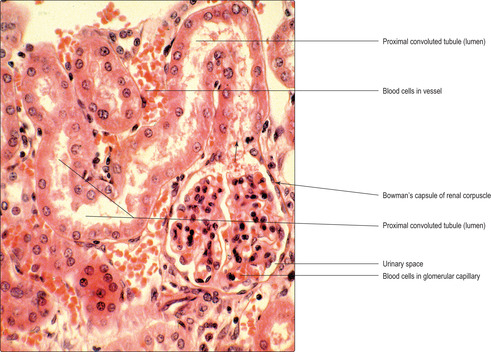
Proximal convoluted tubule
Proximal convoluted tubules are longer than distal convoluted tubules and thus are the predominant tubule seen in histological sections of the kidney cortex (
Fig. 13.11). They are lined by simple cuboidal (or low columnar) epithelial cells (
Figs 13.7 and
13.8). The lumen of proximal tubules is indistinct in many histological sections as the lumen that transports filtrate in life is virtually filled by the apical surface of the epithelial cells.
Proximal convoluted tubules resorb about 80% of the water that has passed into the glomerular filtrate and many of the sodium and chloride ions. This resorption is facilitated by microvilli on the apical (luminal) surfaces of the epithelial cells lining the proximal tubules, providing a large interface between the cell membrane and the filtrate. The movement of sodium ions out of the filtrate is an energy-dependent process powered by mitochondria in the epithelial cells. Water passively follows the sodium ions.
Resorbed water and ions readily pass into blood in the capillaries, which form a meshwork around proximal convoluted tubules (
Figs 13.7,
13.8 and
13.11). The movement of these ions and water helps maintain electrical neutrality and osmotic equilibrium in the capillaries. Amino acids and glucose in the glomerular filtrate are resorbed from the filtrate in proximal tubules and return to blood. Small proteins in the filtrate are taken into the epithelial cells lining proximal tubules by endocytosis. After fusion of the endocytotic vesicles with lysosomes the proteins are digested and the amino acids produced return to blood.
In addition, the epithelial cells lining proximal tubules (in humans) are able to secrete waste molecules such as creatinine into the lumen of the proximal tubules, and they are able to prevent the resorption of some toxins and drugs which have passed from blood into the glomerular filtrate.
Loop of Henle
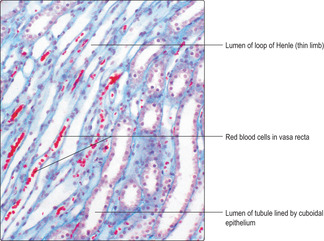
The tubule of the first part of a loop of Henle drains a proximal convoluted tubule and it passes (descends) as a straight tubule towards, and usually into, the medulla. The tubule then loops and passes (ascends) as a straight tubule into the cortex (
Fig. 13.6). The lower part of the descending limb of the loop of Henle and some of the ascending limb are tubules that have relatively wide lumina and thin walls. These ‘thin’ regions are lined by a simple squamous epithelium (
Fig. 13.12). The final portion of the loop of Henle is lined by a simple cuboidal epithelium and it continues, in the cortex, as a distal convoluted tubule which is lined by a similar cuboidal epithelium.
The functions of the various regions of loops of Henle are related to producing a hypertonic environment in the sparse connective tissue (interstitium) between tubules in the medulla (
Fig. 13.3). This is essential for the production of hypertonic urine (see below). The interstitium comprises few cells, e.g. fibroblasts, sparse fibres and complex carbohydrate molecules. Several factors are involved in modulating the tonicity of the interstitium:
■ the majority of the (cuboidal) epithelial cells lining the ascending limb is impermeable to water
■ cuboidal epithelial cells lining the ascending limb actively pump sodium ions into the interstitium and chloride ions follow passively; the hormone aldosterone from the adrenal glands (
Chapter 14) stimulates this resorption of sodium ions
■ some ions in the interstitium diffuse into nearby straight capillaries (vasa recta) (
Fig. 13.12)
■ some ions re-enter the descending limb but may be pumped out again as they pass again along the distal (ascending) limb
■ the net movement of ions results in:
■ a hypertonic interstitium (particularly near the apex of the medullary pyramids)
■ a hypotonic fluid passing into the distal tubule in the cortex.
The term ‘countercurrent multiplier mechanism’ is used to describe this complex movement of ions.
Distal convoluted tubule
Distal convoluted tubules are shorter than proximal convoluted tubules so appear to be fewer in number in histological sections (
Fig. 13.11). They are lined by a simple epithelium consisting of small cuboidal or low columnar cells (
Fig. 13.13) which do not have microvilli. As a result, in comparison with proximal tubules, distal tubules appear to have more distinct lumina and more closely packed cells in their walls (
Fig. 13.13).
Each distal convoluted tubule returns to the vascular pole of the renal corpuscle from which it arose and forms part of a functional unit known as the juxtaglomerular apparatus (see below). Epithelial cells lining distal tubules resorb sodium ions from the filtrate. Resorbed molecules return to blood in nearby capillaries (
Fig. 13.11). In addition, hydrogen ions are secreted. The ion movements in this region are important in regulating the tonicity and acid–base balance of blood. The movements of these ions are affected by the hormone aldosterone from the adrenal gland (
Chapter 14).
Collecting ducts
Several distal convoluted tubules in the cortex drain into each collecting duct (
Fig. 13.6). Collecting ducts are straight tubules which lie clustered in parallel with other straight tubules (e.g. loops of Henle) that pass between the cortex and the medulla. The regions containing parallel tubules are known as medullary rays (
Fig. 13.3). Collecting ducts are lined by a simple cuboidal epithelium and in the medulla are surrounded by the hypertonic interstitium. This aids resorption of water and the production of hypertonic urine. Two hormones are involved in controlling this resorption of water and some ions. Aldosterone stimulates the epithelial cells lining collecting ducts to resorb sodium and chloride ions, and water follows passively. In addition, antidiuretic hormone (from the posterior pituitary gland; see
Chapter 14) controls the permeability of the cells lining collecting ducts and regulates the amount of water lost in urine. Collecting ducts join together and form wider ducts (of Bellini) which drain the urine into minor calyces at the medullary papillae, and thence into the major calyces of the renal pelvis.
Juxtaglomerular apparatus
Each juxtaglomerular apparatus (Fig. 13.14) involves an afferent arteriole supplying blood to glomerular capillaries in a renal corpuscle, the efferent arteriole draining these capillaries and adjacent connective tissue. In addition, a short region of the distal tubule of the uriniferous tubule draining filtrate from that corpuscle and lying adjacent to the vascular pole, is also involved in the juxtaglomerular apparatus.
Cells in the wall of afferent arterioles secrete the hormone renin. Via its effects on the liver and lungs, renin controls the production of a molecule (angiotensin) which in turn stimulates the production of aldosterone by the adrenal glands (
Chapter 14). The epithelial cells lining the distal tubule at the vascular pole appear to be packed together. This region is known as the macula densa (
Figs 13.14 and
13.15). These epithelial cells probably detect changes in sodium concentration in the filtrate from that corpuscle and help control the rate of filtration by affecting renin secretion.
Connective tissue within, and adjacent to, the renal corpuscle is known as mesangial tissue and it is sparse. Mesangial cells around glomerular capillaries and in the region of the juxtaglomerular apparatus are able to contract and may affect blood flow in the region. In addition, they have phagocytic functions which may be involved in maintaining the basement membranes filtering the blood. Erythropoietin, a molecule which stimulates the production of red cells by bone marrow, is produced by kidneys and it is possible that mesangial cells are the source of this hormone.
Urinary tract
Renal pelves and ureters
Urine drains from the ducts of Bellini in the medulla of each kidney into the lumen of the pelvis of the kidney. The renal pelves are lined by transitional epithelium, the specialised layered epithelium (urothelium) lining all the urinary tract. Each pelvis narrows and is continuous with a ureter (
Fig. 13.2).
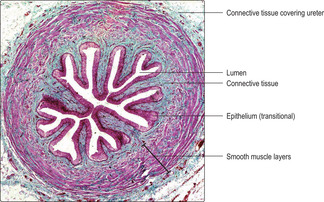
The wall of each ureter is formed from three layers (
Fig. 13.16):
■ An inner lining mucosa. The mucosa comprises the transitional epithelium lining the lumen and the underlying, supporting connective tissue. This specialised epithelium prevents excess ions and toxic molecules in hypertonic urine from diffusing from the urine through the epithelium and returning to circulate in blood.
■ A middle muscularis layer. The muscularis is composed of an outer longitudinal and an inner circular layer of smooth muscle in the upper two-thirds of the ureter. In the lower third, there is an additional outer layer of smooth muscle that spirals around the long axis of the ureter. The muscularis layer undergoes peristaltic contractions and propels urine along the ureter towards and into the bladder.
■ A thin connective tissue covering. The outer connective tissue covering holds each ureter against the adjacent posterior abdominal wall and separates them from overlying peritoneum (the serous membrane of the peritoneal cavity).
Urinary bladder
The urinary bladder is similar to the ureters in that its wall comprises three layers: a mucosa, a muscularis and an outer layer. However, the bladder is able to distend and store large quantities of urine and, after infancy, it is able to contract and expel urine at socially acceptable times.
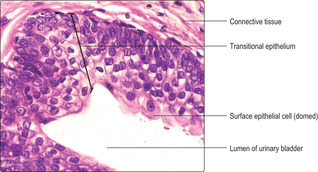
The bladder mucosa is composed of transitional epithelium (urothelium) and a supporting connective tissue lamina propria (
Fig. 13.17). The mucosa is folded and forms rugae in an empty bladder and there are several layers of cells in the epithelium; some surface cells may appear domed (
Fig. 13.17). In addition, in the empty bladder parts of the cell membrane of surface epithelial cells are drawn into the cytoplasm. As the volume of urine stored in the bladder increases the rugae unfold, the number of cell layers in the epithelium decreases and the epithelial cells are flattened and their cell membranes lengthen. The epithelium lining the bladder is impermeable and it ensures that hypertonic urine does not equilibrate with surrounding isotonic tissues and blood, and that toxic molecules, e.g. urea, remain in urine and are excreted.
The lamina propria of the bladder mucosa connects the epithelium to the muscularis layer, which is known as the detrusor muscle. The smooth muscle cells in the detrusor muscle are arranged in three interlacing layers. Contraction of the detrusor muscle is brought about by stimulation of the muscle cells by parasympathetic nerves which are under conscious control after infancy. This contraction empties the bladder as urine is voided via the urethra. Parts of the bladder are covered by peritoneum and, as the bladder fills, it moves smoothly against adjacent structures. Connective tissue over other parts of the bladder attaches it to adjacent structures.
Urethra
The urethra is a tube that carries urine from the urinary bladder to the exterior. In the male, it also shares reproductive functions. Further details of the male urethra are given in
Chapter 15. In females, the urethra is a relatively short tube and most is lined with a stratified squamous epithelium.
Clinical note
Clinical note
Bladder cancer Carcinogens (cancer-causing substances) are widely used in chemical industries, particularly in those producing dyes and rubber. Without due care, people working in these industries are at increased risk of developing bladder cancer. The carcinogens and/or their metabolites are excreted via the kidneys, and the bladder epithelium, exposed to high levels of the carcinogens, undergoes cancerous changes.
The urinary system
■ This consists of paired kidneys and the urinary tract (paired ureters, urinary bladder and urethra).
Kidneys
■ The kidneys comprise cortex and medulla.
■ The functional unit is the nephron, comprising the renal corpuscle and uriniferous tubule.
■ Renal corpuscles are present in the cortex:
■ an afferent arteriole supplies glomerular capillaries within each corpuscle and an efferent arteriole drains the glomerular capillaries
■ the glomerular capillaries are surrounded by a double-walled cup lined by epithelial cells
■ the inner layer of epithelial cells (podocytes) is closely applied to the endothelial cells lining the capillaries and they share basement membranes:
■ filtration of blood occurs across the endothelial cells, basement membranes and podocytes
■ the filtrate enters the urinary space between the walls of the cup
■ the filtrate drains from the cup into a uriniferous tubule.
■ Each uriniferous tubule comprises an initial, convoluted proximal tubule (in the cortex), a straight looped portion (loop of Henle) extending from the cortex and into the medulla and a distal convoluted tubule (in the cortex).
■ The low columnar epithelial cells lining proximal tubules absorb water, proteins and many of the filtered sodium and chloride ions.
■ Some of the cuboidal epithelial cells lining the loops of Henle actively pump sodium ions into the surrounding connective tissue, which becomes hypertonic.
■ Cuboidal epithelial cells lining the distal tubules resorb sodium ions.
■ Some resorbed ions return to blood capillaries in the cortex and medulla.
■ Uriniferous tubules drain into collecting ducts.
■ Aldosterone and antidiuretic hormone control the amount of water and ions resorbed and the tonicity of urine, thus aiding homeostasis.
Ureters
■ Each transports urine from a renal pelvis to the urinary bladder.
■ They are lined by a mucosa consisting of a transitional (multilayered) epithelium and connective tissue:
■ the transitional epithelium ensures hypertonic urine does not equilibrate with isotonic tissue fluid and blood
■ They have smooth muscle layers which contract and pass the urine into the bladder.
■ They are attached by an outer layer of connective tissue to adjacent structures.
Urinary bladder
■ The urinary bladder has a mucosa and muscularis similar to the ureter but is able to store a large volume of urine. As urine is stored the mucosa unfolds, the number of layers of epithelial cells is reduced and the epithelial cells flatten.
■ After infancy, parasympathetic nerves (under conscious control) stimulate contraction of the muscularis and this empties the bladder into the urethra at micturition.
Urethra
■ It is short in females and lined by a stratified squamous epithelium.
■ It is longer in males and shared with the reproductive system.
[/level-membership-for-basic-science-category][not-level-membership-for-basic-science-category]The urinary system consists of paired kidneys and the urinary tract, which comprises paired ureters, a urinary bladder and a urethra (Fig. 13.1). Urine is produced by the kidneys and passes along the ureters to the urinary bladder, where it is stored until it is voided via the urethra. The route taken by urine along the urethra to the exterior is an independent closed system in females, but is shared with the reproductive system in males (Chapter 15).
The urinary system is essential in maintaining the homeostasis of the body. It does this by regulating the water and mineral salts in, and the acid–base balance of, blood. It is particularly important in excreting toxic molecules containing nitrogen (e.g. urea and creatinine) produced by the breakdown of endogenous proteins. The urinary system also ensures that useful molecules in blood, e.g. proteins and carbohydrates, are not lost during the formation of urine. The toxic molecules and excess ions, dissolved in water, leave the kidneys as urine, which passes along the urinary tract before being voided at micturition. In addition, the kidneys produce and secrete into blood two molecules, renin and erythropoietin. The former is important in regulating blood pressure and the latter in stimulating the formation of red blood cells.
Kidneys
The gross structure of a kidney is best described as seen in a longitudinal hemi-section (
Fig. 13.2). Facing towards the mid-line of the body, each kidney has an indentation forming a hilum. At the hilum blood enters and leaves each kidney in, respectively, a renal artery and a renal vein, and urine drains into a ureter.
The gross appearance of the kidney displays an outer and an inner region, the cortex and the medulla. The cortex has a granular appearance due to the presence of spherical structures about 200
μm in diameter known as renal corpuscles. These corpuscles filter blood in the initial stage in the formation of urine. The cortex also contains convoluted tubules involved in forming urine. Renal corpuscles are not present in the medulla and the medulla appears smooth or may show striations. Straight and arching tubules are present in the medulla and they also are involved in forming urine. Renal corpuscles and tubules may be distinguished in histological sections even at low magnification (Fig. 13.3). In humans, the medulla projects centrally as several pyramids and the cortex extends as columns between the pyramids (Fig. 13.2). The apices of the pyramids project as renal papillae into urine-filled spaces (minor calyces). Urine drains from tubules in the medulla into the calyces. Each minor calyx drains urine into major calyces that together form the renal pelvis. The relatively large, fluid-filled space of each renal pelvis drains into a ureter (Fig. 13.2). Fat cells surround the renal pelvis, the ureter and vessels at the hilum of the kidney and pack a space known as the renal sinus. A dense layer of fat surrounds each kidney.The blood supply to kidneys ensures that the whole volume of blood in the body passes through the kidneys every 5 minutes or so. The arrangement of blood vessels is based on supplying (and draining) each medullary pyramid and its associated cortical tissues separately, a unit described as a renal lobe. (The human kidney is multilobar, but uni-lobar kidneys occur in many species.) Each renal artery branches and forms interlobar arteries. In turn, interlobar arteries branch further and then form arcuate arteries which arch between the cortex and medulla (
Fig. 13.4). Branches from arcuate arteries eventually supply each renal corpuscle with an afferent arteriole. This arrangement is essential in filtering the blood (see below) as well as supplying oxygen and nutrients.
Each afferent arteriole divides within a renal corpuscle into capillaries, known as glomerular capillaries (Fig. 13.5), and these drain from the corpuscle into an efferent arteriole (
Fig. 13.6). This arrangement of capillaries draining to an arteriole is unique to the kidney and results in a relatively high blood pressure in glomerular capillaries (
Fig. 13.5) which aids the filtration of blood. Efferent arterioles divide and form a second network of capillaries. Some of these capillaries are straight vessels, vasa recta, which lie between straight tubules in the kidney whereas others form an extensive meshwork surrounding convoluted tubules in the cortex (
Fig. 13.6). The second set of capillaries supplies oxygen and nutrients to the cells of the kidney and is also involved in modifying the filtrate and forming urine. Veins drain capillaries (other than glomerular capillaries) in the kidney and the routes they take closely follow the arterial pattern.
Nephrons and collecting ducts
A nephron is the functional unit of the kidney (
Fig. 13.6) and there are about a million in each kidney in humans. A nephron consists of a renal corpuscle, which filters blood, and the uriniferous tubule attached to it which drains and modifies the filtrate. Eventually, the modified filtrate becomes urine and drains, via collecting ducts, into the renal pelvis.
Renal corpuscles
Each renal corpuscle consists of glomerular capillaries (a renal glomerulus) (Fig. 13.5) within a Bowman’s capsule. The capsule is shaped like a hollow, double-walled cup (
Fig. 13.6) and it is lined by epithelial cells. The outer parietal layer of Bowman’s capsule is lined by squamous epithelial cells (
Fig. 13.7) and it is continuous with the inner visceral layer of epithelial cells. The cells of the inner layer, known as podocytes, have numerous cytoplasmic processes which abut the capillaries of the glomerulus. The endothelial cells lining the capillaries and the podocytes (and their fused basement membranes) form the filtration barrier between the blood and the filtrate. The space between the inner and outer layers of the capsule receives the filtrate and is known as the urinary space (
Fig. 13.7) even though the filtrate is not yet urine.
Each Bowman’s capsule has two poles, and it is extremely rare to see all the features of each pole in any one (histological) section of a corpuscle.
■
Vascular pole (
Figs 13.6 and
13.7). At this pole an afferent arteriole enters each renal corpuscle, branches and forms glomerular capillaries. These capillaries rejoin and drain into an efferent arteriole (not a venule), which leaves the corpuscle at the vascular pole.
■
Tubular pole (
Fig. 13.6,
Fig. 13.7 and
Fig. 13.8). At this pole the filtrate drains from the urinary space of the renal corpuscle. It enters the proximal part of the uriniferous tubule, which extends from the parietal layer of Bowman’s capsule at this pole.
Filtration
Blood in glomerular capillaries is filtered through the endothelium lining the capillaries. It then traverses the fused basement membranes of the endothelium and of the adjacent podocytes (the visceral epithelial cells of Bowman’s capsule) (
Fig. 13.9).
Filtration is affected by:
■ the pressure of blood in the afferent arteriole
Buy Membership for Basic Science Category to continue reading.
Learn more here
[/not-level-membership-for-basic-science-category]
Histology An Illustrated Colour Text