Chapter 12
The Pons and Cerebellum
Vasculature of the Pons and Cerebellum
Summary of Descending Pathways
Internal Vasculature of the Pons
Vascular Syndromes of the Pons
The metencephalon consists of the pons and cerebellum. The pons is the middle segment of the brainstem, the caudal part being the medulla and the rostral portion being the midbrain. Although comprising only about 1.3% of the brain by weight, the pons has many important functions. The motor and sensory nuclei and the exit points of cranial nerves V to VIII are associated with the pons. The cerebellum is not part of the brainstem but rather is considered a suprasegmental structure because it is located superior to the brainstem. The cerebellum is comparatively large, comprising about 10.5% of the total brain weight. Functionally, it is part of the motor system. The blood supply to the pons and cerebellum arises from branches of the basilar and cerebellar arteries.
DEVELOPMENT
The pons and cerebellum are considered together in this chapter because they arise from the same region of the developing neural tube. The metencephalon extends from the pontine flexure to the mesencephalic isthmus (Fig. 12-1). At this level, the cavity of the neural tube is enlarged, forming the parts of the fourth ventricle associated with the pons and cerebellum.
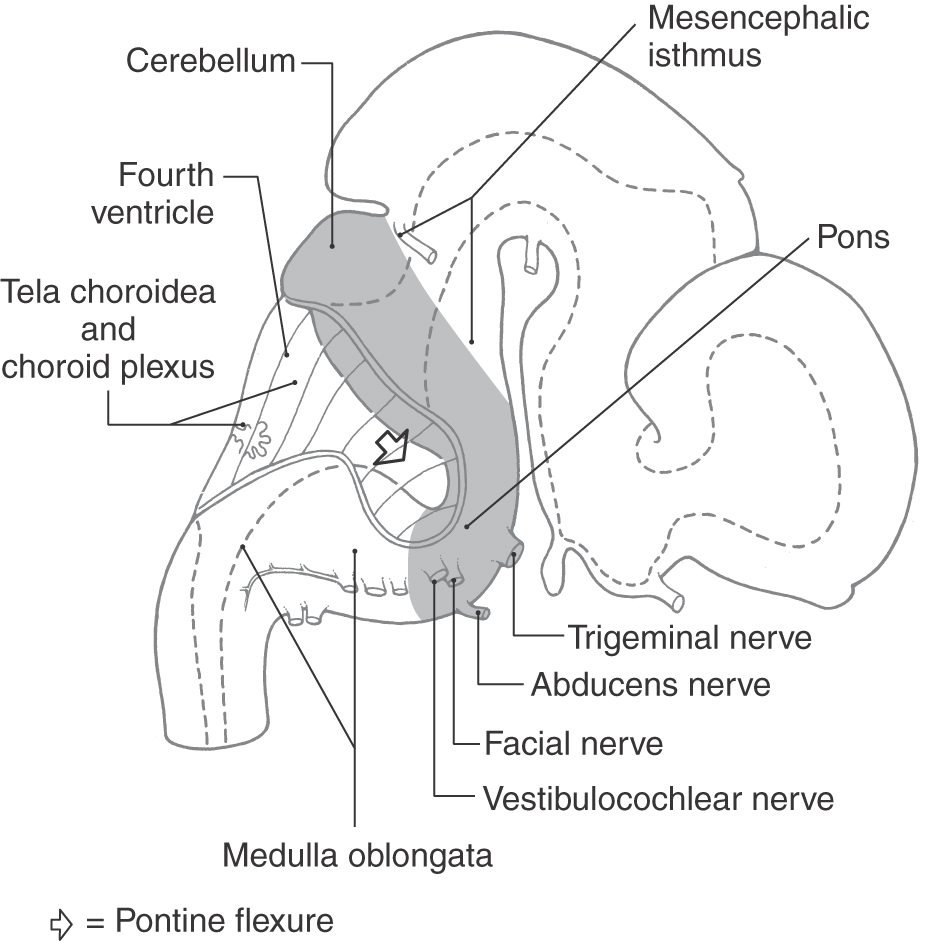
Figure 12-1. Lateral view of the brain at about 7 weeks of gestational age. The pons and cerebellum are highlighted.
Basal and Alar Plates
The basal and alar plates of the brainstem extend from the medulla rostrally into the developing pons. The cranial nerve motor nuclei found in the pons (trigeminal, abducens, facial, and superior salivatory) originate from the basal plate and are located medial to the sulcus limitans (Fig. 12-2A, B). The functional components of these motor neurons include somatic efferent (SE) (trigeminal, facial, and abducens) and visceral efferent (VE) (superior salivatory).
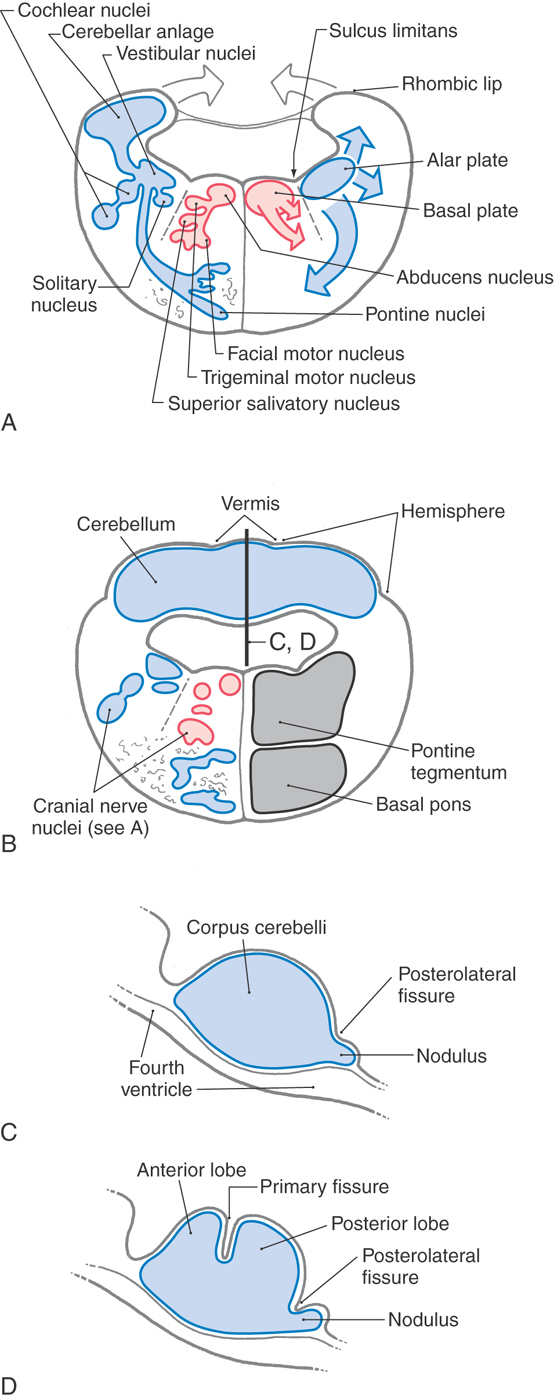
The cranial nerve sensory nuclei located in the pons include portions of the trigeminal and vestibulocochlear nuclei and the rostral tip of the solitary nucleus. These nuclei originate from the alar plate and are found lateral to the sulcus limitans (Fig. 12-2A, B). Their functional components are somatic afferent (SA) for the trigeminal nucleus and vestibular and cochlear nuclei and visceral afferent (VA) for the solitary nucleus. This recognizes the fact that taste may be generally regarded as a special sense. The portion of the posterior pons that contains these motor and sensory nuclei as well as the reticular formation and several ascending and descending tracts is the pontine tegmentum (Fig. 12-2B).
The anterior area of the developing pons is invaded by large numbers of descending cortical fibers. Although some will terminate here, others pass through to more caudal targets. Immature alar plate neurons also migrate into this anterior pontine region to form the basilar pontine nuclei. These nuclei, their axons, and the descending fibers passing to and through this area collectively form the basilar pons (Fig. 12-2B).
Cerebellum
The cerebellum develops from the rhombic lips of the pontine alar plates. These lips expand posteromedially toward each other until they meet at the midline and fuse to form the cerebellar plate (Fig. 12-2A, B), which is the rudiment of the cerebellum. As development progresses, the cerebellum is divided by transverse fissures into lobes and lobules. The first of these fissures to appear is the posterolateral fissure, which separates the flocculonodular lobe caudally from the corpus cerebelli rostrally. The primary fissure, the second to appear, divides the corpus cerebelli into the anterior and posterior lobes (Fig. 12-2B-D). Internal changes, such as development of the cerebellar cortex and nuclei, take place concurrently with these external events.
EXTERNAL FEATURES
Basilar Pons
The portion of the brainstem lying between the midbrain rostrally and the medulla caudally is the pons (pons, Latin for “bridge”). Anteriorly and laterally (Fig. 12-3), the pons consists of a massive bundle of transversely oriented fibers that enter the cerebellum as the middle cerebellar peduncle (brachium pontis). The exit of the trigeminal nerve marks the transition from the basilar pons, which is anterior to the trigeminal root, to the middle cerebellar peduncle, which lies posterior to the exit of the trigeminal nerve (Figs. 12-3 and 12-4). Rostrally, the large axonal bundles forming the crus cerebri of the midbrain extend into the basilar pons. Caudally, some of these axons emerge to form the pyramids of the medulla (Fig. 12-4A).

Figure 12-3. Anterior (ventral) view of the brainstem with emphasis on the pons.
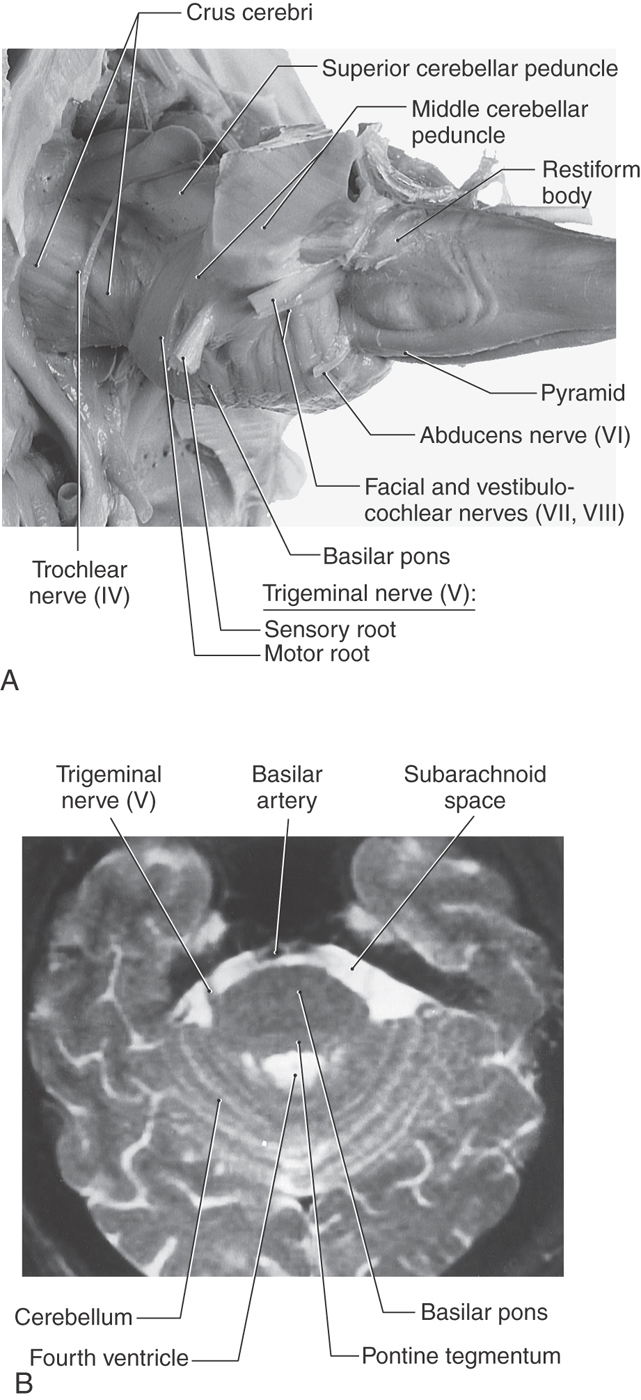
The cranial nerves that emerge from the pons are the trigeminal (V), abducens (VI), facial (VII), and vestibulocochlear (VIII). The trigeminal nerve exits laterally and is composed of a large sensory root (the portio major) and a small motor root (the portio minor) (Fig. 12-4A). The portion of the trigeminal nerve that traverses the subarachnoid space between the pons and the trigeminal ganglion forms a landmark that is visible on magnetic resonance imaging at this level (Fig. 12-4B). The abducens, facial, and vestibulocochlear nerves emerge in medial to lateral sequence along the pons-medulla junction (Fig. 12-3). Although cranial nerve VII is commonly called the facial nerve, it is actually composed of two roots, the facial nerve (SE fibers) and the intermediate nerve (VA, VE, and SA fibers). The vestibulocochlear nerve (SA fibers) emerges posterolaterally and, with the facial and intermediate nerves and labyrinthine artery, occupies the internal acoustic meatus.
Rhomboid Fossa of the Pons
The rhomboid fossa forms the floor of the fourth ventricle. Its caudal portion is located in the medulla, and its larger, more rostral area is in the pons. The posterior surface of the pontine tegmentum, which forms the floor of the fourth ventricle, is visible only when the cerebellum is detached from the brainstem (Fig. 12-5). This part of the ventricular floor is characterized by the facial colliculus located between the median fissure and the superior fovea of the sulcus limitans and by the vestibular area located lateral to the sulcus limitans. The facial colliculus is formed by the underlying abducens nucleus and internal genu of the facial nerve, and the vestibular area marks the location of the vestibular nuclei. The brachium pontis and the brachium conjunctivum form the lateral walls of the fourth ventricle in the pons; the roof is formed by the anterior medullary velum, by a small part of the cerebellum, and by a portion of the tela choroidea (Fig. 12-6).
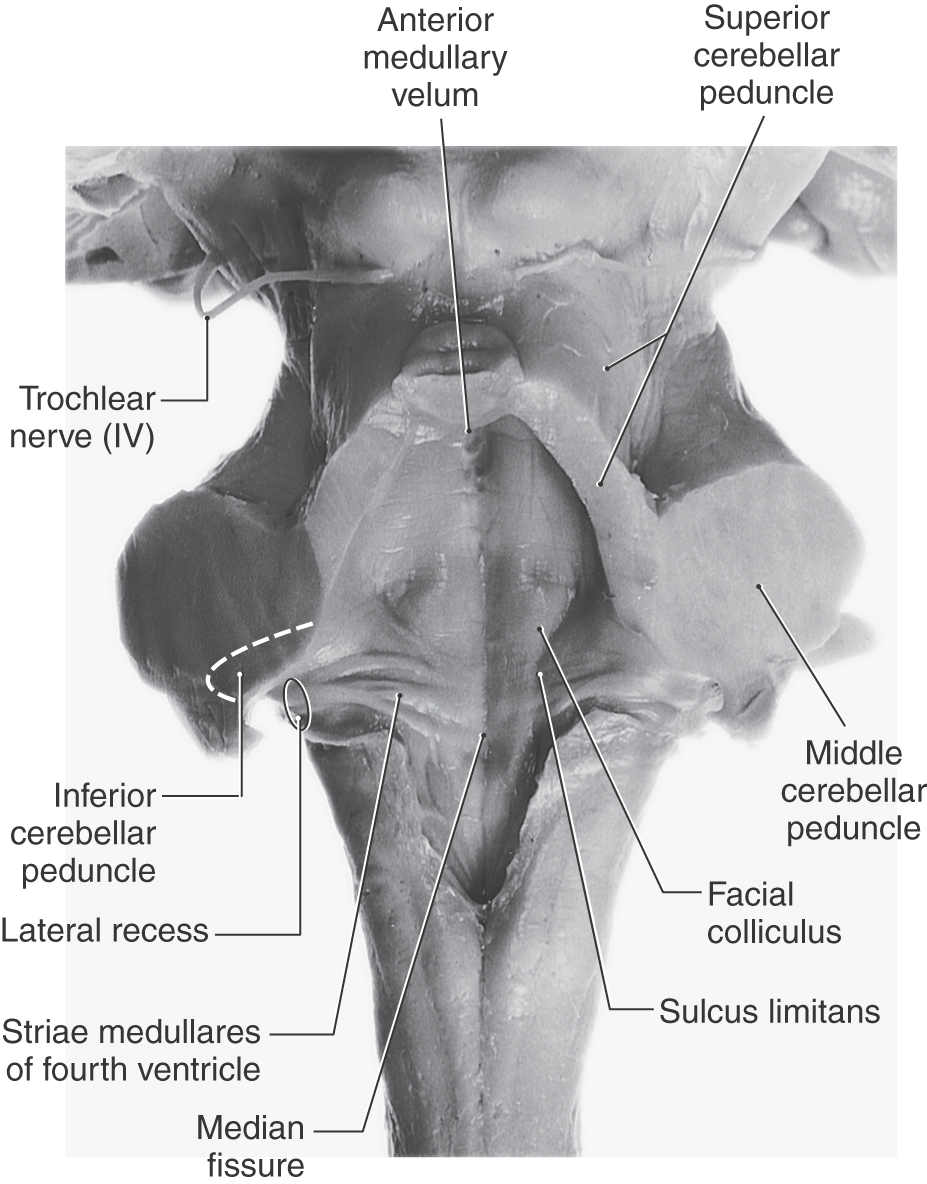
Figure 12-5. Pontine part of the fourth ventricle and rhomboid fossa. Also see Figure 10-4 for further details of the rhomboid fossa.
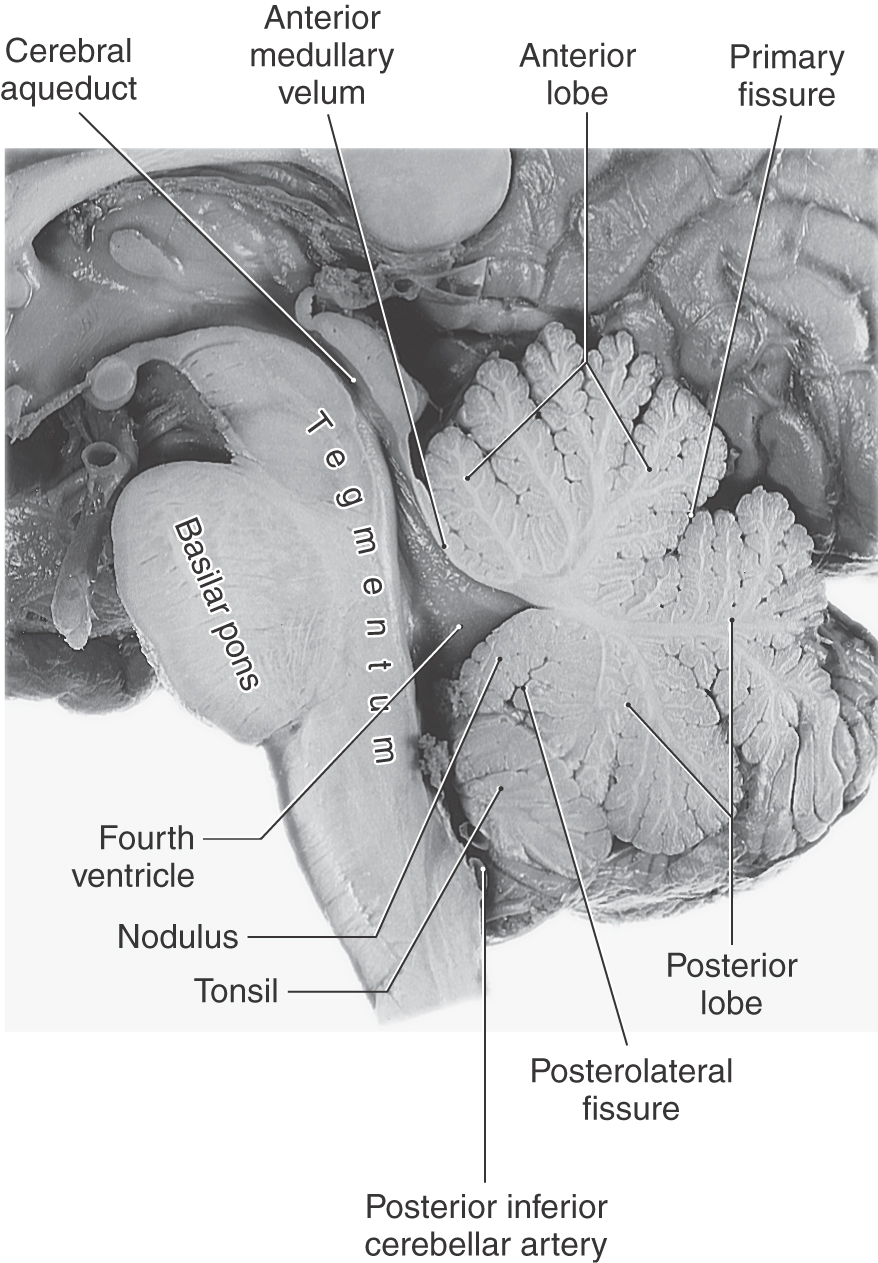
Figure 12-6. Sagittal view of the brainstem (with emphasis on the pons) and cerebellum.
Cerebellum
The cerebellum is located posterior to the brainstem and fills much of the posterior fossa. It is attached to the brainstem by three pairs of cerebellar peduncles (superior, middle, and inferior). In sagittal section, the human cerebellum appears wedge shaped (Fig. 12-6), with its superior surface apposed to the tentorium cerebelli and its inferior surface curving toward the foramen magnum.
The cerebellum consists of anterior, posterior, and flocculonodular lobes; each lobe, in turn, is composed of lobules (Figs. 12-6 and 12-7). Lobes and lobules are separated from each other by fissures. Lobules are made up of yet smaller folds of cerebellar cortex called folia (singular, folium). Cerebellar folia, lobules, and lobes can often be followed across the midline from one side of the cerebellum to the other.
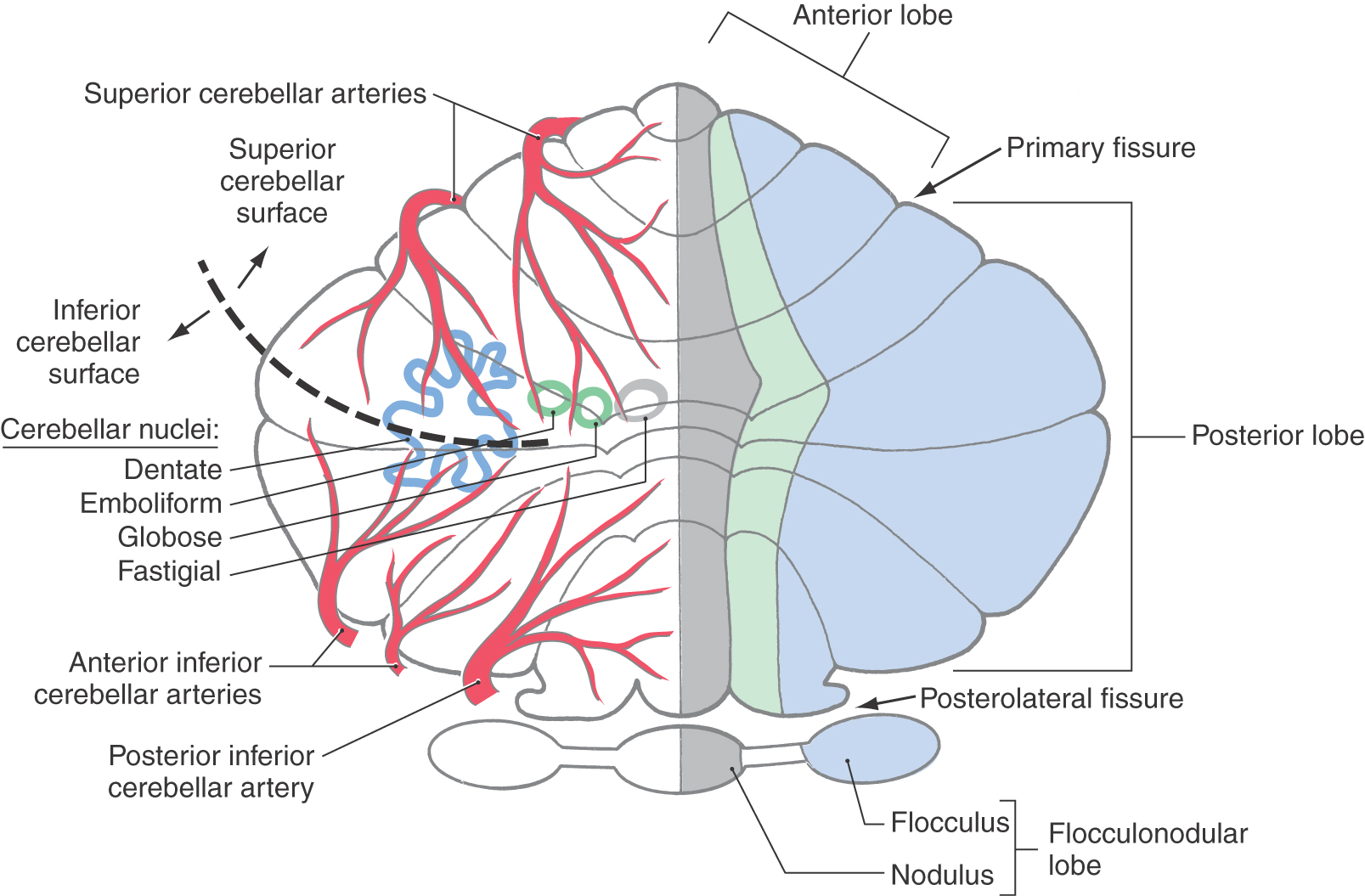
Each cerebellar lobe (and lobule) is also divided into rostrocaudally oriented regions of cortex commonly called the vermis (medial), intermediate (paravermis), and hemisphere (lateral) zones (Fig. 12-7). The vermal zone is approximately 1.0 cm across at its widest point. The hemisphere is expansive in the human cerebellum and is separated from the vermis by the intermediate zone.
Four cerebellar nuclei are located in the white matter core of each hemisphere. From medial to lateral, they are the fastigial, globose, emboliform, and dentate nuclei (Fig. 12-7). These cells receive input from branches of cerebellar afferent fibers and from Purkinje cells located in the cerebellar cortex. In turn, axons of cerebellar nuclear cells provide the main output signals of the cerebellum. The structure, function, and connections of the cerebellar cortex and nuclei are considered in greater detail in Chapter 27.
Vasculature of the Pons and Cerebellum
The basilar artery and its branches serve basilar and tegmental areas of the pons. The internal distribution of the basilar artery is discussed later in this chapter. The superior cerebellar artery, through its medial and lateral branches, distributes to the superior surface of the cerebellum and most of the cerebellar nuclei; the inferior surface of the cerebellum is served by anterior and posterior inferior cerebellar arteries (Fig. 12-7).
INTERNAL ANATOMY OF THE PONS
Summary of Ascending Pathways
The major ascending pathways seen in the medulla continue into the pons (Fig. 12-8). These include the medial lemniscus, anterolateral system, anterior trigeminothalamic fibers, and anterior spinocerebellar tract. Although most of these fibers continue through the pons, some anterolateral system fibers terminate in the pontine reticular formation (as spinoreticular fibers), and anterior spinocerebellar axons enter the cerebellum on the surface of the superior cerebellar peduncle. The restiform body, a prominent structure in the rostral medulla, sweeps posteriorly into the cerebellum in the caudal pons as the largest part of the inferior cerebellar peduncle.
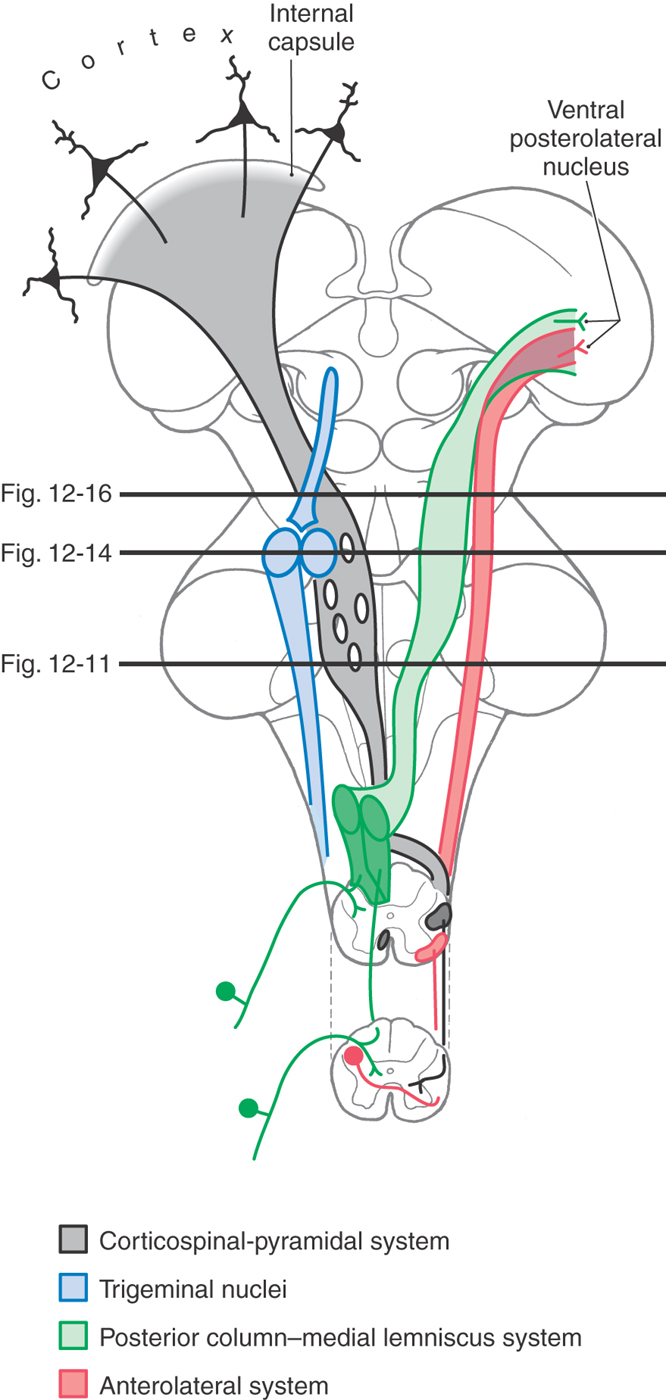
Figure 12-8. Diagrammatic representation of the brain showing the location and trajectory of three important pathways and the trigeminal nuclei. The color coding for each is continued in Figures 12-11, 12-14, and 12-16.
Summary of Descending Pathways
The most prominent groups of descending fibers arise from cells located in the midbrain or forebrain and therefore traverse the pons (Fig. 12-8). These include the corticospinal fibers, the central tegmental and rubrospinal tracts, and the tectobulbospinal system. The medial longitudinal fasciculus occupies a characteristic position near the midline in the floor of the fourth ventricle. At the pons-medullary junction, this bundle contains mainly descending fibers; in the rostral pons, it is made up primarily of ascending fibers.
Caudal Pontine Level
The pons is divided into a posterior part, the tegmentum, and an anterior region, the basilar pons. This section and the following two sections describe the anatomy of the pons at three levels: caudal, midpontine, and rostral pontine levels. Each level is described beginning with the tegmentum and proceeding to the basilar pons.
At caudal pontine levels, the facial colliculus is formed by the underlying abducens nucleus and fibers comprising the internal genu of the facial nerve (Figs. 12-10 and 12-12). Axons from the SE cells of the abducens nucleus course anteriorly through the tegmentum, pass adjacent to the corticospinal fibers in the basilar pons, and exit the brainstem at the pons-medullary junction as the abducens nerve (Figs. 12-10 and 12-12). The internal genu of cranial nerve VII is composed of the axons of SE cells from the facial nucleus. These axons loop around the abducens nucleus from caudal to rostral, as the internal genu, then course anterolaterally to exit the brainstem (Fig. 12-12). Anterolateral to the abducens nucleus, these SE fibers are surrounded by cells of the superior salivatory nucleus, the axons of which exit the brainstem as the VE component of the intermediate nerve (Fig. 12-12).
Medial to the abducens nucleus is the medial longitudinal fasciculus and the tectobulbospinal system (Fig. 12-11). As in the medulla, these bundles are internal to the ventricular space and adjacent to the midline.
The posterolateral tegmentum contains the vestibular nuclei and the solitary tract and nucleus (Figs. 12-9 and 12-11). The lateral, medial, and inferior vestibular nuclei are present at this level, whereas the superior vestibular nucleus becomes prominent more rostrally. The small bundles of fibers coursing between the vestibular nuclei and the cerebellum in the wall of the fourth ventricle form the juxtarestiform body (Figs. 12-10 and 12-11). This structure is composed of vestibulocerebellar and cerebellovestibular fibers and along with the laterally adjacent restiform body constitutes the inferior cerebellar peduncle. The rostral portions of the solitary tract and nucleus are located anterior to the vestibular nuclei and consist of a core of primary sensory fibers (tract) surrounded by cell bodies (nucleus). This part of the solitary complex receives mainly taste input (VA fibers) and is sometimes called the gustatory nucleus.
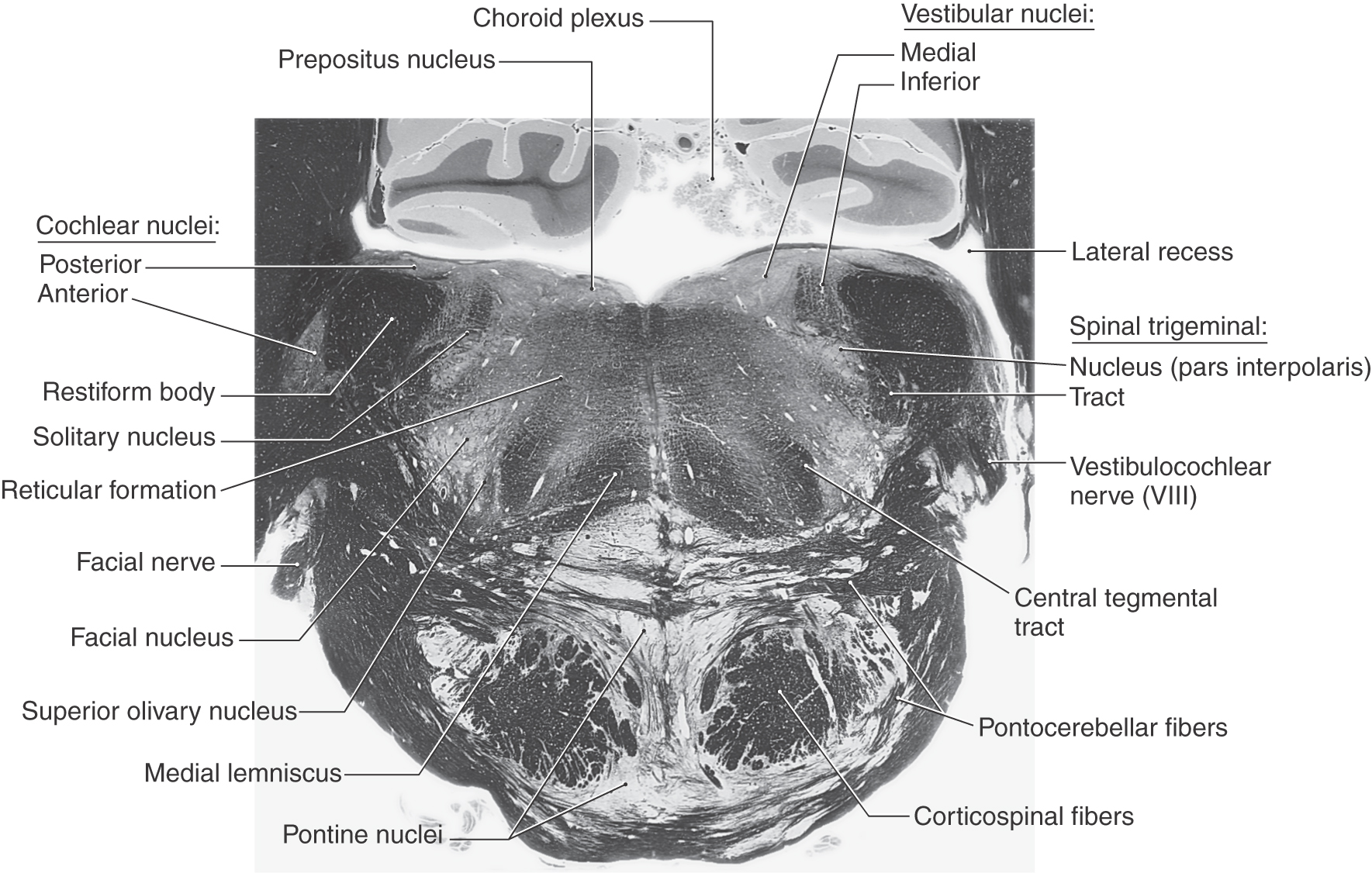
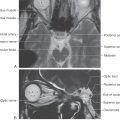
 Figure 12-9. A fiber (myelin)–stained cross section at the level of the facial motor nucleus (caudal pons).
Figure 12-9. A fiber (myelin)–stained cross section at the level of the facial motor nucleus (caudal pons).
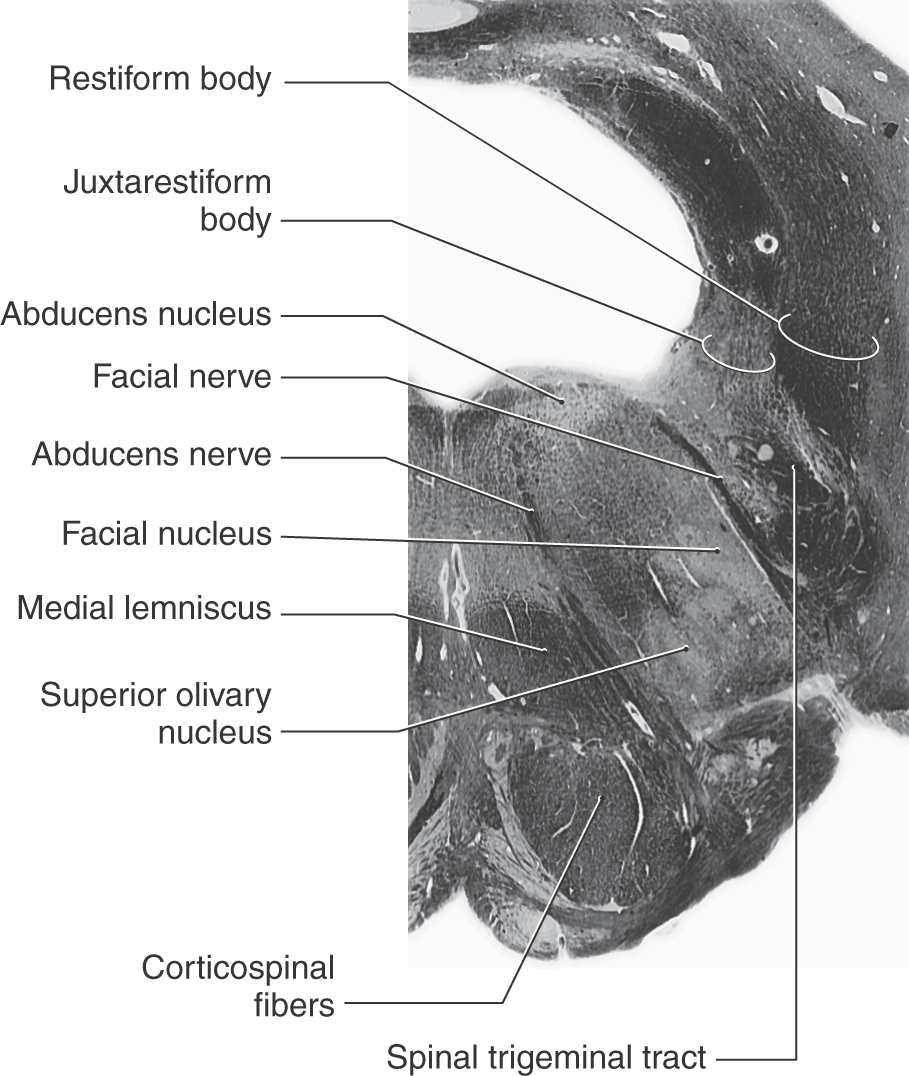
 Figure 12-10. A fiber (myelin)–stained cross section of the pons at the level of the facial colliculus. Compare with Figure 12-11.
Figure 12-10. A fiber (myelin)–stained cross section of the pons at the level of the facial colliculus. Compare with Figure 12-11.
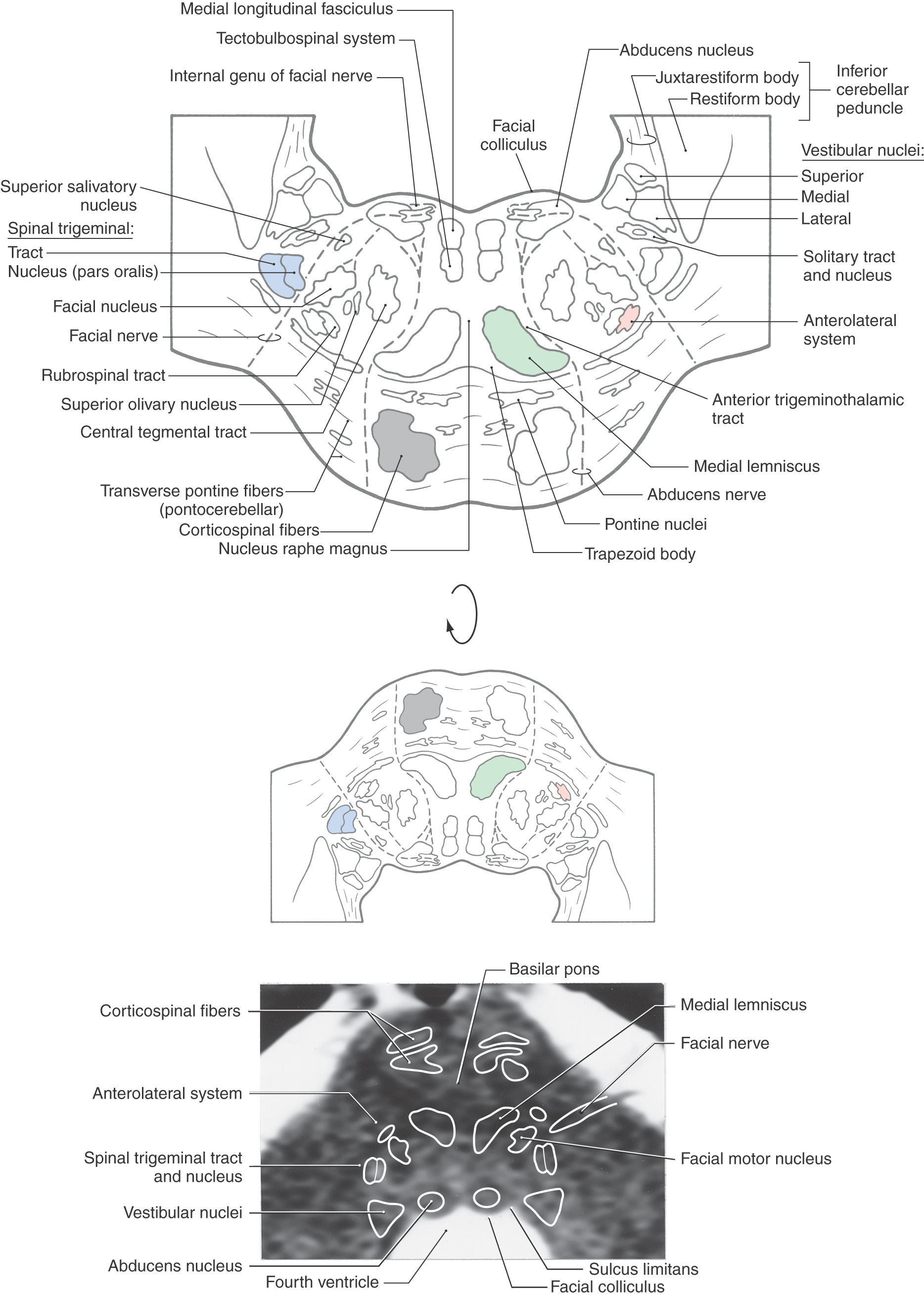
Figure 12-11. Cross section through the caudal pons at the level of the facial colliculus. Correlate with Figure 12-8. The anatomic orientation is flipped to illustrate internal structures in a clinical orientation; the clinically important tracts and nuclei are shown on a T2-weighted magnetic resonance image at a comparable level of the facial colliculus in the caudal pons.
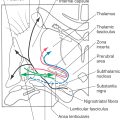
The central portion of the pontine tegmentum at caudal levels contains, from medial to lateral, the central tegmental tract, the superior olivary nucleus, the facial motor nucleus, and the spinal trigeminal tract and nucleus (Figs. 12-9, 12-11, and 12-12). A major part of the central tegmental tract includes fibers coursing from the red nucleus of the midbrain to the inferior olive of the medulla (rubroolivary fibers). Cells of the superior olive receive input from the anterior cochlear nucleus and send their axons into the lateral lemniscus on both sides. The route followed by motor facial fibers is shown in Figure 12-12; these axons innervate the ipsilateral muscles of facial expression. The spinal trigeminal tract is composed of sensory (SA) fibers from the ipsilateral half of the face, oral cavity, and much of the scalp. Although most of this input is via the trigeminal nerve (hence the name of the tract), cranial nerves VII, IX, and X also make modest contributions to the spinal trigeminal tract and nucleus. Axons of the spinal trigeminal tract synapse in the spinal trigeminal nucleus, the cells of which project to the contralateral thalamus as anterior trigeminothalamic fibers.
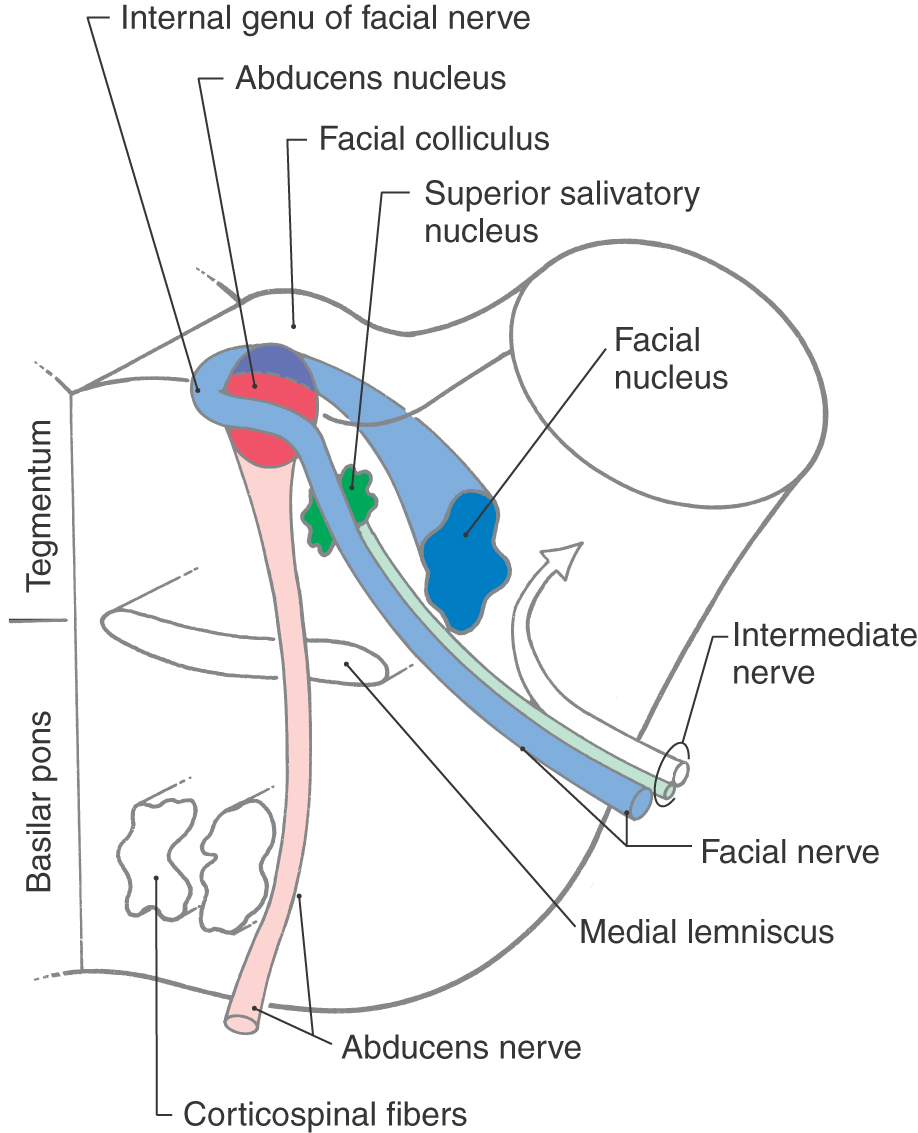
 Figure 12-12.
Figure 12-12. The anterolateral system, rubrospinal tract, and trapezoid body are located in the anterolateral tegmentum (Fig. 12-11). Although pain and temperature signals from the contralateral side of the body are conveyed by anterolateral system fibers, some of these axons end in the pontine reticular formation as spinoreticular fibers. The trapezoid body is composed of decussating axons from the cochlear nuclei. After crossing, these fibers ascend to form the lateral lemniscus and convey auditory signals to the midbrain.
The medial lemniscus is oriented vertically in the medulla, but in the caudal pons it begins to shift to a horizontal position (Fig. 12-13; see also Fig. 12-9). At this level, the anterior part of the medial lemniscus (lumbosacral representation) shifts somewhat laterally, and its posterior portion (cervicothoracic representation) assumes a more medial location. The medial lemniscus forms the border between the tegmentum and basilar pons.
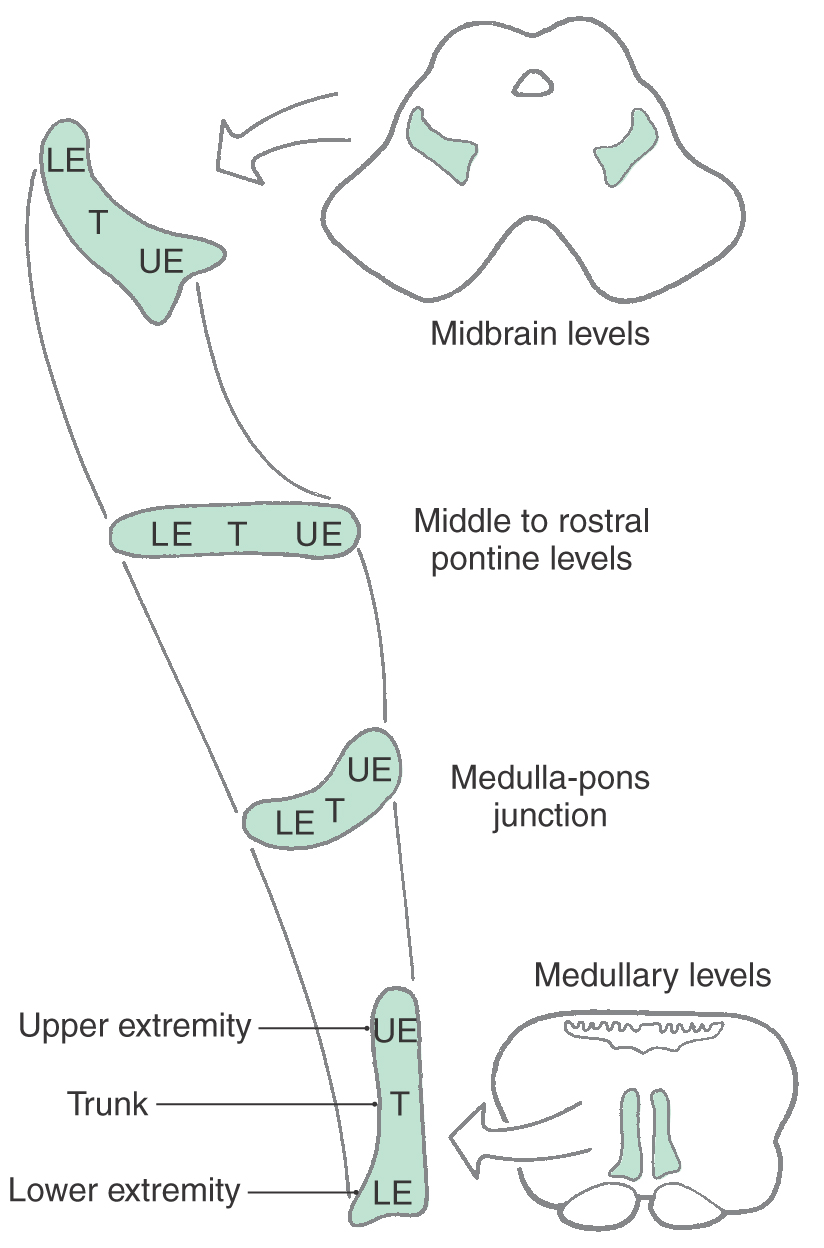
 Figure 12-13. The orientation of the medial lemniscus at all brainstem levels.
Figure 12-13. The orientation of the medial lemniscus at all brainstem levels.
The anterior pons contains the basilar pontine nuclei, longitudinally running corticospinal and corticopontine fibers, and transversely oriented pontocerebellar fibers (Figs. 12-9 and 12-11). On each side of the midline, the corticospinal fibers located in the pyramid of the medulla are, in the basilar pons, completely surrounded by the basilar pontine nuclei. These pontine cells receive input from diverse regions of the neuraxis. In turn, most of their axons cross the midline and enter the cerebellum via the middle cerebellar peduncle (brachium pontis) as pontocerebellar fibers.
Midpontine Level
Prominent features of the tegmentum at this level include the principal (or chief) sensory trigeminal nucleus, the trigeminal motor nucleus, and the mesencephalic tract and nucleus (Figs. 12-14 and 12-15). The principal sensory and motor trigeminal nuclei are located in the lateral tegmentum, and the mesencephalic tract and nucleus extend rostrally in the lateral wall of the central gray. Cells of the principal sensory nucleus receive SA input from the ipsilateral trigeminal nerve and project to the thalamus via posterior trigeminothalamic (uncrossed) and anterior trigeminothalamic (crossed) fibers. The SE cells of the trigeminal motor nucleus innervate the masticatory muscles on the ipsilateral side. Last, the unipolar cell bodies of the mesencephalic nucleus and their laterally adjacent processes, the mesencephalic tract, convey SA proprioceptive input to a variety of nuclei, including the trigeminal motor nucleus.
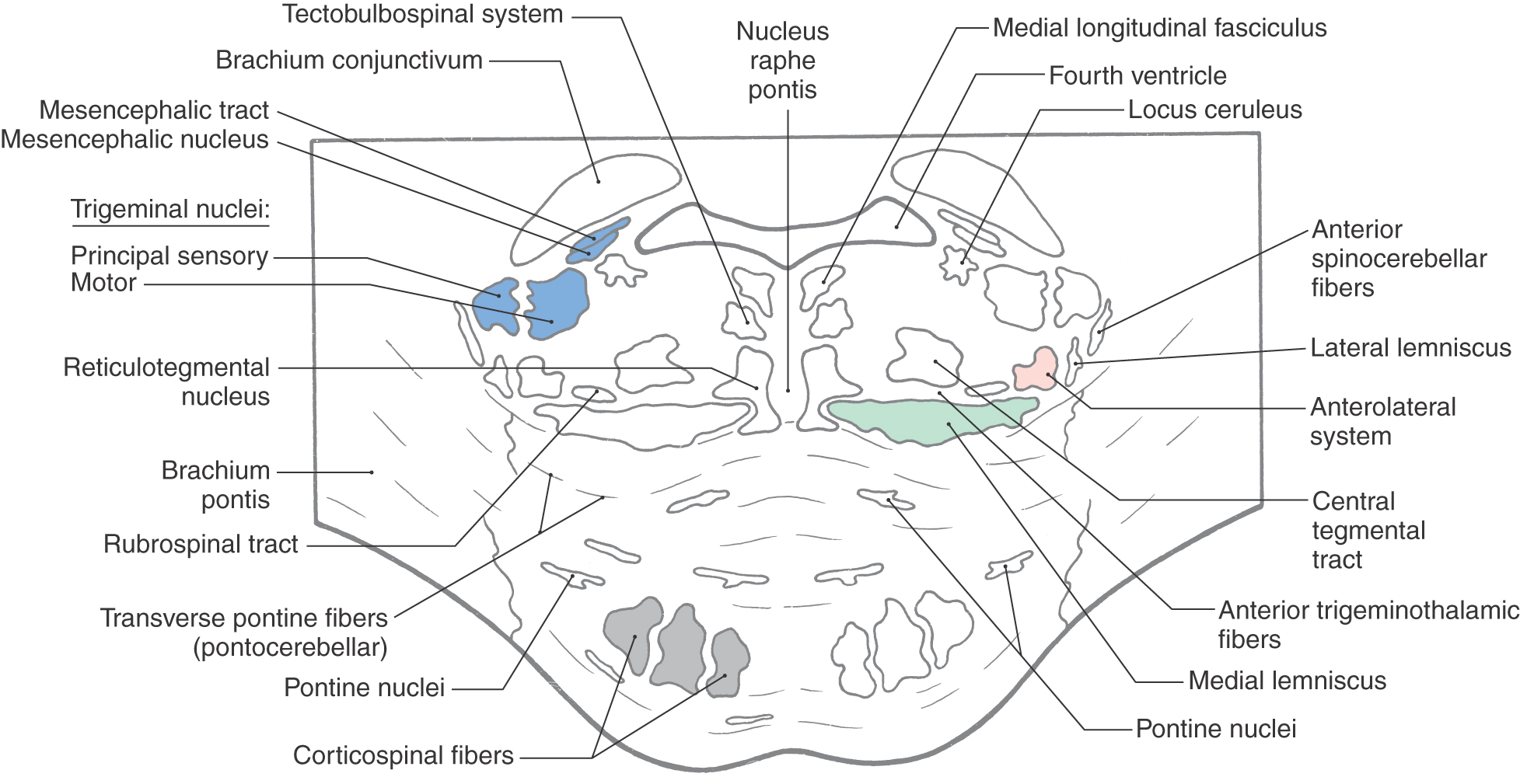
 Figure 12-14. Cross section of the mid pons at the level of the principal sensory and motor trigeminal nuclei. Correlate with Figure 12-8.
Figure 12-14. Cross section of the mid pons at the level of the principal sensory and motor trigeminal nuclei. Correlate with Figure 12-8.
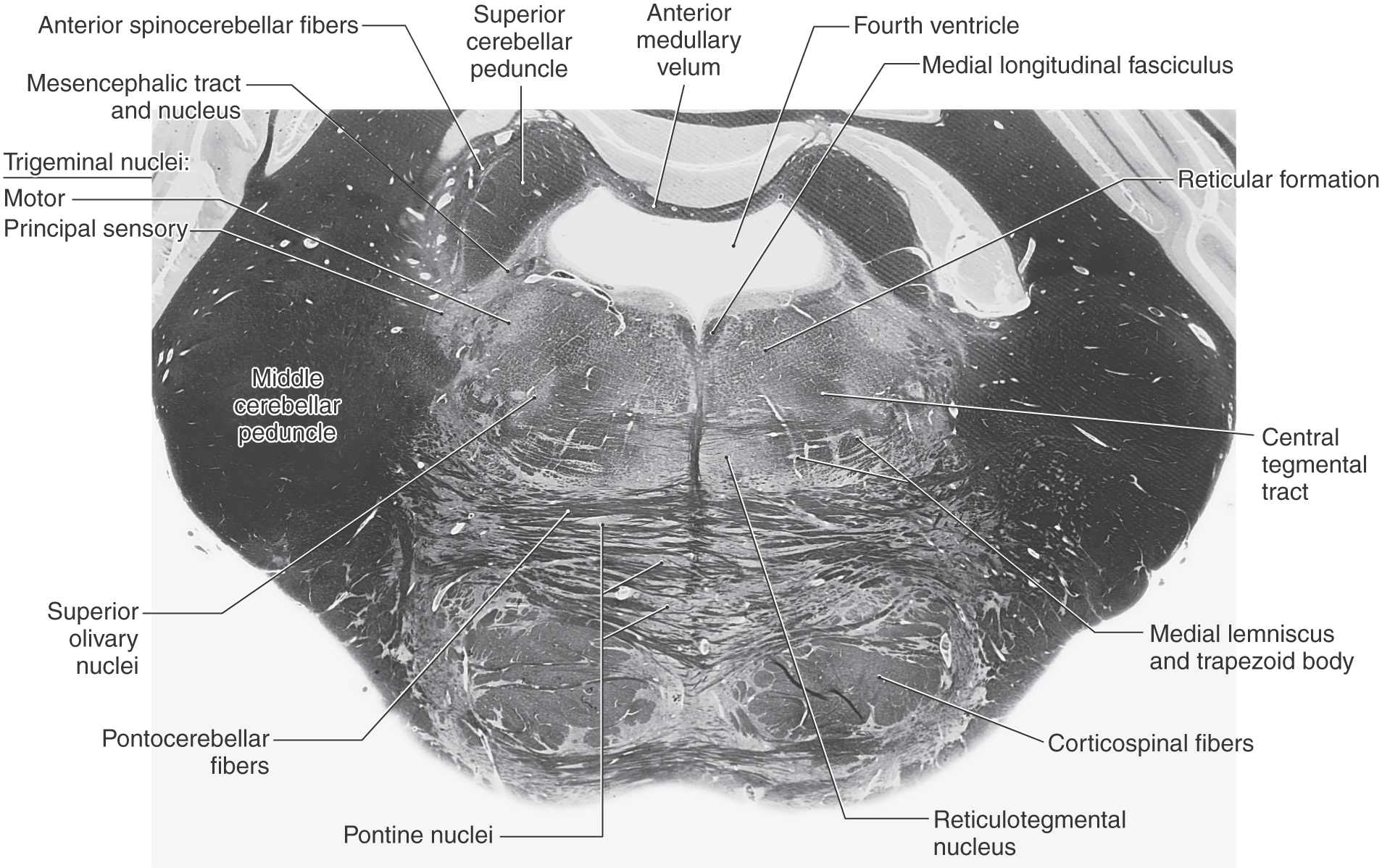
 Figure 12-15. A fiber (myelin)–stained cross section at the level of the principal sensory and motor trigeminal nuclei. Compare with Figure 12-14.
Figure 12-15. A fiber (myelin)–stained cross section at the level of the principal sensory and motor trigeminal nuclei. Compare with Figure 12-14.
The nucleus (locus) ceruleus is located in the lateral floor of the fourth ventricle at this level (Fig. 12-14). Ceruleus neurons contain pigment (hence the alternative name nucleus pigmentosus pontis) and constitute the largest single location of noradrenaline/norepinephrine–containing cells in the central nervous system. Axons arising in the nucleus ceruleus project to widespread targets throughout the cerebral cortex, most of the diencephalon, limbic system, brainstem, cerebellar cortex and nuclei, and spinal cord. This extensive projection arises from a cell group containing only about 15,000 cell bodies, which indicates that these fibers branch profusely. When the activity level of the neurons in this nucleus is low, a state of quiescence is promoted, as in sleep. When there are sudden changes in the patient’s environment, such as waking unexpectedly or if facing a threatening situation, activity of the nucleus ceruleus is high, noradrenaline/norepinephrine is released throughout the nervous system, and the individual is able to attend to the emergency or disruption. During times of normal nonstress and nonsleep activity, these neurons have an intermediate level of activity. However, when the activity of nucleus ceruleus neurons fluctuates outside of levels that are correlated with normal life activities, the patient may experience behavior problems that require medical or psychiatric intervention.
A comparison of Figures 12-11 and 12-14 reveals that most major tracts in the pontine tegmentum (medial longitudinal fasciculus, tectobulbospinal system, medial lemniscus, anterolateral system, and anterior trigeminothalamic fibers) occupy positions comparable to those seen at more caudal levels. Consequently, this section emphasizes only the features that are new at midpontine levels. The medial lemniscus is oriented horizontally at this point (Figs. 12-13 and 12-15), and the rubrospinal fibers have shifted medial to the anterolateral system. Most auditory fibers are now concentrated in the lateral lemniscus, and rostral parts of the superior olivary nucleus appear just lateral to the central tegmental tract. Anterior spinocerebellar fibers migrate posteriorly and enter the cerebellum by coursing over the surface of the superior cerebellar peduncle. The brachium conjunctivum (superior cerebellar peduncle) arises from the cerebellar nuclei, sweeps rostrally, forming the lateral wall of the fourth ventricle (Figs. 12-14 and 12-15), and enters the caudal midbrain tegmentum, where it decussates.
Neurons in the ventral tegmentum close to the midline extend into posterior portions of the basilar pons and constitute the reticulotegmental nucleus (Figs. 12-14 and 12-15). This cell group is continuous with the basilar pontine nuclei, and its axons enter the cerebellum through the contralateral brachium pontis. These cells also share similar features and connections with neurons of the basilar pons.
Rostral Pontine Level
The only cranial nerve structures present in the pontine tegmentum at rostral levels are the mesencephalic nucleus and tract. These structures are located in the lateral aspect of the periaqueductal gray and remain in this position into the midbrain (Fig. 12-16; [see p. 167]; see also Fig. 12-14). Anterior to the mesencephalic tract and nucleus is the locus (nucleus) ceruleus; this noradrenergic cell group also extends into the caudal midbrain.
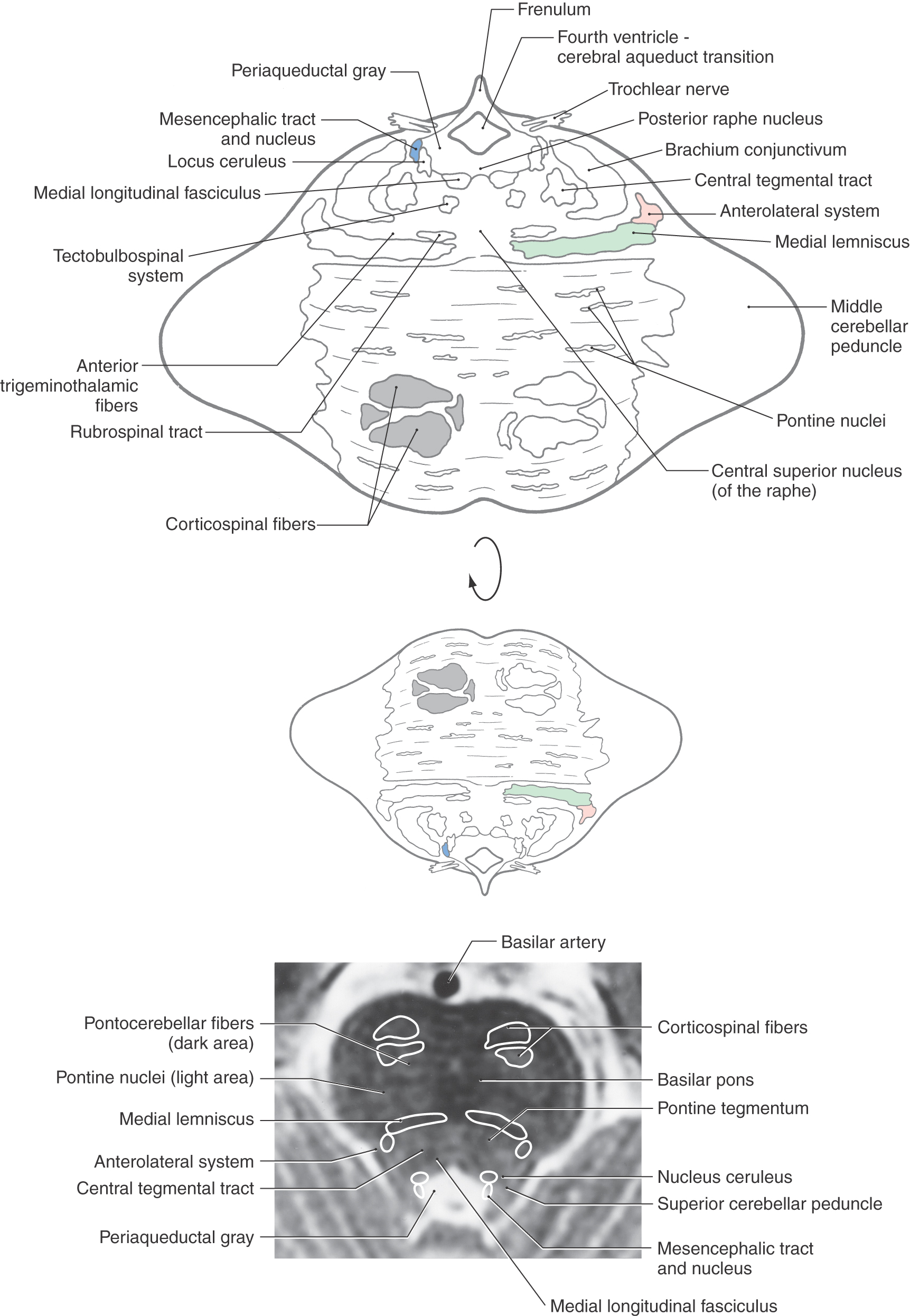
Figure 12-16. Cross section of the rostral pons. Correlate with Figure 12-8. The anatomic orientation is flipped to illustrate internal structures in a clinical orientation; the clinically important tracts and nuclei are shown on a T2-weighted magnetic resonance image at a comparable level of the rostral pons.
The brachium conjunctivum (superior cerebellar peduncle) converges toward its decussation in the caudal midbrain, and most other major tracts in the tegmentum occupy positions comparable to those seen at midpontine levels (compare Fig. 12-14 with Fig. 12-16). The rubrospinal tract is shifted even more medially, and the lateral lemniscus is close to the posterolateral surface of the brainstem at this point. Also, the composition of the basilar pons is essentially the same as that seen at midpontine levels (Figs. 12-14 and 12-16).
Reticular and Raphe Nuclei
Much of the pontine tegmentum is occupied by the reticular formation. This central core is generally divided into a medial area of primarily large neurons (magnocellular region) and a lateral area of mainly small neurons (parvocellular region) (Fig. 12-17; see p. 168). The magnocellular reticular nuclei of the pons are, from caudal to rostral, the gigantocellular reticular nucleus and the caudal and oral pontine reticular nuclei. The parvocellular area nuclei of the pons contain a diffuse lateral reticular formation at caudal and midpontine levels and include the medial and lateral parabrachial nuclei at rostral levels. The latter cell groups are located adjacent to the brachium conjunctivum.
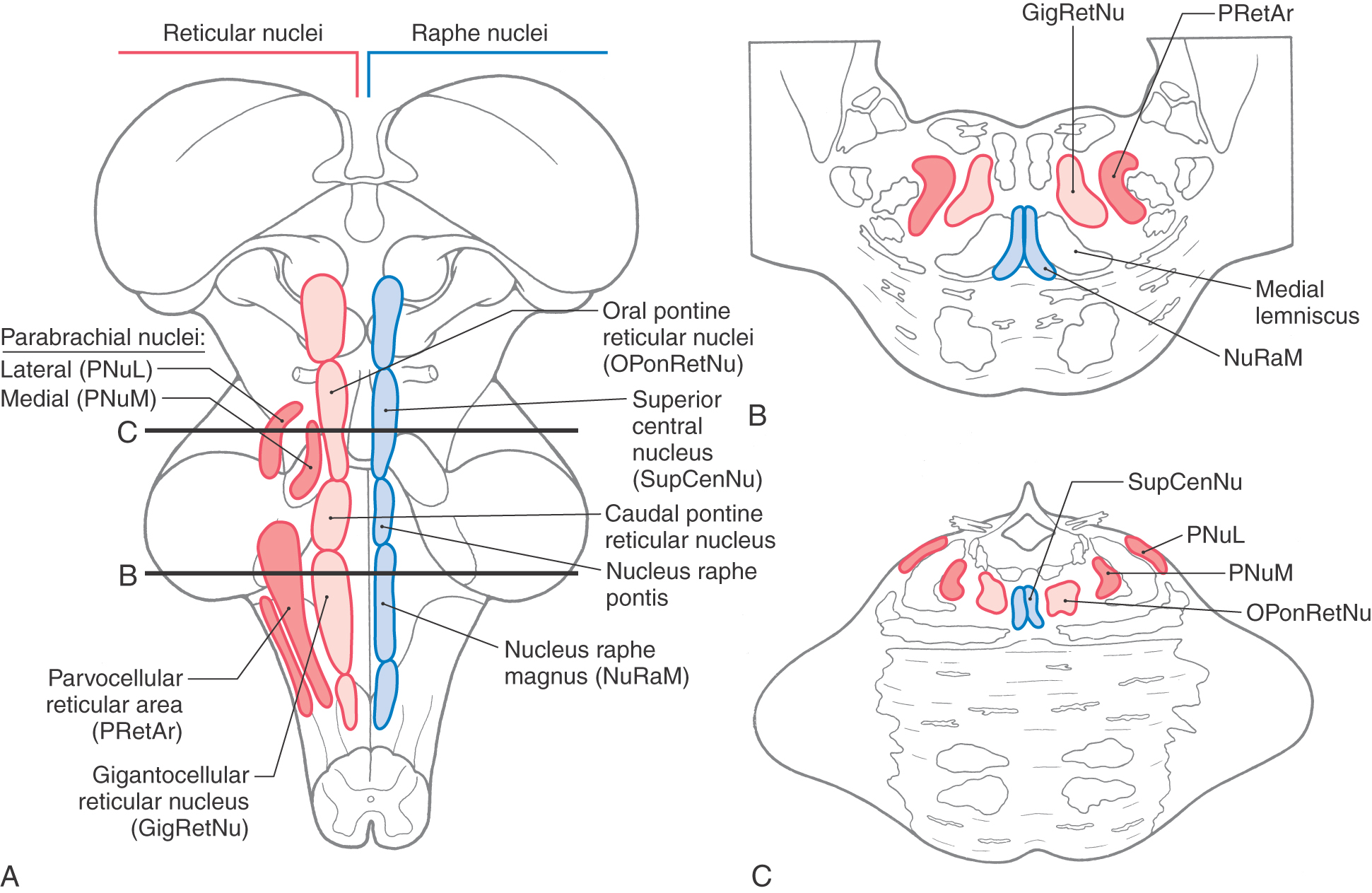
 Figure 12-17.
Figure 12-17. The raphe nuclei are symmetrically distributed on either side of the midline (Fig. 12-17). Next to the medial lemniscus at caudal pontine levels is the nucleus raphes magnus (the raphe magnus). This cell cluster extends caudally into the rostral medulla and is an important synaptic station for signals involved in the inhibition of pain at medullary and spinal levels. In the caudal third of the tegmental pons, this cell group is replaced by the nucleus raphes pontis, which extends a little beyond midpontine levels. The superior central nucleus and posterior (dorsal) raphe nucleus are found in the rostral pons; the latter extends into the caudal midbrain (see Fig. 13-16). The more caudal of the pontine raphe nuclei (magnus) project primarily to the spinal cord; the more rostral (pontis, superior central) project primarily rostrally to innervate a variety of forebrain targets. Although the main neurotransmitter associated with the pontine raphe nuclei is serotonin, there are some enkephalin-containing cells in the raphe magnus.
Internal Vasculature of the Pons
Internal areas of the tegmental and basilar pons are served by branches of the basilar artery (Fig. 12-18). Paramedian branches distribute to medial areas of the basilar pons, including corticospinal fibers and the exiting fibers of the abducens nerve. The lateral part of the basilar pons is served by short circumferential branches, and the entire tegmental area plus a wedge of the middle cerebellar peduncle receives blood via the long circumferential branches. At caudal levels (levels of the facial colliculus), the long circumferential supply is supplemented by branches of the anterior inferior cerebellar artery. Rostrally, beginning at about the level of the principal sensory and motor trigeminal nuclei, the blood supply to the pontine tegmentum is supplemented by branches of the superior cerebellar artery.
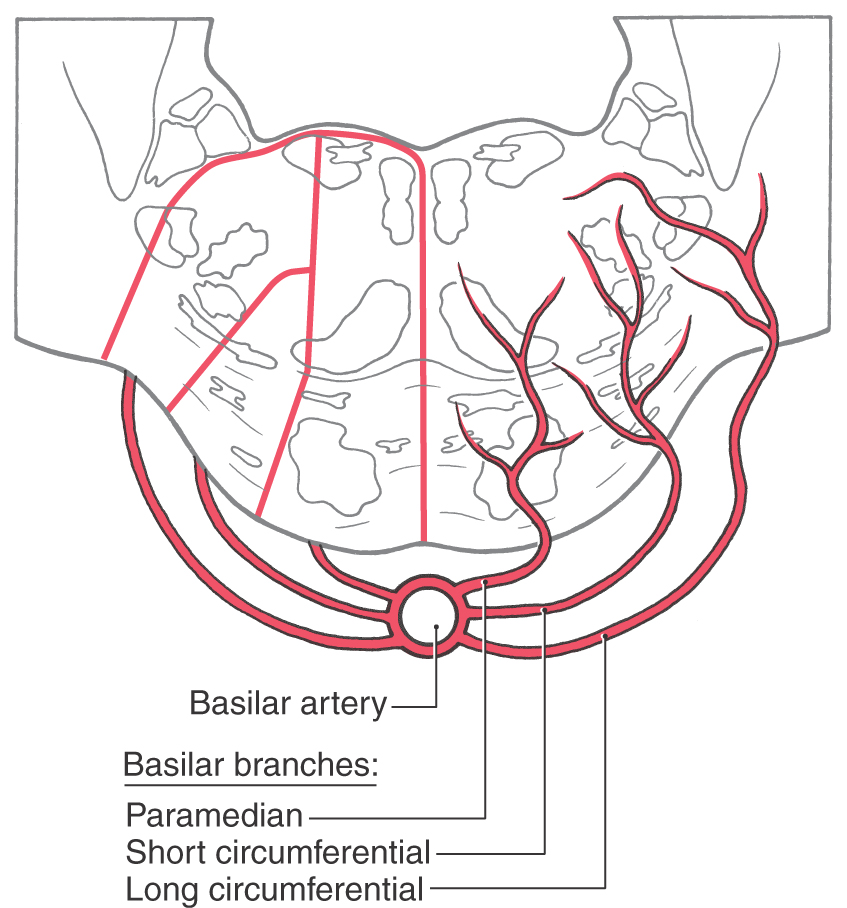
 Figure 12-18. Blood supply of the pons. Arteries are shown on the right, and the general territories served by each are shown on the left.
Figure 12-18. Blood supply of the pons. Arteries are shown on the right, and the general territories served by each are shown on the left.
VASCULAR SYNDROMES OF THE PONS
At caudal pontine levels (Figs. 12-11 and 12-18), the territory of the paramedian branches of the basilar artery includes the exiting fibers of the abducens nerve, corticospinal fibers, and, most likely, portions of the medial lemniscus. Occlusion of the paramedial branches at this level results in an ipsilateral abducens nerve paralysis and a contralateral hemiparesis (the Foville syndrome) with a variable contralateral sensory loss reflecting various degrees of damage to the medial lemniscus. If the lesion in the Foville syndrome extends posteriorly into the pontine tegmentum, the patient may have additional deficits, such as an ipsilateral horizontal gaze paralysis, indicating damage to the medial portions of the pontine reticular formation. If the area of damage is shifted somewhat laterally to include the root of the facial nerve along with corticospinal fibers, the patient has a contralateral hemiparesis and an ipsilateral paralysis of the facial muscles (the Gubler or Millard-Gubler syndrome).
At midpontine levels, occlusion of paramedial and short circumferential branches results in the syndrome of the midpontine base. The main structures damaged and their corresponding deficits are corticospinal fibers (contralateral hemiparesis), sensory and motor trigeminal roots (ipsilateral loss of pain and thermal sense and paralysis of the masticatory muscles), and fibers of the middle cerebellar peduncle (ataxia). Lesions within the pontine tegmentum may also combine cranial nerve signs and long tract signs. For example, a lesion in the caudal pontine tegmentum may damage the abducens and facial nuclei (paralysis of the ipsilateral lateral rectus muscle and facial musculature) and the anterolateral system (contralateral loss of pain and thermal sense on the body) (Fig. 12-11). At mid to more rostral pontine levels (Fig. 12-14), damage to the tegmentum may involve the anterolateral system in combination with the trigeminal motor nucleus or the sensory and motor roots of the trigeminal nerve. All of these examples illustrate a hallmark of brainstem vascular lesions, that is, an ipsilateral cranial nerve sign coupled with a contralateral long tract sign.
INTERNAL ANATOMY OF THE CEREBELLUM
Cerebellar Cortex
Each folium of the cerebellum is organized into three layers oriented parallel to the cortical surface. From external to internal, these are the molecular, Purkinje cell, and granular layers (Fig. 12-19). Internal to the granular layer, and forming the core of each folium, is a layer of subcortical white matter composed of all fibers arriving (afferents to the cortex) or leaving (efferents of the cortex) the cerebellar cortex.
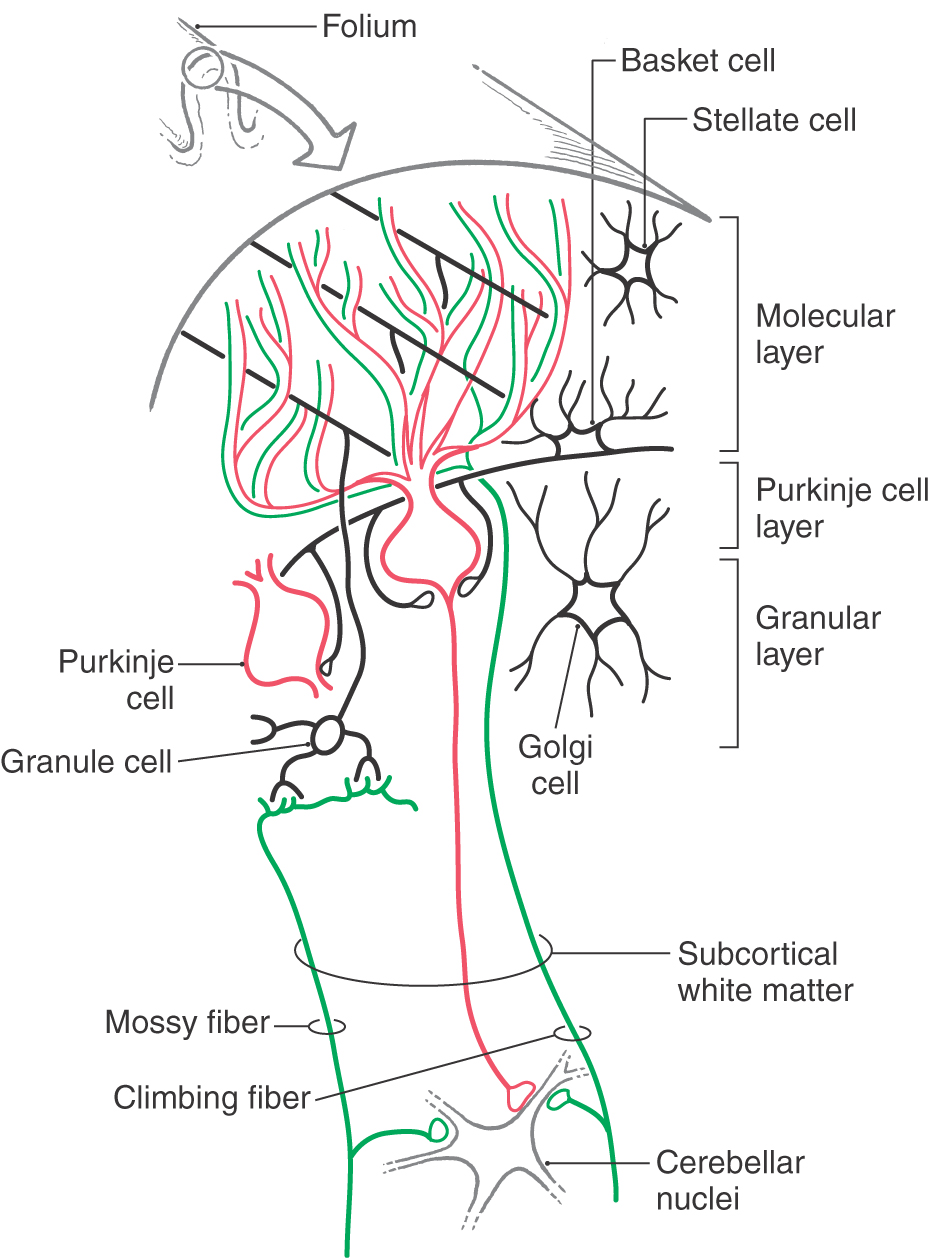
Figure 12-19. Diagrammatic representation of the cells and fibers of the cerebellar cortex.
The molecular layer contains stellate cells, basket cells, Golgi cell dendrites, and the expansive dendrites of Purkinje cells (Fig. 12-19). In addition, the molecular layer contains climbing fibers and parallel fibers. The climbing fibers originate from the contralateral inferior olive and the parallel fibers from the granule cells (Fig. 12-19).
Purkinje cells are the efferent neurons of the cerebellar cortex. Their large somata form a single layer at the molecular layer–granular layer interface (Fig. 12-19). Each Purkinje cell has a complex dendritic tree that extends into the molecular layer and exhibits a fan-like orientation that is perpendicular to the long axis of the folium. Axons of Purkinje cells pass through the subcortical white matter to end in the cerebellar and vestibular nuclei.
The granular layer contains granule cells, Golgi cells, and mossy fibers (Fig. 12-19). The mossy fibers originate from cells located in many nuclei throughout the brainstem and spinal cord. Traversing the granular layer are climbing fibers, en route to the molecular layer, and Purkinje cell axons leaving the cortex.
Cerebellar Nuclei
The cerebellar nuclei are, from medial to lateral, the fastigial, globose, emboliform, and dentate (Fig. 12-7). They receive input from Purkinje cell axons and from collaterals of cerebellar afferent fibers. In general, Purkinje cells of the vermis relate to the underlying fastigial nucleus, those of the intermediate cortex to the globose and emboliform nuclei, and those of the lateral cortex to the dentate nucleus (Fig. 12-7). Cerebellar nuclear cells project to a variety of cell groups throughout the neuraxis, primarily via the brachium conjunctivum.
Vasculature of the Cerebellum
The blood supply to the cerebellar cortex and nuclei arises via branches of the superior cerebellar artery and the anterior and posterior inferior cerebellar arteries (Fig. 12-7). The superior cerebellar artery serves the superior surface of the cerebellum; the fastigial, globose, and emboliform nuclei; and most areas of the dentate nucleus and the superior and middle cerebellar peduncles.
Branches of the anterior inferior cerebellar artery distribute to the more lateral areas of the inferior cerebellar surface (Fig. 12-7). They also serve parts of the middle cerebellar peduncle, a small caudal region of the dentate nucleus, and the choroid plexus in the subarachnoid space at the cerebellopontine angle.
The posterior inferior cerebellar artery (PICA) branches to more medial regions of the inferior cerebellar surface and to the nodulus (Fig. 12-7). The choroid plexus of the fourth ventricle receives branches from the PICA. This vessel is also an important source of blood to posterolateral regions of the medulla. The cerebellum is described in more detail in Chapter 27.
Sources and Additional Reading
Readings for the brainstem chapters are listed at the end of Chapter 13.