Chapter 11
The Medulla Oblongata
D.E. Haines and G.A. Mihailoff
Internal Anatomy of the Medulla
Summary of Descending Pathways
Spinal Cord–Medulla Transition
Caudal Medulla: Level of the Motor Decussation
Caudal Medulla: Level of the Sensory Decussation
Rostral Medulla and Pons-Medulla Junction
The medulla oblongata, or myelencephalon, is the most caudal segment of the brainstem. It extends from the level of the foramen magnum to the pons. The cavity of the medulla consists of a narrow, caudal part, which is the continuation of the central canal of the cervical spinal cord, and a flared, rostral portion, which is the medullary part of the fourth ventricle. The modest size of the medulla (0.5% of total brain weight) belies its importance. All the tracts passing to or from the spinal cord traverse the medulla, and 7 of the 12 cranial nerves (VI to XII) are associated with the medulla or the pons-medullary junction. Also, the medullary reticular formation contains cell groups that influence heart rate and respiration. The blood supply to the medulla arises from branches of the vertebral arteries.
DEVELOPMENT
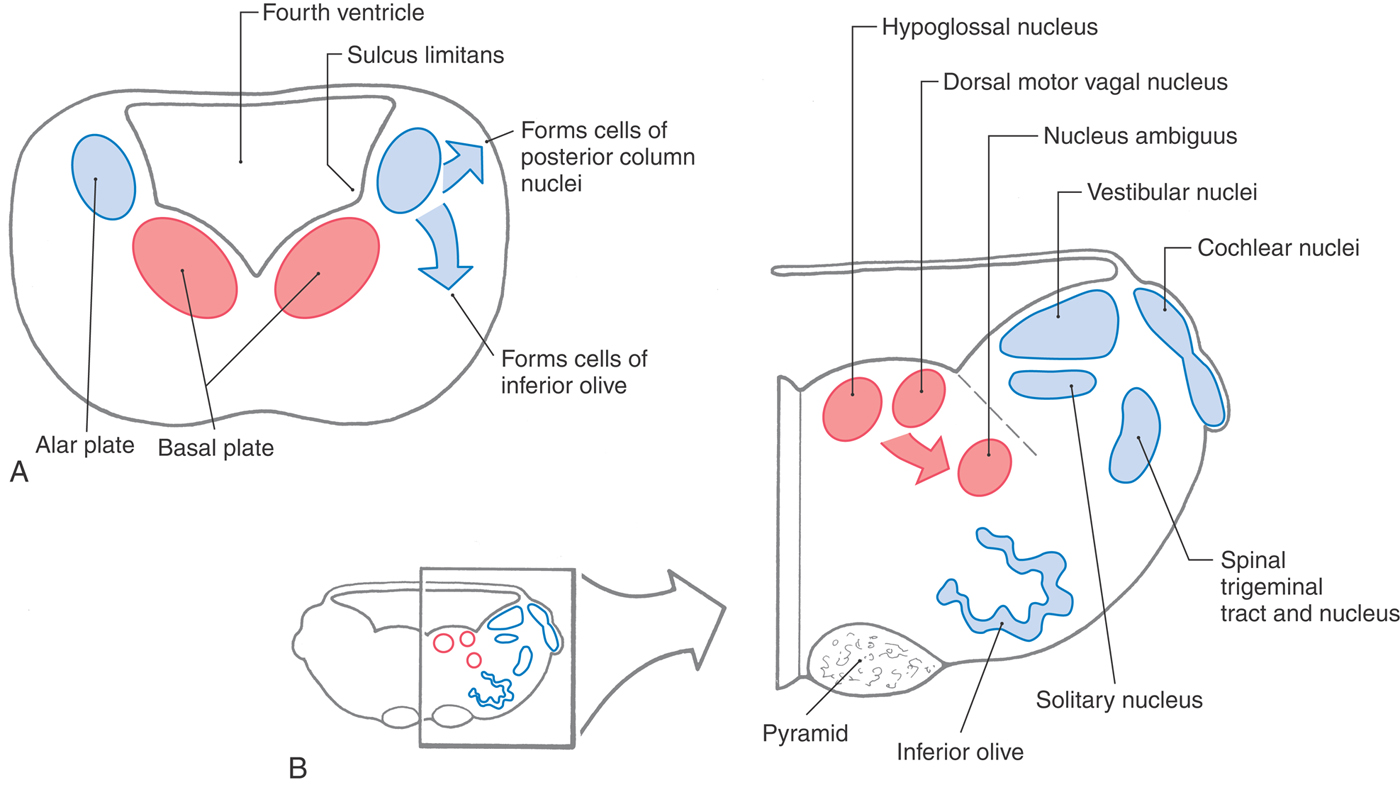
The basic structural plan of the medulla is an elaboration of that seen in the spinal cord (Figs. 11-1 and 11-2). The basal and alar plates give rise to specific nuclei, and the surrounding mantle layer is invaded by axons originating from other levels. Beginning in the medulla, however, the basic derivatives of the primitive neural tube are augmented by the appearance of other structures that characterize each brainstem level.
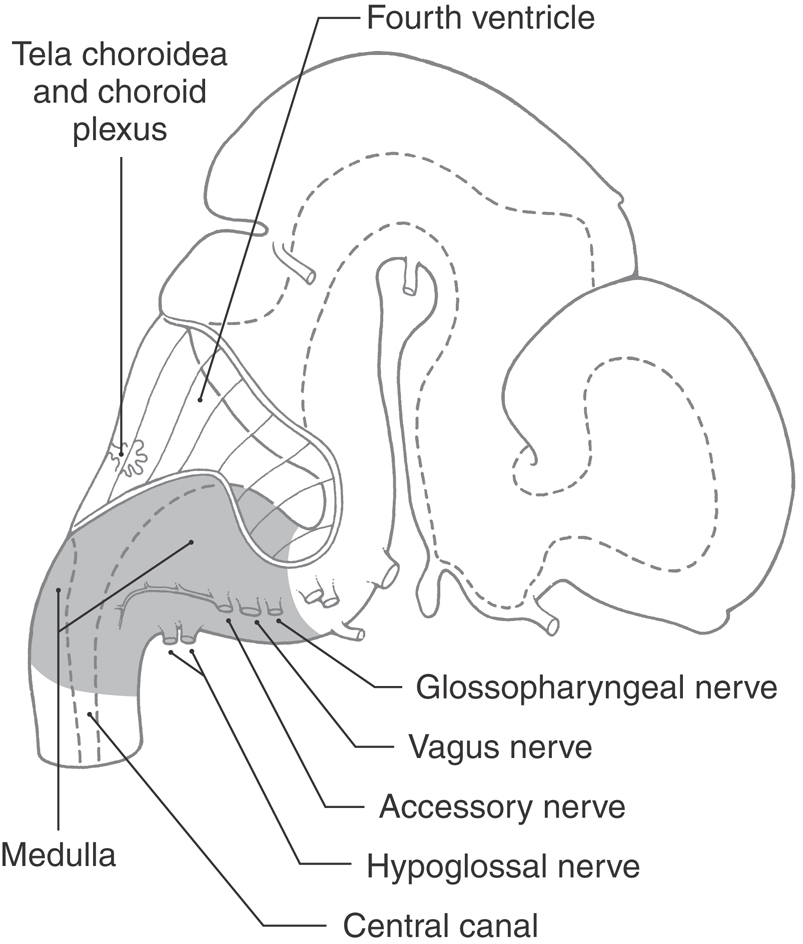
Figure 11-1. Lateral view of the brain at about 7 weeks of gestational age. The medulla is highlighted.
Basal and Alar Plates
Maturing neurons of the basal plate of the medulla give rise to the hypoglossal nucleus (somatic efferent [SE] cells), the dorsal motor vagal nucleus and the inferior salivatory nucleus (both contain visceral efferent [VE] cells), and the nucleus ambiguus (somatic efferent [SE] cells) (Fig. 11-2). Caudal to the obex, the hypoglossal and dorsal motor vagal nuclei are small and are found in the central gray surrounding the central canal. Rostral to the obex, all of these nuclei are located medial to the sulcus limitans (Fig. 11-2B).
The cranial nerve nuclei derived from the alar plate in the medulla and their corresponding functional components include the vestibular and cochlear nuclei (somatic afferent [SA]), the solitary nucleus (visceral afferent [VA]), and the spinal trigeminal nucleus (somatic afferent [SA]) (Fig. 11-2). Alar plate neuroblasts caudal to the obex give rise to the gracile and cuneate nuclei. Rostral to the obex, some alar plate cells migrate ventromedially to form the nuclei of the inferior olivary complex.
Concurrent with these developmental events, ascending and descending fibers are traversing the medulla. An especially prominent bundle of axons collects on the anterior (ventral) surface of the medulla to form the pyramids (Fig. 11-2B).
EXTERNAL FEATURES
Anterior Medulla
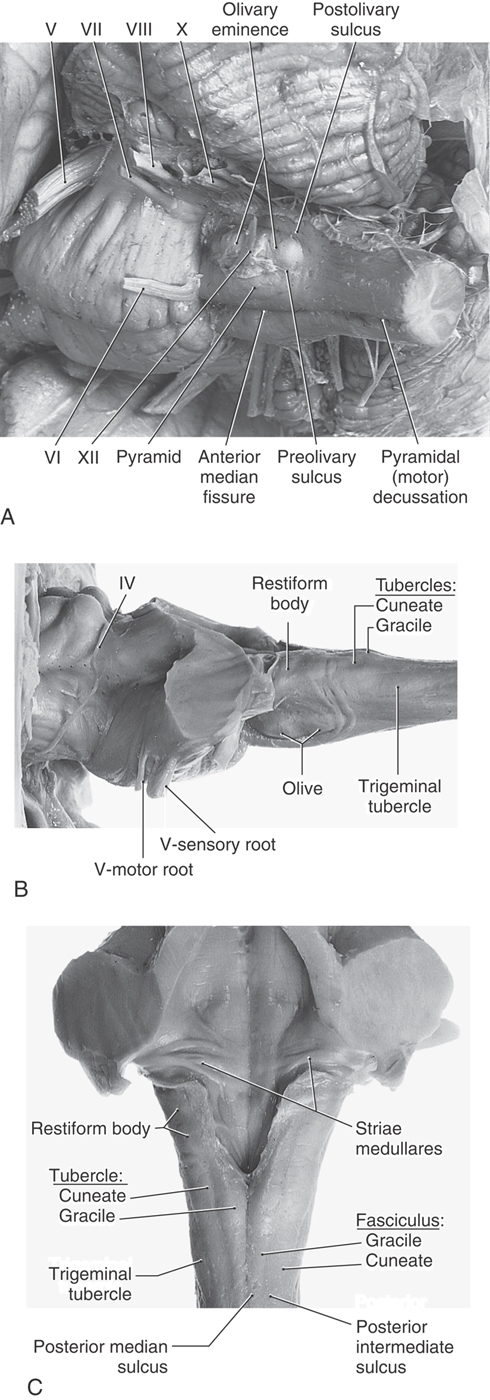
The anterior (ventral) aspect of the medulla is characterized by an anterior median fissure; two laterally adjacent longitudinal ridges, the pyramids; and the olive (inferior olivary eminence) (Fig. 11-3). The pyramids issue from the basilar pons and extend caudally to the motor (pyramidal) decussation, where about 90% of their fibers cross the midline. Most of the fibers that form the pyramid arise in the motor cortex as corticospinal fibers; consequently, their crossing is called the motor decussation. Rootlets of the hypoglossal nerve (cranial nerve XII) exit the medulla via the preolivary sulcus, a shallow groove located between the pyramid and the olive. The abducens nerve (cranial nerve VI) emerges at the pons-medullary junction, generally in line with the rootlets of cranial nerve XII.
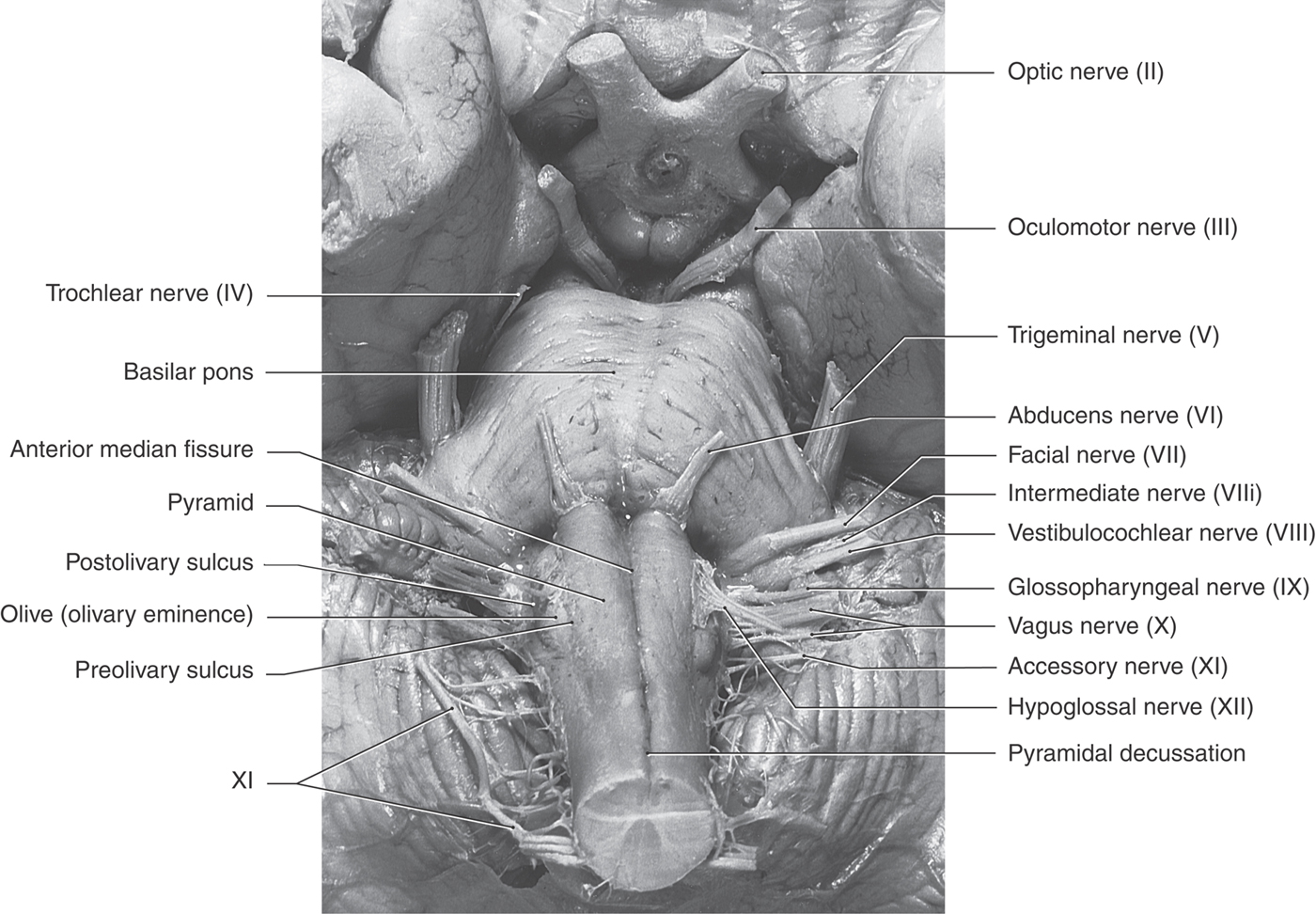
Figure 11-3. Anterior (ventral) view of the brainstem with emphasis on structures of the medulla.
Lateral Medulla
On the lateral aspect of the medulla, a shallow trough, the postolivary sulcus or retroolivary sulcus, is located between the restiform body and the large eminence formed by the underlying inferior olivary nucleus (Fig.11-4A, B). Cranial nerves IX (glossopharyngeal) and X (vagus) emerge from the postolivary sulcus. Caudal rootlets of the vagus have been incorrectly called the medullary, or bulbar, root of the accessory nerve. In actuality, the accessory nerve is made up of axons that arise from cells in the upper levels of the cervical spinal cord (C1 to C5 or C6), ascend through the foramen magnum, coalesce to form the accessory nerve, and then exit the skull via the jugular foramen along with the glossopharyngeal and vagus nerves. Structures served by the accessory nerve receive no innervation from the medulla. The facial nerve (VII), along with its intermediate root, and the vestibulocochlear nerve (VIII) emerge from the posterolateral medulla at the pons-medulla interface. The general region of the exit of the facial and vestibulocochlear nerves is clinically regarded as the cerebellopontine angle. Indeed, a vestibular schwannoma (often incorrectly referred to as an acoustic neuroma) is a tumor of the vestibular portion of the eighth cranial nerve and is a lesion located at the cerebellopontine angle. On the lateral medullary surface caudal to the level of the obex, fibers of the spinal trigeminal nucleus and tract assume a superficial location and form the trigeminal tubercle (tuberculum cinereum) (Fig. 11-4B, C). Rostral to the obex, these trigeminal fibers are located internal to a progressively enlarging restiform body.
Posterior Medulla
At and caudal to the level of the obex, the posterior surface of the medulla is characterized by the gracile and cuneate fasciculi and their respective tubercles (Fig. 11-4C). These tubercles are formed by the underlying gracile and cuneate nuclei. Rostrolateral to the gracile and cuneate tubercles and forming a prominent elevation on the posterolateral aspect of the medulla is the restiform body. This structure contains a variety of afferent cerebellar fibers and becomes progressively larger as it extends toward the pons-medulla junction. In the caudal pons, fibers of the restiform body join with a much smaller bundle, the juxtarestiform body, to form the inferior cerebellar peduncle.
Vasculature
In general, the blood supply to the entire medulla and to the choroid plexus of the fourth ventricle arises from branches of the vertebral arteries (see Fig. 11-16). The exceptions are the portion of the choroid plexus that extends out of the foramen of Luschka and the adjacent cochlear nuclei; these are served by branches of the anterior inferior cerebellar artery, a branch of the basilar artery. In general, the medial medulla is served by the anterior spinal artery, the anterolateral medulla by small branches from the vertebral artery, and the posterolateral medulla rostral to the obex by the posterior inferior cerebellar artery (PICA). Caudal to the obex, the posterior medulla is served by the posterior spinal artery.
The vascular territory of the anterior spinal artery encompasses the medial lemniscus, hypoglossal root, and corticospinal fibers in the pyramid. Consequently, lesions of this vascular region result in somatosensory and motor deficits reflecting damage to these structures. In similar manner, lesions in the territory of the PICA will damage the anterolateral system and spinal trigeminal tract and nucleus and give rise to deficits reflecting these structures.
INTERNAL ANATOMY OF THE MEDULLA
Summary of Ascending Pathways
The ascending tracts that originate from the spinal cord gray matter (anterolateral system, posterior and anterior spinocerebellar tracts) and from posterior root ganglion cells (gracile and cuneate fasciculi) continue into the medulla (Fig. 11-5). Some anterolateral system fibers terminate in the medulla, as spinoreticular fibers, and others convey pain and temperature input to more rostral levels, including the thalamus, as spinomesencephalic and spinothalamic fibers. Posterior column fibers synapse in the medulla, but the tactile and vibratory information carried by these fibers continues rostrally via the medial lemniscus (Fig. 11-5). Spinocerebellar axons enter the cerebellum through the restiform body (posterior tract) or the superior cerebellar peduncle (anterior tract). Other ascending bundles, such as spinoolivary and spinovestibular fibers, terminate in the medulla.
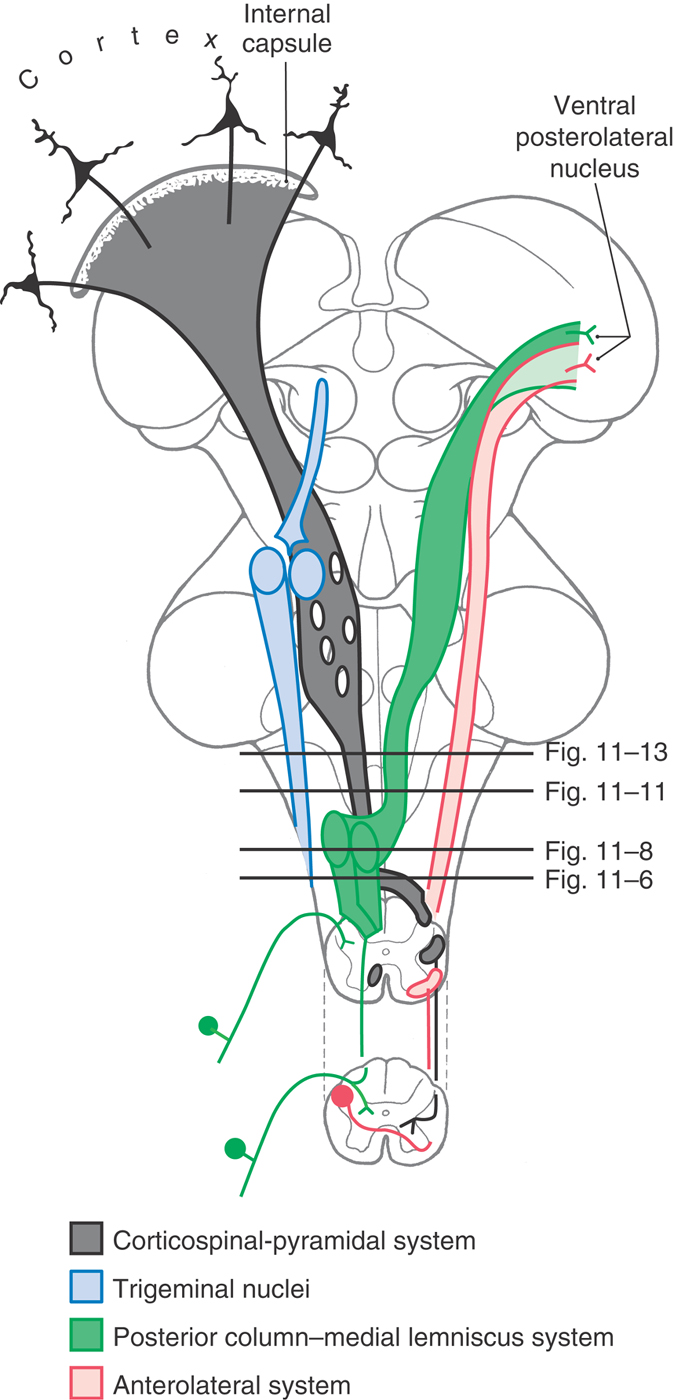
Figure 11-5. Diagram of the brain showing the location and trajectory of three important pathways and the trigeminal nuclei. The color coding for each is continued in Figures 11-6, 11-8, 11-11, and 11-13.
Summary of Descending Pathways
The descending tracts that originate from the cerebral cortex (corticospinal, Fig. 11-5) and from the midbrain (rubrospinal, tectobulbospinal) and pons (reticulospinal, vestibulospinal) traverse the medulla en route to the spinal cord. The medulla contributes additional fibers to the last two fiber systems. At this level, the medial longitudinal fasciculus contains only descending fibers. The majority of these descending axons influence, either directly or indirectly through interneurons, the discharge patterns of motor neurons in the spinal cord gray matter.
Spinal Cord–Medulla Transition
The spinal cord–medulla transition is characterized by changes that begin at the caudal level of the motor decussation (Figs. 11-6 and 11-7). The spinal cord gray matter is replaced by the motor (crossing of corticospinal fibers) decussation; the central gray matter enlarges; the posterolateral tract (dorsolateral fasciculus) and substantia gelatinosa of the spinal cord merge, respectively, into the spinal trigeminal tract and nucleus; and nuclei characteristic of the medulla appear. The caudal medulla is described in the following sections beginning at the levels of the motor and sensory decussations.
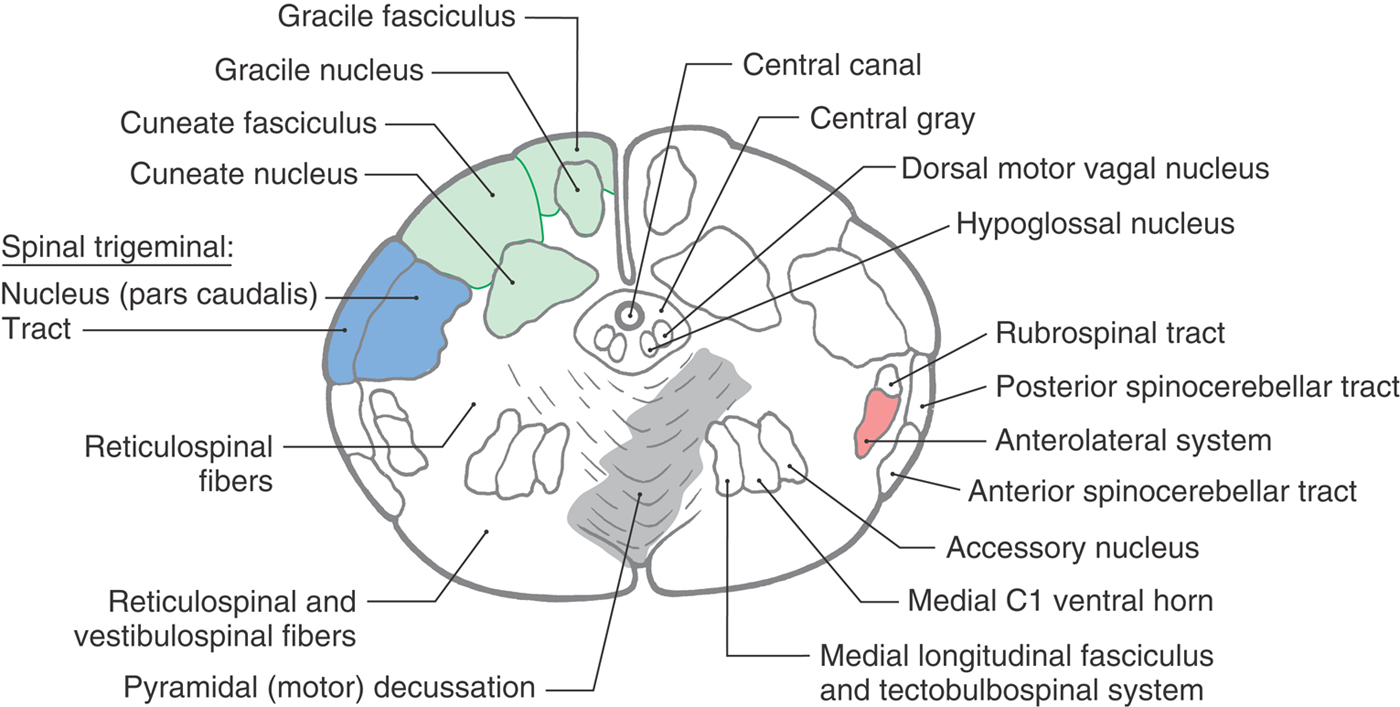
 Figure 11-6. Cross section of the medulla at the level of the motor decussation. Correlate with Figure 11-5.
Figure 11-6. Cross section of the medulla at the level of the motor decussation. Correlate with Figure 11-5.
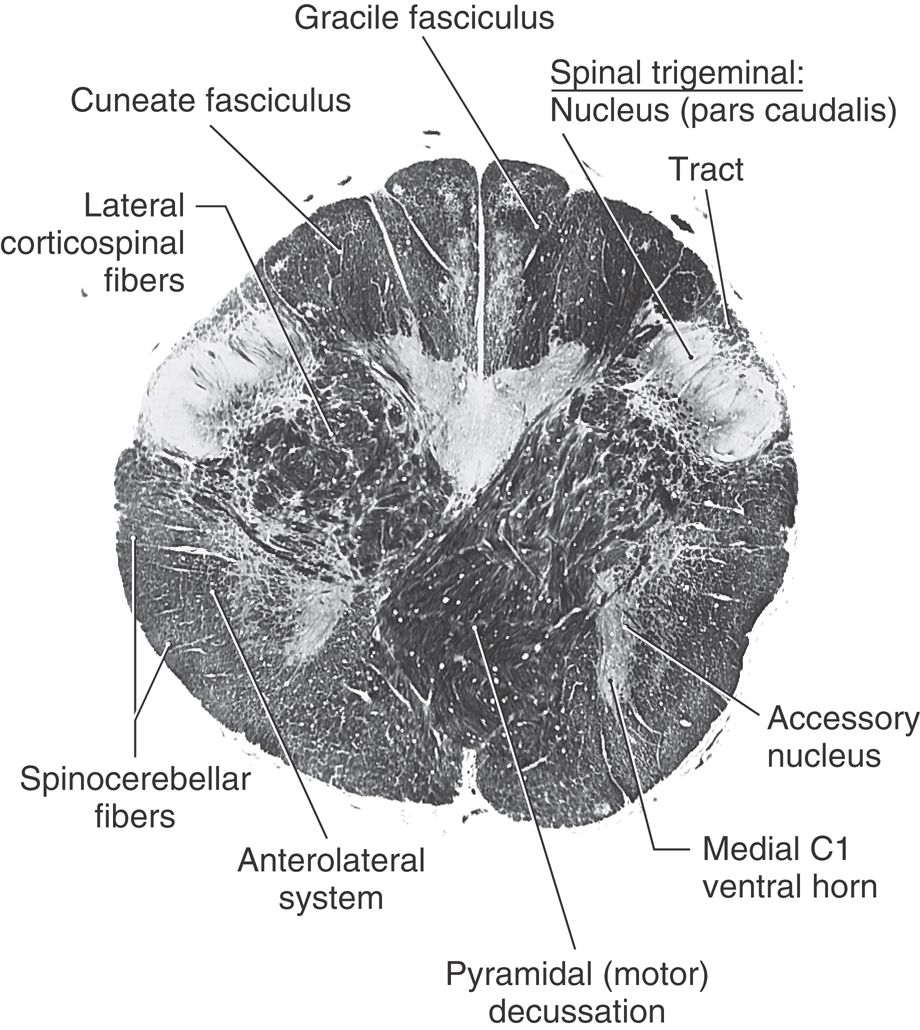
 Figure 11-7. A fiber (myelin)–stained cross section of the medulla at the level of the motor decussation. Compare with Figure 11-6. (From Parent A: Carpenter’s Human Neuroanatomy, 9th ed. Philadelphia, Lippincott Williams & Wilkins, 1995.)
Figure 11-7. A fiber (myelin)–stained cross section of the medulla at the level of the motor decussation. Compare with Figure 11-6. (From Parent A: Carpenter’s Human Neuroanatomy, 9th ed. Philadelphia, Lippincott Williams & Wilkins, 1995.)
Caudal Medulla: Level of the Motor Decussation
At the level of the motor decussation (pyramidal decussation), about 90% of corticospinal fibers cross the anterior midline to form the contralateral lateral corticospinal tract of the cord (Figs. 11-5 to 11-7). Posteriorly, at this level, the gracile and cuneate nuclei first appear in their respective fasciculi (Figs. 11-6 and 11-7). Because the gracile and cuneate fasciculi are collectively called the posterior (or dorsal) columns, their respective nuclei are frequently referred to as the posterior column nuclei. Laterally, the spinal trigeminal tract is visible on the surface of the medulla as the trigeminal tubercle or tuberculum cinereum. Internal to the spinal trigeminal tract is the spinal trigeminal nucleus, pars caudalis (Fig. 11-6).
The spinal trigeminal tract is composed of central processes of primary sensory fibers that enter the brain mainly in the trigeminal nerve. This tract also receives fibers that originate from cranial nerves VII, IX, and X. These primary sensory fibers terminate on cells of the spinal trigeminal nucleus, which in turn projects to the contralateral thalamus as the anterior (ventral) trigeminothalamic tract.
In the lateral medulla, the anterolateral system and rubrospinal tract are found medial to the superficially located posterior and anterior spinocerebellar tracts (Figs. 11-6 and 11-7). It is important to emphasize that anterolateral system fibers (conveying pain and temperature input from the contralateral side of the body) and spinal trigeminal tract fibers (conveying pain and temperature from the ipsilateral face) are located adjacent to each other throughout the lateral area of the medulla.
The anterior medulla contains the most rostral part of the accessory nucleus (cranial nerve XI), remnants of the medial motor cell column of C1, and the medial longitudinal fasciculus and tectobulbospinal system. The most rostral remnants of the accessory nucleus (cranial nerve XI) and the medial motor cell column of C1 are seen at the spinal cord–medulla junction but do not extend into the medulla. Immediately adjacent to these cell groups are the small fiber bundles of the medial longitudinal fasciculus and the tectobulbospinal system (Figs. 11-6 and 11-7). At this level, the tectospinal fibers in the tectobulbospinal system are incorporated into the medial longitudinal fasciculus. These small bundles are displaced laterally by the motor decussation compared with their medial positions at more rostral levels.
The central gray surrounds the central canal of the medulla and contains the caudal extremes of the hypoglossal (XII) and dorsal motor vagal (X) nuclei (Fig. 11-6). When the ventricle flares open at the level of the obex, these nuclei occupy the medial floor of the ventricular space.
Caudal Medulla: Level of the Sensory Decussation
The sensory decussation is so named because it is the point at which a major ascending sensory pathway (posterior column–medial lemniscus) crosses the midline. At this level of the medulla, the posterior columns (gracile and cuneate fasciculi) are largely replaced by the gracile and cuneate nuclei (Figs. 11-8 and 11-9). Fibers conveying tactile and vibratory sensations from lower and upper levels of the body terminate, respectively, in the gracile and cuneate nuclei. The axons of these cells swing anteromedially to form the internal arcuate fibers, which in turn cross the midline as the sensory decussation and collect to form the medial lemniscus on the contralateral side (Figs. 11-5, 11-8, and 11-9). Information from the lower extremities (gracile cell axons) is conveyed in the anterior part of the medial lemniscus, and information from the upper extremities (cuneate cell axons) is conveyed in the posterior part of the medial lemniscus (see Fig. 12-13). The accessory cuneate nucleus is located lateral to the cuneate nucleus (Fig. 11-8). Its cells receive primary sensory input via cervical spinal nerves and project to the cerebellum as cuneocerebellar fibers. In doing so, they represent the upper extremity equivalent of the posterior spinocerebellar tract.
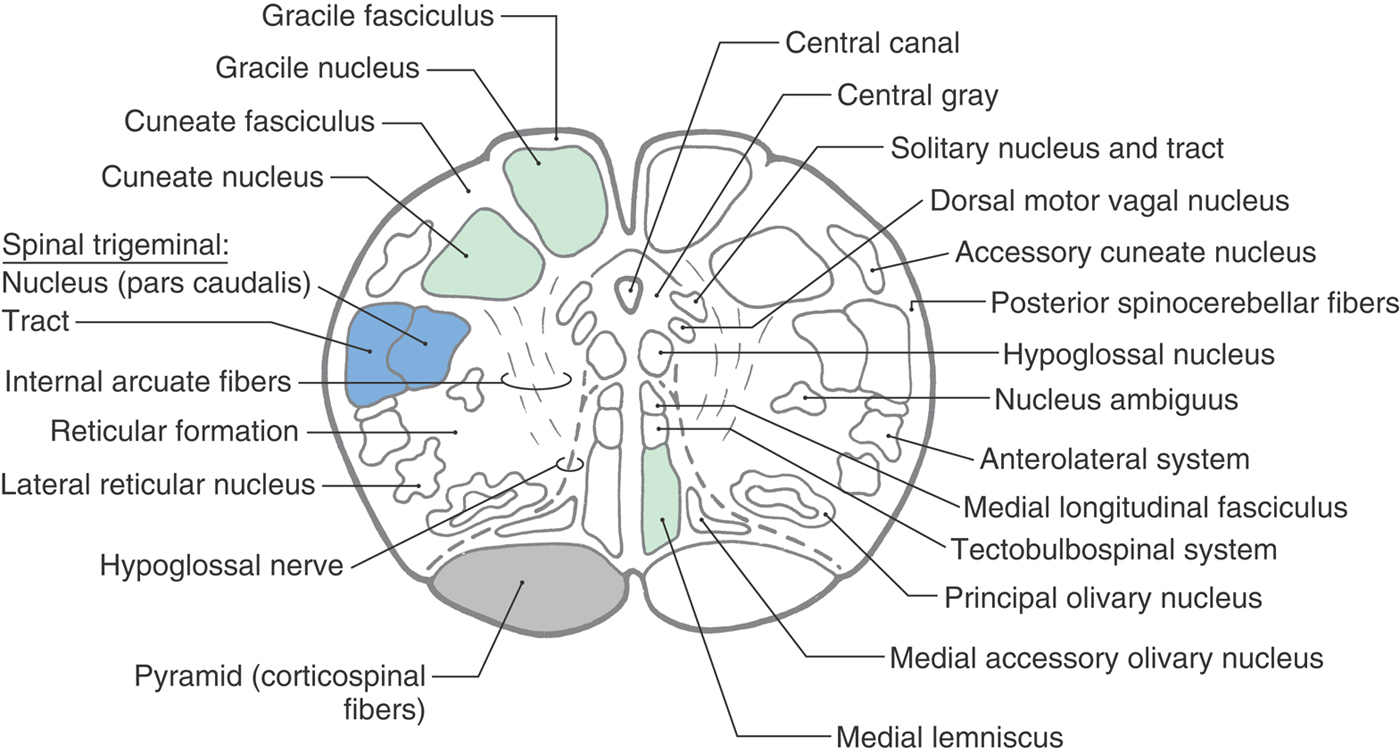
 Figure 11-8. Cross section of the medulla at the level of the sensory decussation. Correlate with Figure 11-5.
Figure 11-8. Cross section of the medulla at the level of the sensory decussation. Correlate with Figure 11-5.
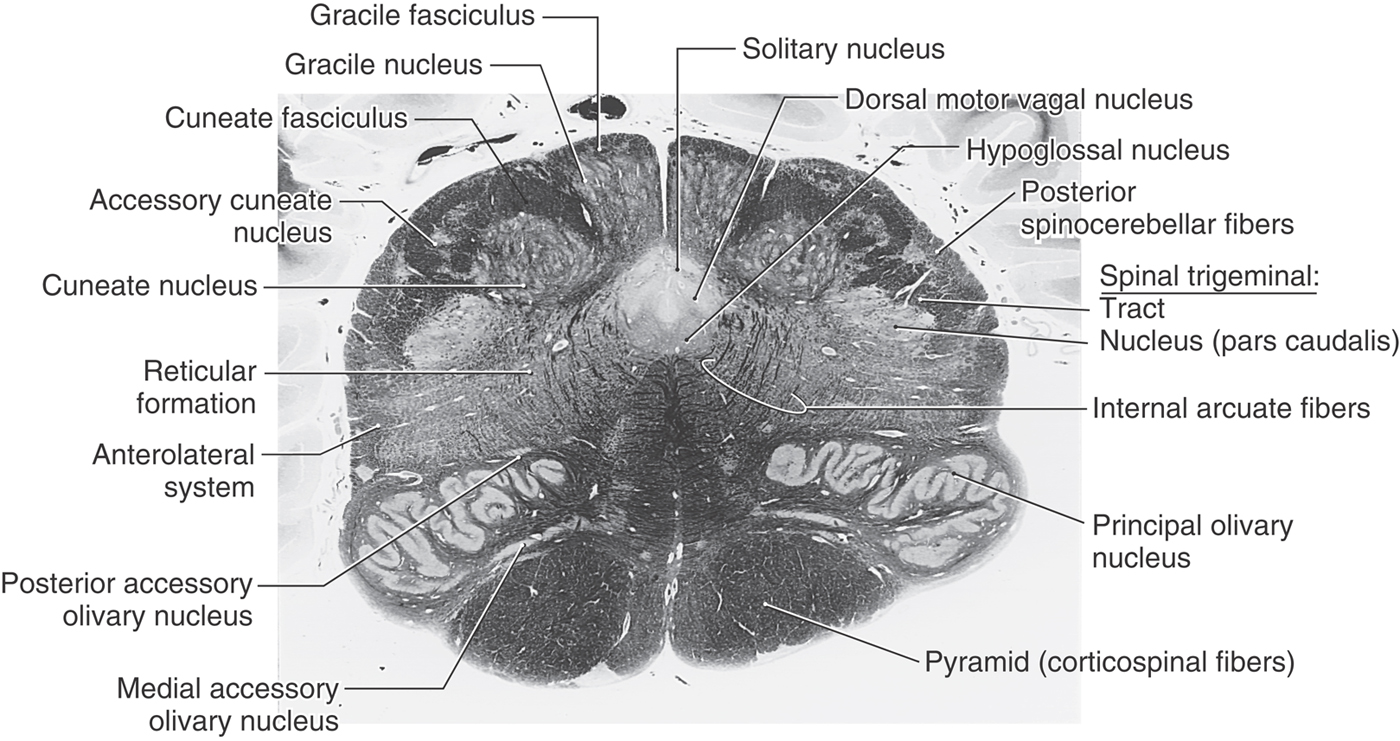
 Figure 11-9. A fiber (myelin)–stained cross section of the medulla at the level of the sensory decussation. Compare with Figure 11-8.
Figure 11-9. A fiber (myelin)–stained cross section of the medulla at the level of the sensory decussation. Compare with Figure 11-8.
The spinal trigeminal tract and nucleus (pars caudalis) maintain their position in the lateral medulla. The pars caudalis is that portion of the spinal trigeminal nucleus located caudal to the level of the obex. At this level, however, posterior spinocerebellar fibers have migrated posteriorly to cover the spinal tract, heralding the beginnings of the restiform body (Figs. 11-8 and 11-9). Just medial to the spinal trigeminal nucleus, a small column of motor neurons, the nucleus ambiguus, appears (Fig. 11-8). The axons of these SE cells travel in the glossopharyngeal (IX) and vagus (X) nerves. Fibers of the anterolateral system and rubrospinal tract are located in the anterolateral medulla (Fig. 11-8). The lateral reticular nucleus, a distinct cell group adjacent to the anterolateral system, receives spinal input and projects to the cerebellum.
Structures characteristic of the anterior surface of the medulla at this level include the pyramid, fibers of the hypoglossal nerve, and the caudal end of the inferior olivary complex (Figs. 11-8 and 11-9). The inferior olivary nuclei (internal to the olivary eminence), which become larger at more rostral levels, receive input from a variety of areas and project primarily to the cerebellum. The inferior olivary nuclei are sometimes described as the inferior olivary complex because they collectively consist of principal, medial accessory, and posterior accessory nuclei. Internal to the pyramid, and along the midline from anterior to posterior, are the medial lemniscus, tectobulbospinal fibers, and medial longitudinal fasciculus (Fig. 11-8). At this level, medial longitudinal fasciculus fibers are characteristically found adjacent to the midline and anterior to structures of the central gray.
The central gray is larger than at the level of the motor decussation, and caudal parts of the hypoglossal and dorsal motor vagal nuclei and the solitary nucleus and tract can be clearly identified along its perimeter (Figs. 11-8 and 11-9). Hypoglossal (SE) motor neurons innervate the ipsilateral half of the tongue. These fibers course anterolaterally along the lateral edge of the medial lemniscus and pyramid and share a common blood supply with these structures. The VE cells of the dorsal motor vagal nucleus provide preganglionic parasympathetic fibers to visceromotor ganglia (autonomic ganglia), the postganglionic fibers of which innervate viscera in the thorax and abdomen. The solitary tract and nucleus receive VA input from cranial nerves VII, IX, and X. At this caudal level of the medulla, input to the solitary nucleus is primarily visceral sensation from thoracic and abdominal viscera (via cranial nerve X) and the carotid sinus (via cranial nerve IX) and terminates in the cardiorespiratory portion of the solitary nucleus.
The fourth ventricle flares open at the level of the obex (Fig. 11-10). The area postrema is an emetic (vomiting) center located in the wall of the ventricle at this level. Especially noticeable changes at this point compared with more caudal levels include enlargement of the inferior olivary complex and restiform body.
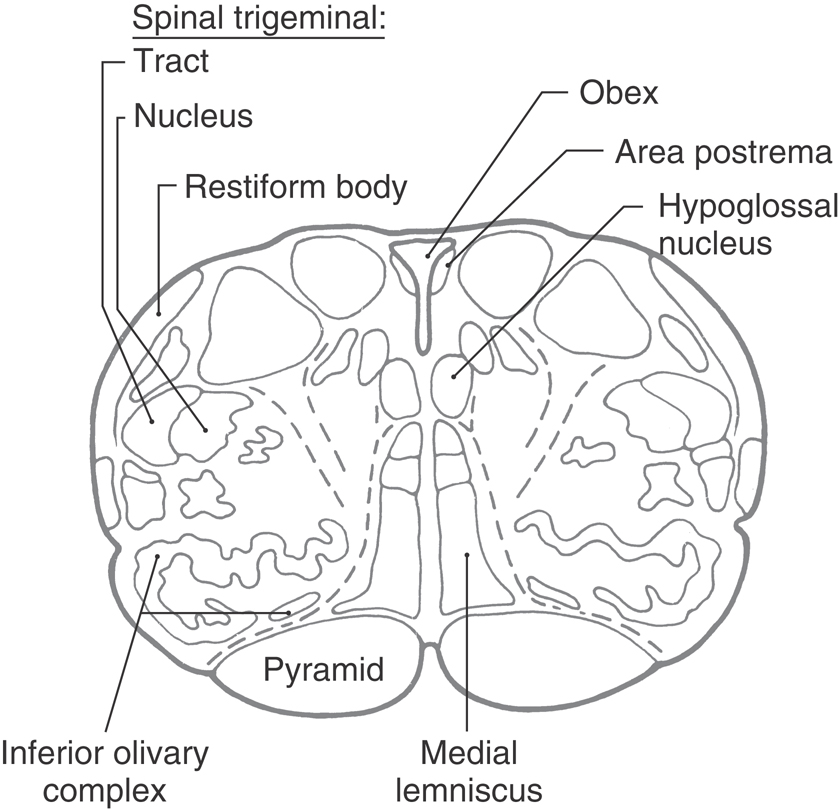
 Figure 11-10. Cross section of the medulla at the level of the obex.
Figure 11-10. Cross section of the medulla at the level of the obex.
Midmedullary Level
The structures in the medial floor of the fourth ventricle are, rostral to the obex, the hypoglossal and dorsal motor vagal nuclei and, lateral to the sulcus limitans, the vestibular nuclei (Figs. 11-11 and 11-12). The latter cell groups consist, at this level, of medial and inferior (or spinal) vestibular nuclei. They receive input from cranial nerve VIII and interconnect with areas of the brain concerned with balance, equilibrium, and eye movement. The solitary tract and nucleus occupy their characteristic position immediately inferior to the vestibular nuclei.
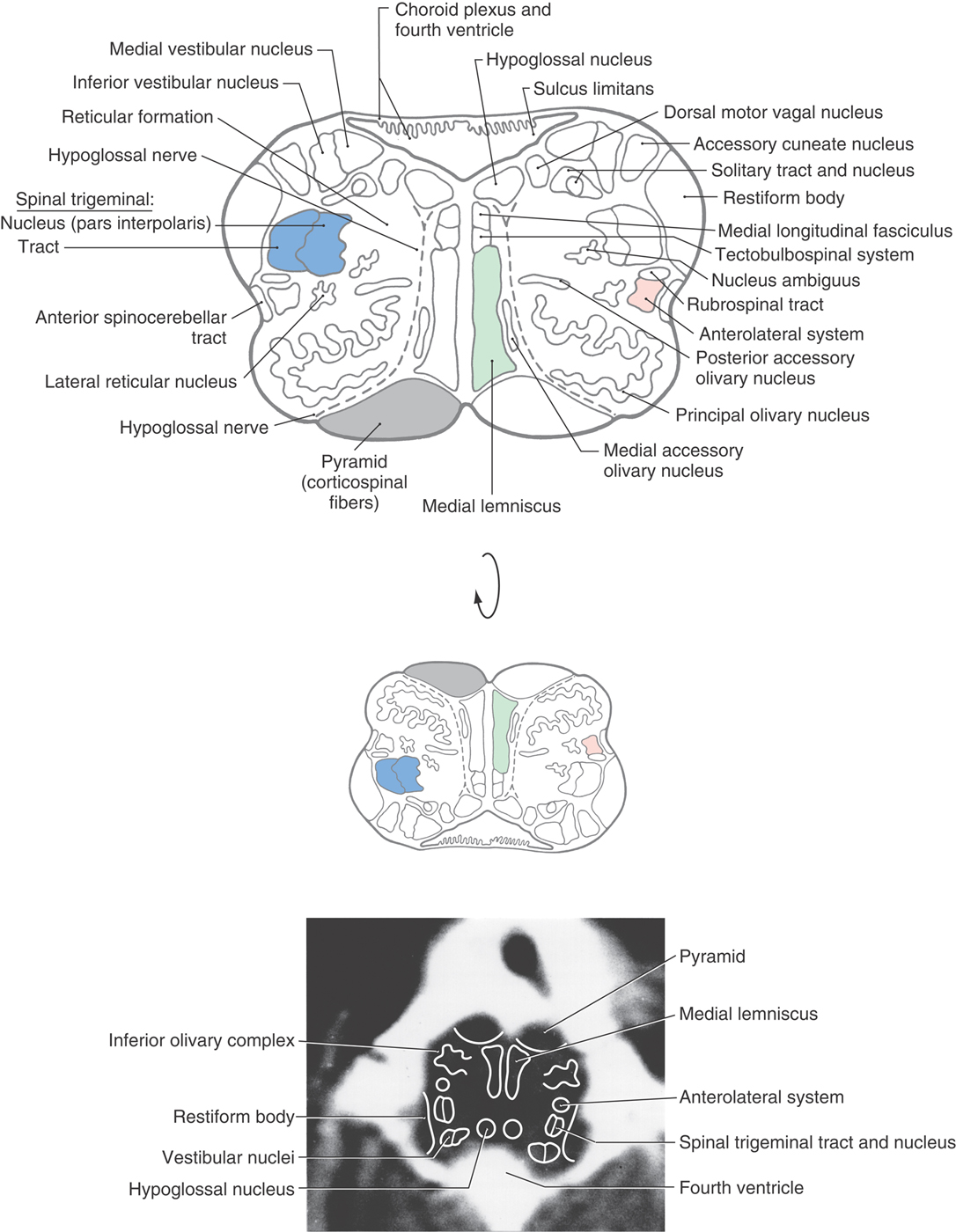
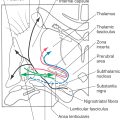
Figure 11-11. Cross section of the medulla at midolivary levels. Correlate with Figure 11-5. The anatomic orientation is flipped to illustrate internal structures in a clinical orientation; the clinically important tracts and nuclei are shown on a T2-weighted magnetic resonance image at a comparable level of the mid medulla.
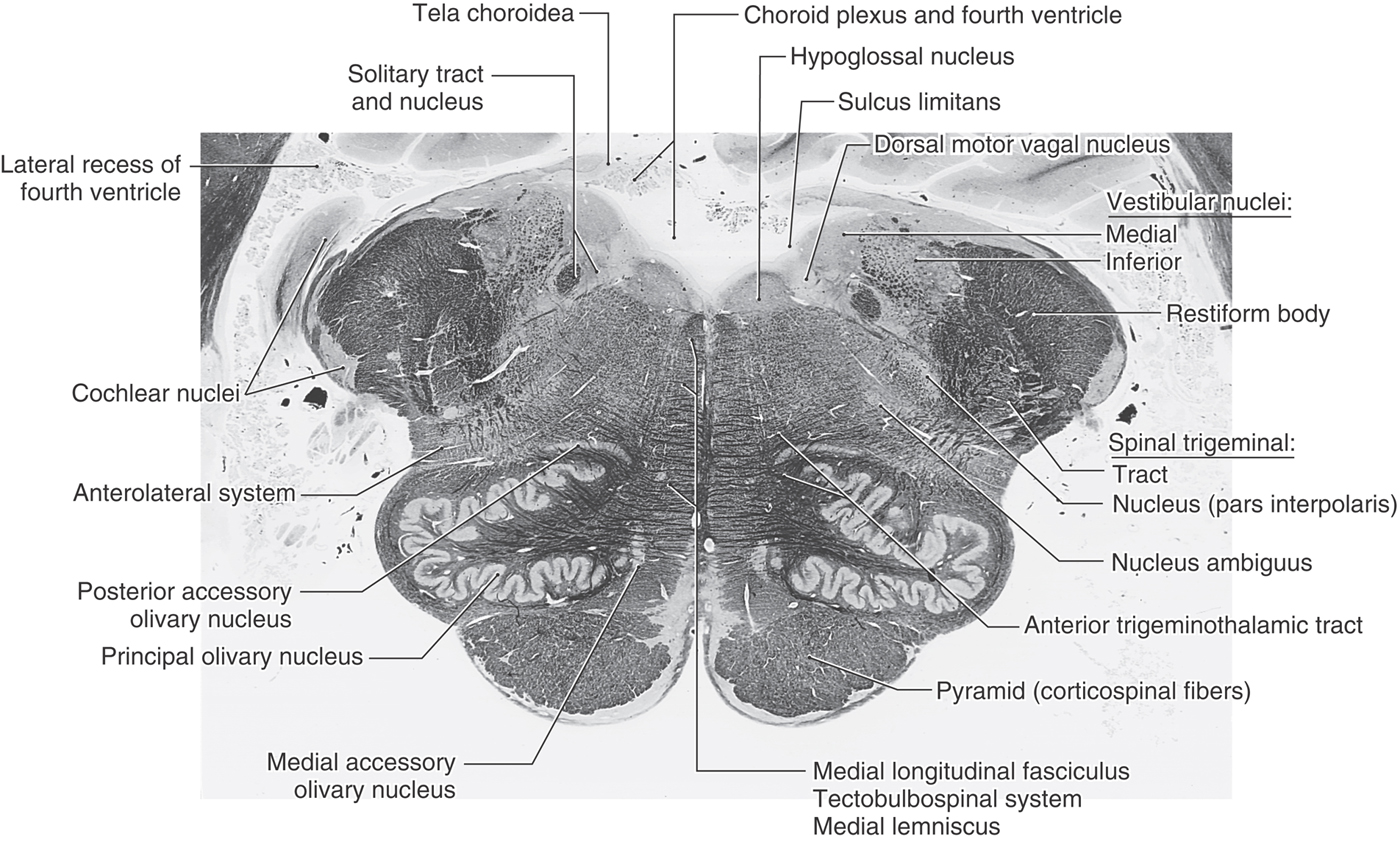
 Figure 11-12. A fiber (myelin)–stained cross section of the medulla at midolivary levels. This section is between the levels represented in Figures 11-11 and 11-13.
Figure 11-12. A fiber (myelin)–stained cross section of the medulla at midolivary levels. This section is between the levels represented in Figures 11-11 and 11-13.
Laterally, the restiform body forms a prominent elevation on the posterolateral aspect of the medulla (Figs. 11-11 and 11-12). This structure contains posterior spinocerebellar, cuneocerebellar, olivocerebellar, reticulocerebellar, and other cerebellar afferents. In the base of the cerebellum, these fibers join with the juxtarestiform body to form the inferior cerebellar peduncle.
The spinal trigeminal tract and nucleus (pars interpolaris) are internal to the restiform body (Figs. 11-11 and 11-12). The pars interpolaris is the part of the spinal trigeminal nucleus located between the levels of the obex and the rostral end of the hypoglossal nucleus. Other structures in the lateral medulla are comparable to those seen more caudally. These include the nucleus ambiguus and the lateral reticular nucleus as well as the anterolateral system, anterior spinocerebellar tract, and rubrospinal tract (Fig. 11-11). At all medullary levels, neurons of the nucleus ambiguus contribute axons to cranial nerves IX and X, which innervate pharyngeal and laryngeal muscles, including those of the vocal folds.
Anterolaterally, the inferior olivary complex is prominent at midmedullary levels and is composed of a large, saccular principal olivary nucleus and diminutive medial and posterior accessory olivary nuclei (Figs. 11-11 and 11-12). These cell groups receive input from a variety of central nervous system nuclei and project primarily to the contralateral cerebellum (as olivocerebellar fibers) through the restiform body. Anteriorly and medially, the orientation of the pyramid, medial lemniscus, medial longitudinal fasciculus, and tectobulbospinal system remains essentially the same as at more caudal levels (Figs. 11-11 and 11-12).
Rostral Medulla and Pons-Medulla Junction
Comparison of Figures 11-11 and 11-13 shows that many of the structures seen in the mid medulla are present in essentially the same locations in the rostral medulla. Therefore, we shall emphasize the features that are different in the rostral medulla.
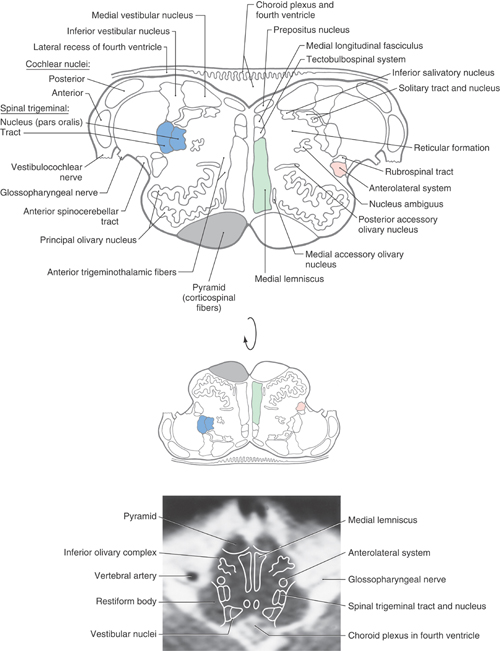
Figure 11-13. Cross section of the medulla at rostral olivary (and medullary) levels. Correlate with Figure 11-5. The anatomic orientation is flipped to illustrate internal structures in a clinical orientation; the clinically important tracts and nuclei are shown on a T2-weighted magnetic resonance image at a comparable level of the rostral medulla.
In the floor of the fourth ventricle, the positions occupied by the hypoglossal and dorsal motor vagal nuclei at more caudal levels are taken by the prepositus (hypoglossal) nucleus and the inferior salivatory nucleus (Fig. 11-13). The prepositus nucleus is a small, somewhat flattened cell group that is easily distinguished from the hypoglossal nucleus. The VE cells of the inferior salivatory nucleus are located immediately inferior to the medial vestibular nucleus and medial to the solitary tract and nucleus. Axons of these salivatory nucleus cells distribute to the otic ganglion via peripheral branches of the glossopharyngeal nerve.
The medial and inferior (or spinal) vestibular nuclei are prominent at this level and are joined, in this plane of section, by the posterior and anterior cochlear nuclei (Figs. 11-12 and 11-13). The latter nuclei are located on the posterior and lateral aspects of the restiform body at the pons-medullary junction. The medial vestibular nucleus appears homogeneous in fiber-stained sections, and the inferior vestibular nucleus has a salt-and-pepper appearance (Fig. 11-12). This appearance results because small descending bundles of myelinated fibers (pepper) are intermingled with cells (salt) in the inferior nucleus. Medial to the restiform body is the spinal trigeminal tract and the pars oralis of the spinal trigeminal nucleus (Fig. 11-13). The pars oralis is that part of the nucleus located rostral to the level of the hypoglossal nucleus.
At this rostral level of the medulla, the solitary tract and nucleus retain their positions immediately inferior to the medial and inferior vestibular nuclei. However, in contrast to the more caudal parts of this nucleus (which receives mostly VA input originating mainly from cranial nerves IX and X), this rostral portion of the nucleus receives primarily VA (taste) input from nerves VII and IX; this is the gustatory nucleus. The solitary tract and nucleus do not extend craniad to the root of the facial nerve.
Although structures in anterior and medial areas of the medulla are unchanged from those at midmedullary levels, some changes take place at the pons-medullary junction that merit comment (Fig. 11-14). Fibers of the restiform body arch posteriorly to enter the cerebellum, where they are joined by fibers of the juxtarestiform body to form (collectively) the inferior cerebellar peduncle. The facial motor nucleus appears anterolaterally, and the trapezoid body and superior olivary nucleus (both conveying auditory information) appear adjacent to the facial nucleus and the spinal trigeminal tract and nucleus (Fig. 11-14). The inferior olivary complex disappears, and the central tegmental tract, one source of input to the inferior olive, appears about where the cell group was located (Fig. 11-14). Finally, the medial lemniscus begins to shift anterolaterally and to rotate from a posteroanterior orientation, which it exhibits in the medulla, toward a horizontal orientation more characteristic of the pons (Fig. 11-14). At the pons-medulla junction, the cross section of the medial lemniscus is oriented obliquely (posteromedial to anterolateral); by the level of the mid pons, it is horizontal.
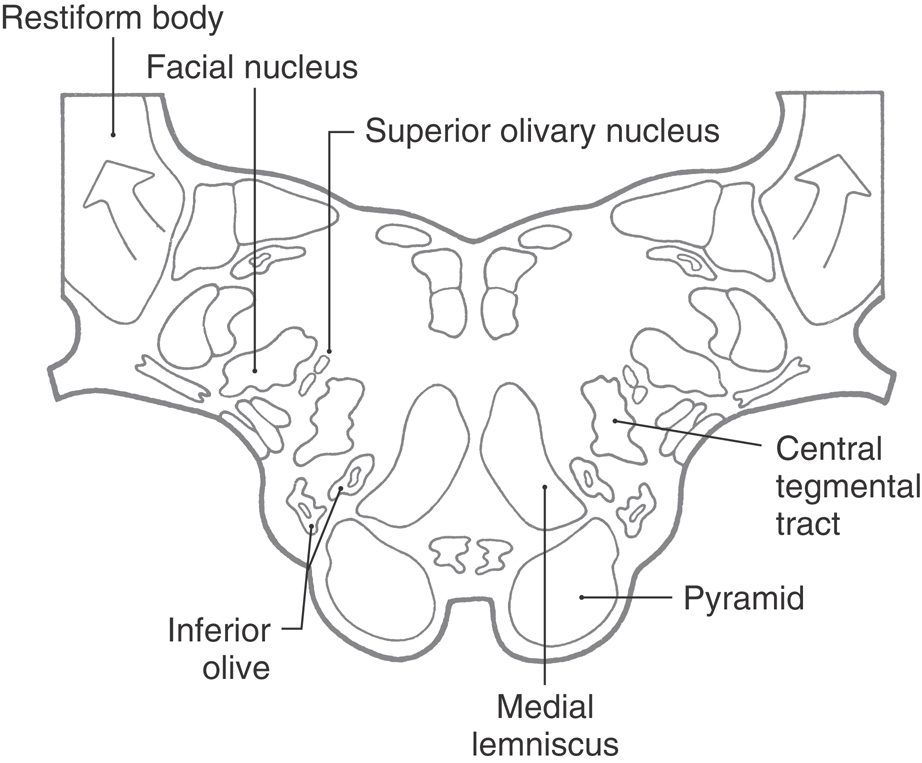
 Figure 11-14.
Figure 11-14. Reticular and Raphe Nuclei
The word reticulum is Latin for “little net” (diminutive of rete, “net”) and denotes mesh-like structures. The reticular nuclei of the brainstem are diffuse and ill-defined and have little apparent internal organization. Collectively, they make up the reticular formation, which may be thought of simply as including all of the cells that are interspersed among the more compact and named structures of the brainstem.
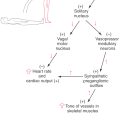
Raphe is a Greek word for “suture” or “seam.” Thus, the raphe nuclei are bilaterally symmetric cell groups in the brainstem that are located directly adjacent to the midline.
The medial medullary reticular area consists of the central nucleus of the medulla at caudal medullary levels and the gigantocellular reticular nucleus rostrally; the latter cell group extends into the pons (Fig. 11-15). The lateral medullary reticular area contains a compact column of cells, the lateral reticular nucleus, and a diffuse population of cells that forms the parvocellular nucleus and the ventrolateral reticular area (area reticularis superficialis ventrolateralis) (Fig. 11-15). The latter cells function in the control of heart rate and respiration. Consequently, a sudden onset of central apnea, indicating damage to these respiratory areas, is often a prime early sign of medullary compression. The raphe nuclei of the medulla are the nucleus raphes pallidus and nucleus raphes obscurus and, at rostral levels, the nucleus raphes magnus (Fig. 11-15). The nucleus pallidus and nucleus obscurus are located at mid to rostral medullary levels along the posterior and anterior midline, respectively. The nucleus raphes magnus begins in the rostral medulla and extends into the caudal pons (Fig. 11-15). In general, the raphe nuclei of the medulla and medulla-pons junction (magnus, pallidus, obscurus) project caudally to spinal cord targets. Cells of these raphe nuclei receive input from several areas, including the central gray of the mesencephalon, and project to the spinal cord. Raphespinal fibers from the raphe magnus are especially important for the inhibition of pain transmission in the posterior horn of the spinal cord. The principal neurotransmitter associated with these nuclei and their fibers is serotonin, although cholecystokinin-containing cells are also found in all three of these raphe nuclei, and enkephalin is found in magnus neurons.
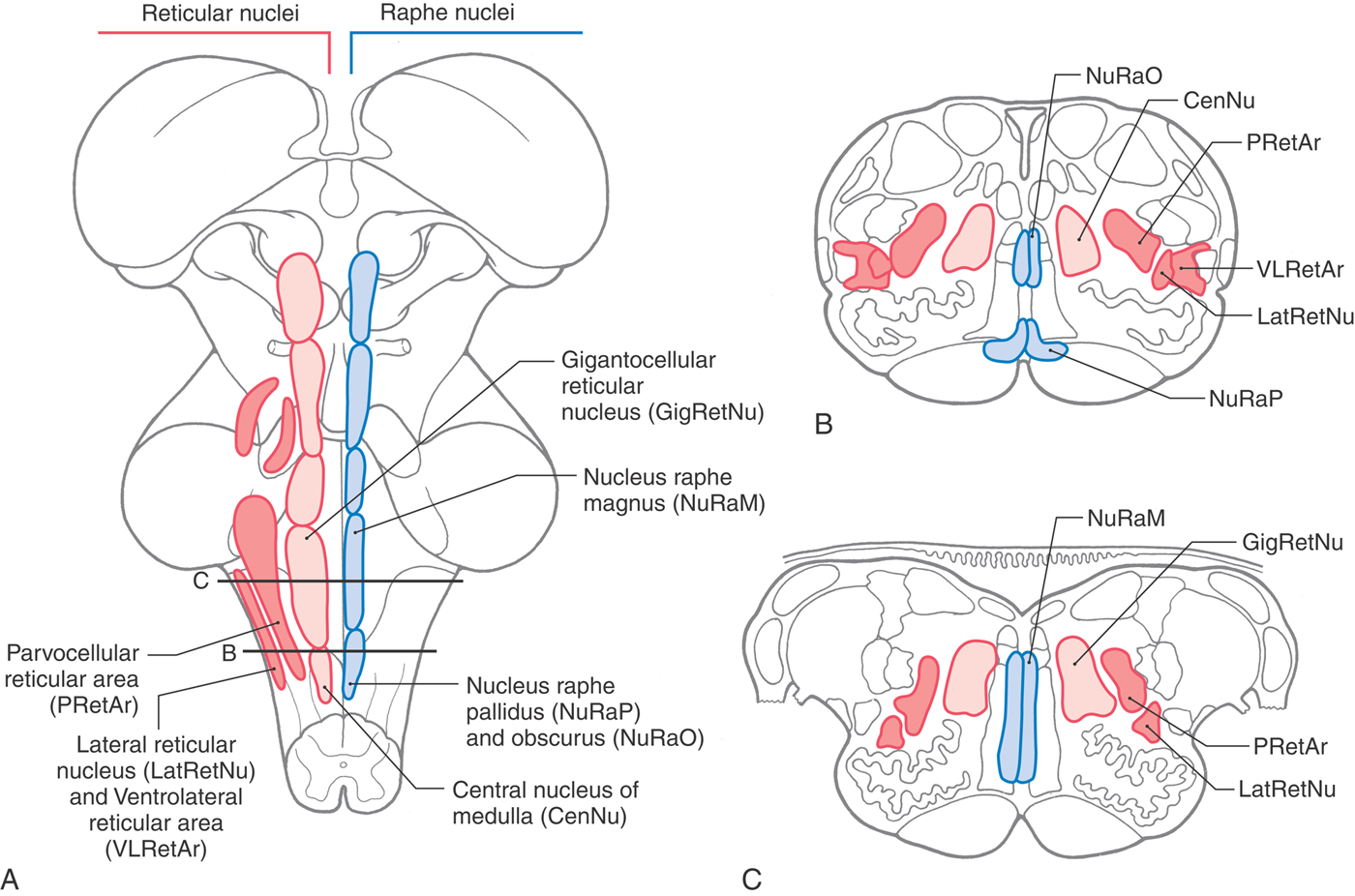
 Figure 11-15.
Figure 11-15. INTERNAL VASCULATURE OF THE MEDULLA AND MEDULLARY SYNDROMES
The blood supply to the medulla arises from branches of the vertebral arteries (Fig. 11-16). These branches are the anterior spinal artery and the PICA. The posterior spinal artery is usually a branch of the PICA.
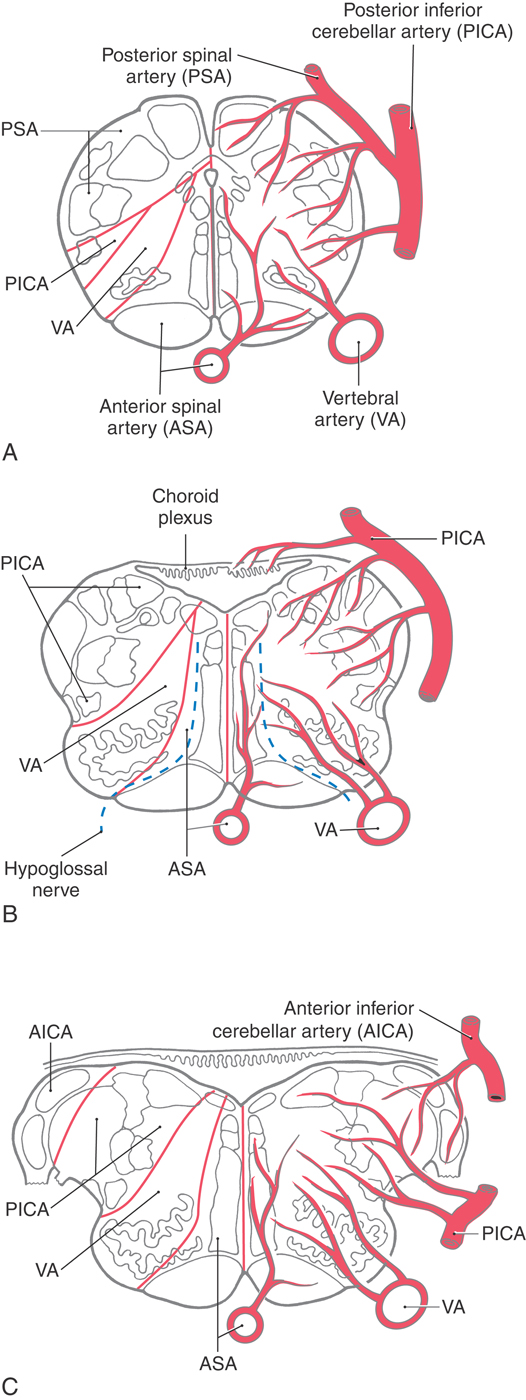
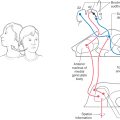
Medial structures of the medulla at all levels, including the pyramid, medial lemniscus, and hypoglossal nucleus and roots, are served by penetrating branches of the anterior spinal artery (Fig. 11-16). The branches of the anterior spinal artery that penetrate into medial portions of the medulla tend to alternate to the right and left. Occlusion of these penetrating branches to one side of the medial medulla may result in a pattern of deficits characteristic of the medial medullary syndrome (Dejerine syndrome). The deficits and corresponding structures damaged in this syndrome include a contralateral hemiparesis (pyramidal and corticospinal damage), a contralateral loss of proprioception and vibratory sense (medial lemniscus), and a deviation of the tongue to the ipsilateral side when it is protruded (hypoglossal root or nucleus injury). On the other hand, occlusion of the anterior spinal artery may result in bilateral deficits reflecting damage to both pyramids, both medial lemnisci, and both of the hypoglossal nuclei or their exiting roots.
The posterior medulla caudal to the obex is served by branches of the posterior spinal artery (Fig. 11-16A). Major structures in this area include the posterior column (gracile and cuneate) nuclei and the spinal trigeminal tract and nucleus. Although vascular lesions of the posterior spinal artery are rare, they may produce an ipsilateral loss of proprioception and vibratory sense on the body (damage to posterior columns and nuclei) coupled with an ipsilateral loss of pain and temperature sensation from the face (spinal trigeminal tract).
Rostral to the obex, the entire posterolateral medulla is served by branches of the PICA (Figs. 11-16B, C and 11-17). Included in the territory served by this vessel are the anterolateral system, spinal trigeminal tract and nucleus, vestibular nuclei, solitary tract and nucleus, and nucleus ambiguus. Vascular insufficiency of the PICA (or blockage of one vertebral artery) gives rise to a characteristic set of sensory and motor deficits commonly called the lateral medullary syndrome, PICA syndrome, or Wallenberg syndrome (Fig. 11-17). The deficits seen and the corresponding structures involved are (1) contralateral loss of pain and temperature sensation from the body (anterolateral system), (2) ipsilateral loss of pain and temperature sensation from the face (spinal trigeminal tract and nucleus), (3) some vertigo and nystagmus (vestibular nuclei), (4) loss of taste from the ipsilateral half of the tongue (solitary tract and nucleus), and (5) hoarseness and dysphagia (nucleus ambiguus or roots of cranial nerves IX and X) (Fig. 11-17C). Patients with the lateral medullary syndrome may also have the Horner syndrome owing to injury to hypothalamospinal fibers descending through the lateral areas of the medulla. We shall further explore the details of these clinical syndromes in later chapters.
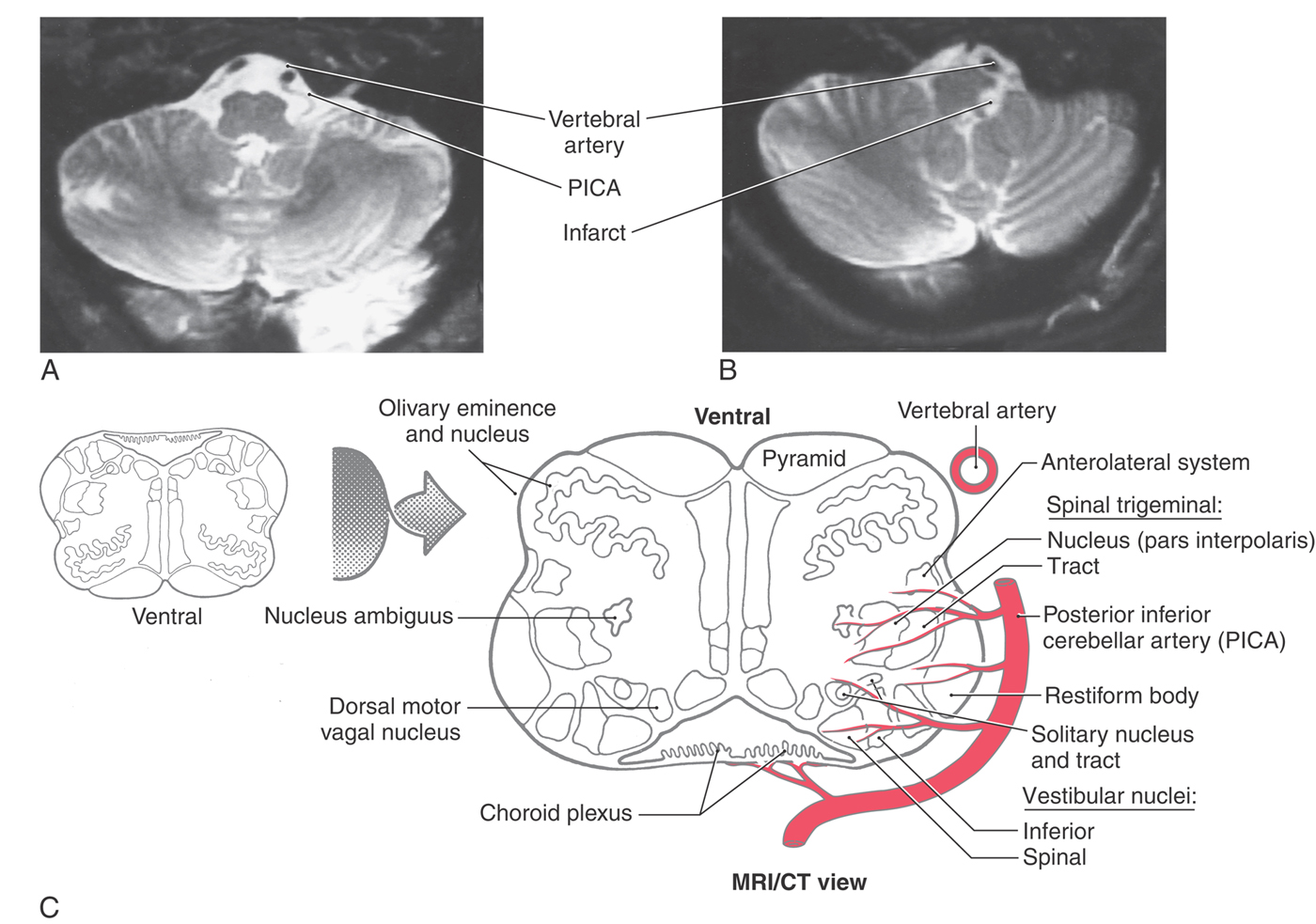
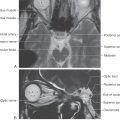
Figure 11-17. Lateral medullary (Wallenberg) syndrome. A normal magnetic resonance image (A) shows the vertebral and posterior inferior cerebellar arteries in relation to the medulla. The patient whose magnetic resonance image is shown in B had an occlusion of the PICA, which resulted in an infarct of the territory of the medulla served by this vessel. The structures damaged in this lesion are shown in C. Compare with Figure 11-16.
In addition to this broad expanse of the medulla, branches of the PICA also serve the choroid plexus of the fourth ventricle. At the pons-medulla junction, the cochlear nuclei and a small adjacent part of the restiform body are served by branches of the anterior inferior cerebellar artery (Fig. 11-16C).
TONSILLAR HERNIATION
Although the tonsil is a portion of the cerebellum, when this structure herniates (Fig. 11-18), it may have a profoundly negative impact on the medulla. The causes of tonsillar herniation vary, but examples include an expanding mass in the posterior fossa (tumor, hemorrhage), lumbar puncture in a patient with a mass lesion in a supratentorial or infratentorial location, and as a complication of surgery in the posterior fossa. In many cases, there is a rapid increase in intracranial pressure, or a shift in pressure, with an extrusion of the cerebellar tonsil downward into and through the foramen magnum. This action impinges on the medulla (Fig. 11-18) and causes damage by two mechanisms. First, the medulla is rapidly compressed and distorted, causing mechanical injury. Second, concomitant with this compression, the blood vessels serving the medulla are occluded, the medulla is deprived of oxygenated blood, and the affected area will become infarcted.
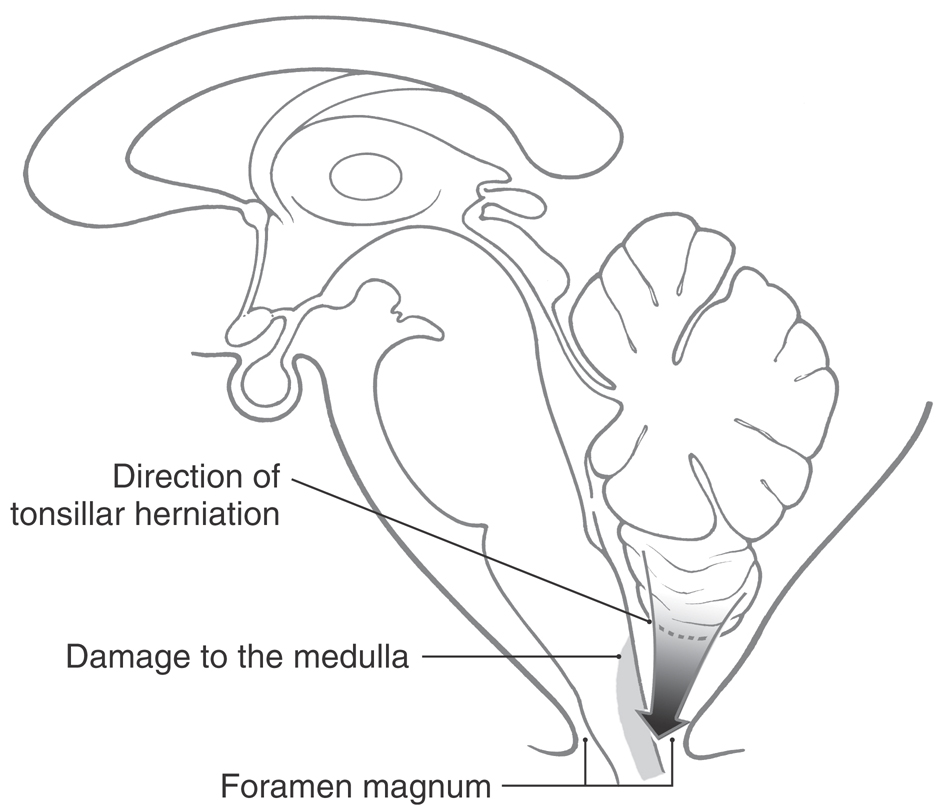
The major concern in acute herniation is damage to the ventrolateral reticular area of the medulla, which contains neurons that influence and control heart rate and respiration. The signs and symptoms include a sudden change in heart rate and respiration (Cheyne-Stokes with intermittent apnea), increase in blood pressure (hypertension), hyperventilation, rapidly decreasing levels of consciousness, and death. In some (chronic) cases the herniation may proceed slowly, and although the medulla is deformed, the patient has minimal neurologic consequences.
Sources and Additional Reading
Readings for the brainstem chapters are listed at the end of Chapter 13.