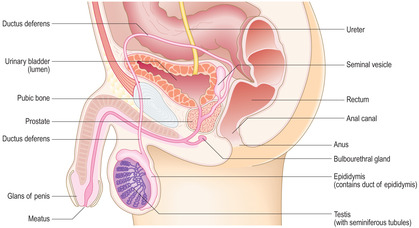
[level-membership-for-basic-science-category]The male reproductive system comprises paired testes, associated glands, ducts and the penis (Fig. 15.1). The testes are the male gonads and are the site of production of spermatozoa (the male gametes). Each spermatozoon (in humans) has 23 chromosomes containing the haploid amount of DNA. Additionally, testes produce and secrete a group of steroid hormones, the male sex hormones (androgens), of which testosterone is the main type. The glands that are associated with the male reproductive system are the paired seminal vesicles, the prostate and paired bulbourethral glands. All these glands contribute fluid secretions which support spermatozoa and form semen. The ducts form the reproductive tract and transport spermatozoa and secretions from the testes and glands to the urethra. The urethra is shared by the reproductive and urinary systems and it passes from the bladder through the prostate and penis and opens at a meatus at the glans penis. The urethra transports urine from the urinary bladder as it is emptied at micturition. The penis is also the organ of copulation able to deliver spermatozoa, in semen, into the vagina of the female reproductive tract.
Testes
Each testis is an organ suspended within a pouch of skin known as the scrotum. The testes thus lie outside the abdominal cavity and this ensures that spermatogenesis (the formation of spermatozoa) occurs at a temperature slightly below body temperature, a requirement for successful gamete production. Each testis is surrounded by a tough connective tissue covering known as the tunica albuginea. Connective tissue septa from the tunica albuginea penetrate into the substance of each testis and divide it into lobules. One septum forms a wedge which penetrates the posterior surface of each testis.
Seminiferous tubules
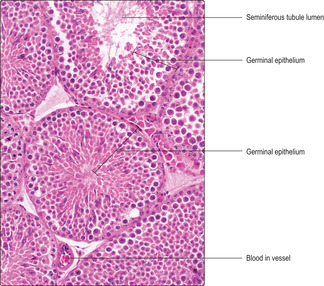
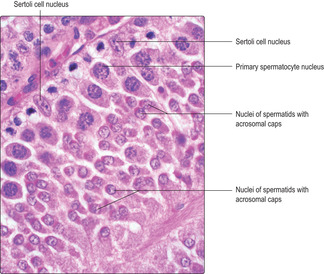
Located within the lobules of each testis are coiled seminiferous tubules which are the sites of spermatogenesis. Seminiferous tubules are lined by several layers of epithelial cells forming a germinal (seminiferous) epithelium (
Fig. 15.2). There are two types of cell in the germinal epithelium of adults: spermatogenic cells, which are in layers, and Sertoli cells, which form a single layer and are supportive of the spermatogenic cells (see below). The germinal epithelium lies on a basement membrane and is surrounded by a thin layer of loose connective tissue. This connective tissue supports blood vessels (
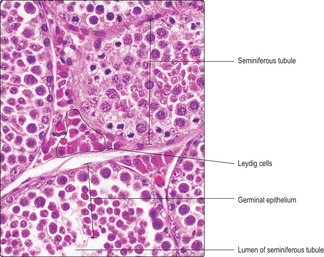
Fig. 15.2), lymph capillaries, nerves and Leydig (interstitial) cells (
Fig. 15.3) which secrete androgens (see below). Myoid cells (
Fig. 15.4) are closely adjacent to seminiferous tubules and are capable of contraction, thus helping to move the spermatozoa along tubules and away from where they develop.
Spermatogenic cells
The formation of spermatozoa (spermatogenesis) may be divided into three stages: mitosis, meiosis and spermiogenesis. These stages begin at about puberty and continue throughout life.
■ Mitosis. In this stage, stem cells (spermatogonia) at the base of the germinal epithelium undergo regular cycles of mitosis and produce new diploid cells, i.e. cells containing the normal number of chromosomes (46 in humans, comprising 22 homologous pairs and two sex chromosomes). Some of the new cells formed by mitosis remain in the basal layer as spermatogonia, others move towards the lumen and enter meiosis.
■
Meiosis. At the start of this phase the cells begin to enlarge and are known as primary spermatocytes. They undergo an S phase when they replicate the DNA in their chromosomes prior to cell division (
Chapter 2). They then enter the prophase of the first division in meiosis (
Fig. 15.6), a process which lasts about 20
days. During this time, chromosomes begin to coil and condense (and stain densely) and homologous pairs of chromosomes move close together and exchange lengths of DNA. This exchange introduces genetic variation to the gametes produced.
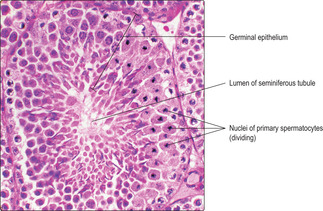
The first meiotic division of meiosis (also known as the reduction division) continues into metaphase. Each pair of homologous chromosomes in each primary spermatocyte line up together on the same part of the equator of the spindle of the metaphase plate, prior to cell division. (This contrasts with mitosis, where each chromosome attaches to a different part of the equator of the spindle.) At this stage, the chromosomes are at their most condensed and densely stained and are readily identified in histological sections (
Fig. 15.7). The paired chromosomes then move away from each other in anaphase and telophase. The result of this reduction division is the production of two secondary spermatocytes each with only half the original number of chromosomes (i.e. 23 in humans). (This reduction division introduces further genetic variation to the new cells as maternally and paternally derived chromosomes in the homologous pairs are randomly arranged on the spindle, and therefore randomly distributed between the two offspring cells.)
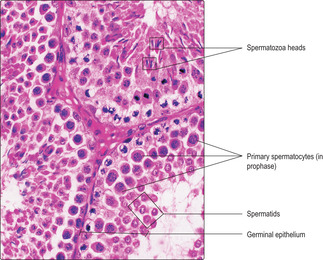
Separation of cytoplasm during meiosis is incomplete and spermatids derived from one spermatogonium are partially joined together. This is thought to aid synchrony of their progression through the next stage (spermiogenesis). Groups of newly formed, related spermatids appear as clusters of small, spherical cells near the luminal surface of seminiferous tubules (
Fig. 15.6).
■
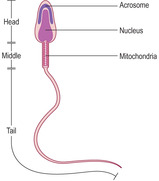
Spermiogenesis. During this stage, spermatids differentiate and each forms one spermatozoon. Approximately 200 million spermatozoa are produced daily by the testes in humans. During spermiogenesis the spherical spermatids change in form and shed cytoplasm as they become highly differentiated spermatozoa characterised by a head, middle region and tail (
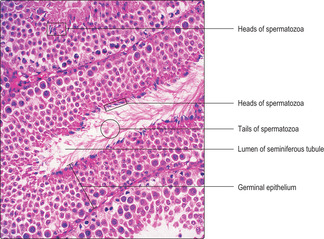
Fig. 15.8). As spermatozoa develop, the head regions remain in the epithelium but the tail regions extend into the lumen of the seminiferous tubule (
Fig. 15.5). Eventually, spermatozoa pass into the lumen and fluid secreted by Sertoli cells (see below) aids their transport away from the seminiferous tubules.
The head of each spermatozoon contains the nucleus in which the DNA in the chromosomes is condensed and stains densely with bases such as haematoxylin (
Figs 15.4,
15.5 and
15.6). At one end of the nucleus of a developing spermatozoon a head ‘cap’ develops known as the acrosome (
Fig. 15.8). The contents of the acrosome include hydrolytic enzymes that take part in the acrosome reaction, which is the process by which the head of a spermatozoon penetrates the cells and structures surrounding the oocyte prior to fertilisation (
Chapter 16). The middle region of a spermatozoon is mainly occupied by mitochondria which provide energy for the movement of the tail (flagellum) (
Fig. 15.8). The tail has the 9 + 2 arrangement of microtubules characteristic of cilia (
Chapter 2), and these provide the motive force which allows spermatozoa to swim. However, spermatozoa become self-propelled only after they have left the testes and are fully matured, when they are then able to undertake unidirectional travel at about 3
mm/minute.
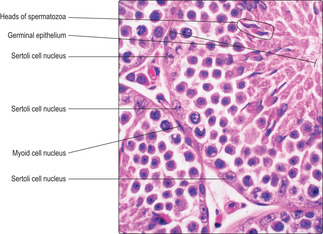
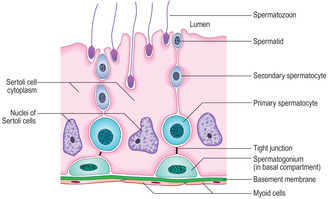
Sertoli cells
Sertoli cells are tall and columnar, and extend from the base of the germinal epithelium to the lumen of the seminiferous tubule (
Fig. 15.10). The nucleus of Sertoli cells may appear oval or triangular and most display a prominent
nucleolus and palely stained chromatin, reflecting their high levels of synthetic activity (Figs 15.4 and
15.9). Developing spermatogenic cells are enfolded in the lateral cell membranes of Sertoli cells and as a result detail of Sertoli cell structure is difficult to distinguish in routine histological sections. Adjacent Sertoli cells are joined to each other near to the basement membrane by tight junctions (
Fig. 15.10). This arrangement separates spermatogonia into a basal compartment of the germinal epithelium and the developing spermatogenic cells (from primary spermatocytes to spermatozoa) into an adluminal compartment. The barrier formed between the two compartments is known as the blood–testis barrier and it ensures that large molecules, e.g. antibodies or microorganisms in blood, do not readily pass to developing spermatozoa.
Variations in germinal epithelium
Various groupings of cells at the same stages of spermatogenesis occur in different regions of the seminiferous tubules. For example, in some regions (
Fig. 15.5) the condensed heads of spermatozoa before they are released into the lumen are apparent, whereas in other regions none of the developing spermatids has condensed chromatin (
Fig. 15.9). These groupings of similar cells developing in synchrony occur as lengths (
Fig. 15.5) or as patches of the epithelium (
Fig. 15.7) in humans. This variation ensures that mature spermatozoa are continuously produced and always available for release at ejaculation.
Leydig (interstitial) cells
Leydig cells are endocrine cells in the loose connective tissue around seminiferous tubules (
Fig. 15.3) and they secrete mainly testosterone. The Leydig cells are stimulated to produce and secrete testosterone by luteinising hormone secreted by cells in the anterior pituitary gland (
Chapter 14). Testosterone enters the circulation and stimulates secondary sexual characteristics including skeletal muscle development and the pattern of hair growth. However, circulating levels of testosterone are insufficient to maintain spermatogenesis. Testosterone levels in the testis are kept high enough to stimulate spermatogenesis as the androgen-binding protein secreted by Sertoli cells binds testosterone in the testis. Another hormone from anterior pituitary cells, follicle-stimulating hormone, stimulates Sertoli cells to secrete the androgen-binding protein.
Ducts draining seminiferous tubules
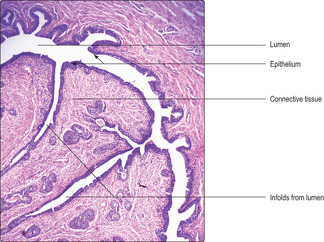
Most seminiferous tubules are coiled loops which have straight regions (tubuli recti) at each end through which fluid and spermatozoa leave. The tubuli recti drain into a meshwork of spaces, lined by cubodial epithelium, known as the rete testis. The rete is embedded in the wedge of connective tissue which extends into each testis from the tunica albuginea. The contents of the rete testis drain into 12 to 15 ductuli efferentes, which in turn drain into a single duct in the epididymis.
Ducts draining each testis (Fig. 15.1)
Ductus epididymis
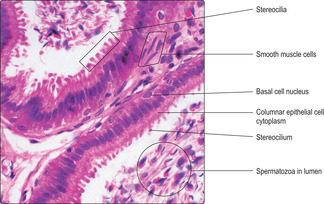
The epithelium lining the ductus epididymis is pseudostratified and composed of basal and columnar epithelial (principal) cells (
Fig. 15.11). The basal cells are stem cells from which the columnar cells are derived. Large cytoplasmic processes, known as stereocilia, extend from the surface of the columnar cells. Stereocilia do not have the structure or function of cilia; instead, they function as microvilli and resorb fluid and remnants of cytoplasm from developing spermatozoa. This fluid resorption and contraction of smooth muscle around the ductus epididymis (
Fig. 15.11) facilitate the movement of spermatozoa away from the testis and towards the ductus deferens.
Ductus deferens
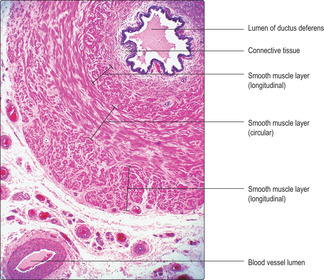
Each ductus deferens is a thick muscular tube that conducts fluid and spermatozoa at ejaculation from a ductus epididymis into an ejaculatory duct in the prostate. At the ejaculatory duct, secretions from a seminal vesicle and the prostate gland are added to the ejaculate and the combined contents (seminal fluid) enter the urethra, where it passes through the prostate.
Each ductus deferens is lined by a pseudostratified columnar epithelium similar to that of the epididymis. There are three thick layers of smooth muscle surrounding the ductus deferens arranged as inner and outer longitudinal layers and a middle circular layer (
Fig. 15.12). Contraction of this relatively large amount of smooth muscle is initiated and coordinated by sympathetic nerves and it ensures that strong contractions propel the seminal fluid along the ductus deferens and on through the urethra at ejaculation.
Glands of the male reproductive system
Prostate
The prostate is a glandular structure that lies beneath the urinary bladder and surrounds part of the urethra. It is a compound tubuloalveolar gland (
Chapter 3) whose epithelium is supported by a fibroelastic stroma which contains numerous irregularly arranged smooth muscle cells (
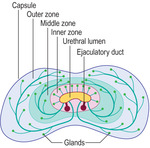
Fig. 15.13). The whole organ is surrounded by a connective tissue capsule. The gland is described in three zones (outer, middle and inner zones) (
Fig. 15.14). The largest zone is the outer zone, and the inner is the smallest. The secretions drain either directly or via short or longer ducts into the urethra.
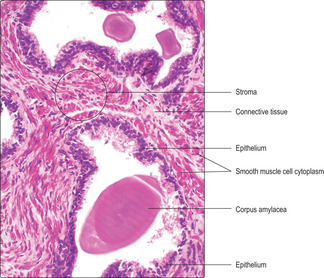
The prostatic epithelium is stimulated to secrete by testosterone. It varies in appearance between being a pseudostratified columnar epithelium (
Fig. 15.13) when stimulated and a low, simple cuboidal epithelium when testosterone levels are low. The lumina of prostatic glands store the secretions which are emptied into the urethra at ejaculation. The propulsive force that
moves the secretions at ejaculation is due to contraction of smooth muscle cells (Fig. 15.13) in the prostatic stroma (under the control of sympathetic nerves). Prostatic secretions contain citric acid, which may provide an energy source for spermatozoa, and various proteins including a form of acid phosphatase unique to the prostate. With age, concretions (corpora amylacea) composed of calcified glycoproteins may be present in the lumina of prostatic glands (
Fig. 15.13) but they are of unknown significance.
Seminal vesicles
Paired seminal vesicles are located on the posterior surface of the urinary bladder. Each contains a highly coiled tube which drains into an ejaculatory duct where, at ejaculation, it is added to fluid from a ductus deferens. (Seminal vesicles in humans do not store spermatozoa.)
Clinical notes
Benign prostatic hypertrophy This condition afflicts 30–40% of men over the age of 50years, and more than 95% of men over the age of 80years. As it is the inner zone of the gland that is affected, any enlargement can readily put pressure on the urethra and affect the ability to pass urine.
Prostate cancer Cancer of the prostate affects about 30% of males over the age of 75years, and in older men it is almost always present. However, in older men it is usually a very slow growing form of cancer and the individuals usually die from some other disease. In cancer of the prostate, it is usually the outer zone that is affected and, in its most aggressive form, it may spread via the bloodstream before any swelling compresses the urethra and inhibits micturition.
A diagnostic test used to indicate an increased risk of prostatic cancer is a blood test to measure the levels of circulating prostate-specific antigen (PSA). This glycoprotein is secreted by prostatic epithelial cells into the lumina of the glands and normally does not circulate in blood. If prostatic epithelial cells have spread beyond their basement membrane their secretions may circulate in blood. However, PSA may also be raised in patients with benign prostatic hypertrophy, so its usefulness as a diagnostic test is limited.
Each tube within a seminal vesicle is lined by a highly folded epithelium which may appear as a pseudostratified columnar epithelium or simple, low cuboidal epithelium. The appearance and function of the epithelium is affected by the amount of circulating male sex hormones. The epithelial-lined tube in each seminal vesicle is supported by fibroelastic connective tissue. Smooth muscle, arranged as an inner circular and outer longitudinal layer, surrounds the coiled tube and on the exterior of the seminal vesicles there is a layer of loose fibroelastic tissue.
Contraction of the smooth muscle in seminal vesicles, stimulated by sympathetic nerves, propels their secretions into the ejaculate. The secretions from the seminal vesicles are watery and contain fructose, which acts as an essential energy source for spermatozoa. They also contain a lipochrome pigment which imparts a yellowish colour to semen.
Bulbourethral glands
The bulbourethral glands are paired glands that lie at the base of the penis and drain directly into the urethra. They secrete a small volume of mucoid material that precedes the ejaculation of semen and acts as a lubricant along the urethra; it may also assist with lubrication of the vagina during sexual intercourse.
Penis
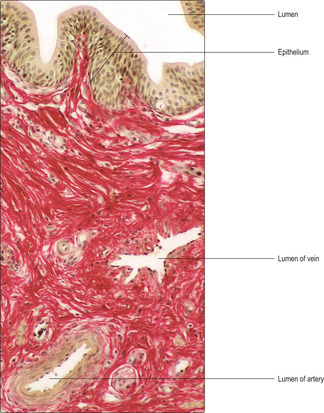
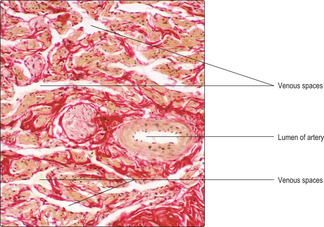
The penis consists of three cylinders (corpora) of erectile tissue, each surrounded by a dense fibrous connective tissue sheath (the tunica albuginea) and enclosed by thin skin. One cylinder, the corpus spongiosum, surrounds the penile urethra (
Fig. 15.15), and the other two (corpora cavernosa) lie dorsally. The erectile tissue of the corpora consists mainly of interconnecting venous spaces (
Fig. 15.17) which are virtually empty when the penis is in its flaccid state.
The penis has a rich blood supply that flows in helicine arteries (
Figs 15.15 and
15.17), which are spiral in form in the flaccid state. Most blood, in the flaccid state, flows directly via arteriovenous shunts linking arterioles (supplied by the helicine arteries) directly to deep venules and veins draining the penis. However, some arterial blood perfuses capillaries and thus supplies the penis with oxygen and nutrients. During an erection, the arteriovenous shunts are closed and arterial blood is diverted and fills and swells the venous spaces (
Fig. 15.17) in all the corpora. As a result, venous return from the penis is restricted, the penis increases in volume, it lengthens, and the helicine arteries extend. The arrangement of the connective tissue around the corpora (
Figs 15.15 and
15.17) ensures that the penis extends as a column. Parasympathetic nerves supplying smooth muscle around arterioles in the penis act as vasodilators and are responsible for the
haemodynamic changes producing an erect penis.Ejaculation is under the control of sympathetic nerves. They stimulate contraction of smooth muscle cells in the ductus deferens (and other ducts), in the prostate gland and in seminal vesicles. The pressure exerted on the urethra by the engorged corpus spongiosum is less than that in the corpora cavernosa. It is normally sufficient to prevent micturition, but not the powerful propulsion of semen at ejaculation. After ejaculation, sympathetic stimulation ceases and smooth muscle contraction ceases. Parasympathetic stimulation also ceases and this opens the arteriovenous shunts and the flaccid state of the penis is restored.
The male reproductive system
■ This consists of paired testes, ducts, associated glands and the penis.
Testes
■ Testes contain coiled seminiferous tubules lined by a stratified germinal epithelium (which produces spermatozoa), Sertoli cells (which support the developing spermatozoa) and Leydig (interstitial) cells (between the tubules) which produce testosterone under the influence of luteinising hormone from the anterior pituitary.
Spermatogenesis
■ This is the series of stages involved in producing spermatozoa: it begins at puberty and continues for life.
■ Spermatogenesis involves mitosis of stem cells (spermatogonia), meiotic division of spermatocytes and spermiogenesis:
■ some cells produced by mitosis of spermatogonia enter meiosis and become primary spermatocytes; others remain as stem cells
■ primary spermatocytes replicate the DNA of their chromosomes prior to meiotic division
■ in the first division (reduction) of meiosis, a primary spermatocyte divides into two secondary spermatocytes, each with half the original number of chromosomes
■ in the second division (mitotic) of meiosis, each secondary spermatocyte divides into two spermatids, each with half the amount of DNA (and half the original number of chromosomes)
■ spermatids differentiate into spermatozoa (the process of spermiogenesis):
■ spermatozoa each have a head, neck and tail (the tail beats and provides unidirectional propulsion).
Ducts
■ Ducts drain spermatozoa and fluid secreted by Sertoli cells from seminiferous tubules.
■ Immotile spermatozoa pass from each testis into a ductus epididymis as fluid is absorbed by epithelial cells lining these ducts.
■ At ejaculation, spermatozoa pass rapidly from each ductus epididymis into and along a ductus deferens, propelled by the peristaltic contraction of the thick muscle layers surrounding each ductus deferens.
Associated glands
■ These add secretions to the ejaculate which provide nutrients for spermatozoa:
■ the prostate gland has smooth muscle in the stroma which contracts and expels the secretions from the epithelial cells
■ the paired seminal vesicles each contain a coiled tubule lined with epithelial cells and surrounded by smooth muscle.
Penis
■ The penis comprises three corpora of erectile tissue, each surrounded by a dense fibrous connective tissue sheath and enclosed by thin skin. One cylinder, the corpus spongiosus, surrounds the penile urethra.
■ increased blood flow into the blood vessels in erectile tissue, particularly in the paired corpora cavernosus, produces an erect penis.
■ At ejaculation, fluid from Sertoli cells, spermatozoa and secretions from the prostate and seminal vesicles enter the prostatic region of the urethra, pass along the penile urethra and are discharged at the meatus of the urethra.
[/level-membership-for-basic-science-category][not-level-membership-for-basic-science-category]The male reproductive system comprises paired testes, associated glands, ducts and the penis (Fig. 15.1). The testes are the male gonads and are the site of production of spermatozoa (the male gametes). Each spermatozoon (in humans) has 23 chromosomes containing the haploid amount of DNA. Additionally, testes produce and secrete a group of steroid hormones, the male sex hormones (androgens), of which testosterone is the main type. The glands that are associated with the male reproductive system are the paired seminal vesicles, the prostate and paired bulbourethral glands. All these glands contribute fluid secretions which support spermatozoa and form semen. The ducts form the reproductive tract and transport spermatozoa and secretions from the testes and glands to the urethra. The urethra is shared by the reproductive and urinary systems and it passes from the bladder through the prostate and penis and opens at a meatus at the glans penis. The urethra transports urine from the urinary bladder as it is emptied at micturition. The penis is also the organ of copulation able to deliver spermatozoa, in semen, into the vagina of the female reproductive tract.
Testes
Each testis is an organ suspended within a pouch of skin known as the scrotum. The testes thus lie outside the abdominal cavity and this ensures that spermatogenesis (the formation of spermatozoa) occurs at a temperature slightly below body temperature, a requirement for successful gamete production. Each testis is surrounded by a tough connective tissue covering known as the tunica albuginea. Connective tissue septa from the tunica albuginea penetrate into the substance of each testis and divide it into lobules. One septum forms a wedge which penetrates the posterior surface of each testis.
Seminiferous tubules
Located within the lobules of each testis are coiled seminiferous tubules which are the sites of spermatogenesis. Seminiferous tubules are lined by several layers of epithelial cells forming a germinal (seminiferous) epithelium (
Fig. 15.2). There are two types of cell in the germinal epithelium of adults: spermatogenic cells, which are in layers, and Sertoli cells, which form a single layer and are supportive of the spermatogenic cells (see below). The germinal epithelium lies on a basement membrane and is surrounded by a thin layer of loose connective tissue. This connective tissue supports blood vessels (
Fig. 15.2), lymph capillaries, nerves and Leydig (interstitial) cells (
Fig. 15.3) which secrete androgens (see below). Myoid cells (
Fig. 15.4) are closely adjacent to seminiferous tubules and are capable of contraction, thus helping to move the spermatozoa along tubules and away from where they develop.
Spermatogenic cells
The formation of spermatozoa (spermatogenesis) may be divided into three stages: mitosis, meiosis and spermiogenesis. These stages begin at about puberty and continue throughout life.
■ Mitosis. In this stage, stem cells (spermatogonia) at the base of the germinal epithelium undergo regular cycles of mitosis and produce new diploid cells, i.e. cells containing the normal number of chromosomes (46 in humans, comprising 22 homologous pairs and two sex chromosomes). Some of the new cells formed by mitosis remain in the basal layer as spermatogonia, others move towards the lumen and enter meiosis.
■
Meiosis. At the start of this phase the cells begin to enlarge and are known as primary spermatocytes. They undergo an S phase when they replicate the DNA in their chromosomes prior to cell division (
Chapter 2). They then enter the prophase of the first division in meiosis (
Fig. 15.6), a process which lasts about 20
days. During this time, chromosomes begin to coil and condense (and stain densely) and homologous pairs of chromosomes move close together and exchange lengths of DNA. This exchange introduces genetic variation to the gametes produced.
The first meiotic division of meiosis (also known as the reduction division) continues into metaphase. Each pair of homologous chromosomes in each primary spermatocyte line up together on the same part of the equator of the spindle of the metaphase plate, prior to cell division. (This contrasts with mitosis, where each chromosome attaches to a different part of the equator of the spindle.) At this stage, the chromosomes are at their most condensed and densely stained and are readily identified in histological sections (
Fig. 15.7). The paired chromosomes then move away from each other in anaphase and telophase. The result of this reduction division is the production of two secondary spermatocytes each with only half the original number of chromosomes (i.e. 23 in humans). (This reduction division introduces further genetic variation to the new cells as maternally and paternally derived chromosomes in the homologous pairs are randomly arranged on the spindle, and therefore randomly distributed between the two offspring cells.)
Separation of cytoplasm during meiosis is incomplete and spermatids derived from one spermatogonium are partially joined together. This is thought to aid synchrony of their progression through the next stage (spermiogenesis). Groups of newly formed, related spermatids appear as clusters of small, spherical cells near the luminal surface of seminiferous tubules (
Fig. 15.6).
■
Spermiogenesis. During this stage, spermatids differentiate and each forms one spermatozoon. Approximately 200 million spermatozoa are produced daily by the testes in humans. During spermiogenesis the spherical spermatids change in form and shed cytoplasm as they become highly differentiated spermatozoa characterised by a head, middle region and tail (
Fig. 15.8). As spermatozoa develop, the head regions remain in the epithelium but the tail regions extend into the lumen of the seminiferous tubule (
Fig. 15.5). Eventually, spermatozoa pass into the lumen and fluid secreted by Sertoli cells (see below) aids their transport away from the seminiferous tubules.
The head of each spermatozoon contains the nucleus in which the DNA in the chromosomes is condensed and stains densely with bases such as haematoxylin (
Figs 15.4,
15.5 and
15.6). At one end of the nucleus of a developing spermatozoon a head ‘cap’ develops known as the acrosome (
Fig. 15.8). The contents of the acrosome include hydrolytic enzymes that take part in the acrosome reaction, which is the process by which the head of a spermatozoon penetrates the cells and structures surrounding the oocyte prior to fertilisation (
Chapter 16
Buy Membership for Basic Science Category to continue reading.
Learn more here
[/not-level-membership-for-basic-science-category]
Histology An Illustrated Colour Text