The endocrine system comprises cells which synthesise particular molecules and secrete them into blood vessels. This contrasts with exocrine secretions in which secreted molecules pass along a duct system to their site of action, e.g. from salivary glands to the mouth. The particular molecules secreted by endocrine cells are known as hormones and the vascular circulation carries them around the body where they interact with various cells described as target cells. The principal means by which hormones achieve their specific action is by interacting with receptor molecules expressed by the target cells in various tissues and organs. Once stimulated, target cells respond in a variety of ways, e.g. by increasing synthesis of certain molecules. Hormones may be steroids, peptides or proteins, or other molecules derived from amino acids. In general, hormones are involved in regulating metabolic activities in cells in many organs and tissues of the body, many of which are important in controlling homeostasis.
The endocrine organs comprise the pituitary gland, thyroid gland, parathyroid glands, adrenal (suprarenal) glands and the pineal gland. In addition, the pancreas contains clusters of endocrine cells known as islets of Langerhans amongst the pancreatic exocrine cells (
Chapter 12), the gonads contain cells secreting reproductive hormones (
Chapters 15 and
16) and secretions from endocrine cells in the hypothalamus affect the secretory activities of the pituitary gland (see below). There are also small groups of neuroendocrine cells in many other regions, e.g. in the epithelial linings of the gastrointestinal tract. Some are known as paracrine cells and their secretions act in surrounding areas.
All endocrine glands and endocrine cells are well supplied by blood providing the metabolites needed to synthesise hormones. The capillaries in endocrine glands are fenestrated and most hormones are secreted directly into them. A fine meshwork of reticulin fibres supports most endocrine cells and capillaries in endocrine glands. This arrangement facilitates the movement of molecules from and to the capillaries.
Hypothalamus and pituitary gland (hypophysis)
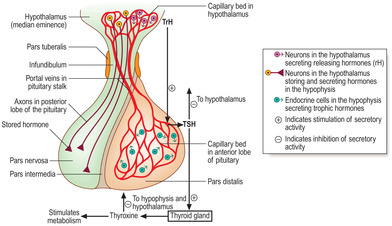
The hypothalamus and pituitary gland function together (
Fig. 14.1). The hypothalamus is the part of the brain connected by a stalk to the pituitary gland. The hormones produced by the hypothalamus and the pituitary affect the function of many cells in different parts of the body.
The pituitary gland is situated in the cranial cavity in a depression in the sphenoid bone of the skull known as the pituitary fossa. The pituitary has a secretory (glandular) and a neural component called, respectively, the adeno-hypophysis and neurohypophysis. Connective tissue surrounds these two components as a capsule. The components differ in embryological origin, structure and function. The adenohypophysis develops from oral ectoderm and it has clusters or strands of secretory (glandular) cells. The neurohypophysis develops from the part of the brain which forms the hypothalamus. Nerve axons are a prominent structural component of the neurohypophysis and hormones are released from the axons.
The hormones secreted by the pituitary gland are dependent in two distinct ways on hormones produced by cells in the hypothalamus (Fig. 14.1).
■
Adenohypophysis and its vascular connections (
Fig. 14.1). A portal system of capillaries connects the hypothalamus, via the pituitary stalk, with the adenohypophysis. (The definition of a portal system is two capillary beds connected by a set of veins; see hepatic portal system
Chapters 10 and
12.) Some hormones produced by neurons in the hypothalamus are secreted into the first capillary bed in the hypothalamus. These capillaries drain into portal veins which carry the hormones along the pituitary stalk to the second capillary bed in the adenohypophysis. By this route hormones from the hypothalamus interact specifically with endocrine cells in the adenohypophysis and control their secretion of a variety of other hormones.
The hypothalamic hormones which pass in the portal system to the adenohypophysis are peptides and most stimulate specific cells in the adenohypophysis to release their specific hormone. These hypothalamic peptides are known as releasing hormones (rH). For example, the rH from the hypothalamus that stimulates cells in the adenohypophysis to release their hormone which stimulates the thyroid is known as thyroid-releasing hormone (TrH). The pituitary hormone is known as thyroid-stimulating hormone (TSH) and it stimulates the thyroid gland to secrete thyroid hormones, e.g. thyroxine (
Fig. 14.1). (In contrast, a few peptide hormones from the hypothalamus inhibit the release of some hormones from the adenohypophysis.)
■
Neurohypophysis and its neural connections (
Fig. 14.1). Axonal processes of some nerve cell bodies in the hypothalamus extend along the pituitary stalk and into the part of the pituitary gland formed by the neurohypophysis. These neurons synthesise peptide hormones which pass along their axons. These hormones are stored in the end regions of the axons in the neurohypophysis before they are released into the vascular system.
Adenohypophysis
The adenohypophysis consists of three parts, the pars distalis (anterior lobe), the pars intermedia and the pars tuberalis (
Fig. 14.1). The cells of the pars distalis form several specific hormones (see below). In humans, the pars intermedia and pars tuberalis are relatively small regions and their significance is not clear.
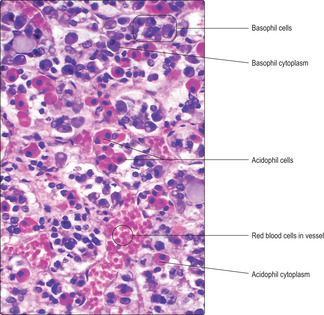
The cells of the pars distalis have been categorised by the reaction of their cytoplasm to various dyes. Cells that bind dyes are termed chromophils and those that do not are categorised as chromophobes. In routine H&E preparations some chromophils bind eosin and are known as acidophils; others bind haematoxylin and are known as basophils (
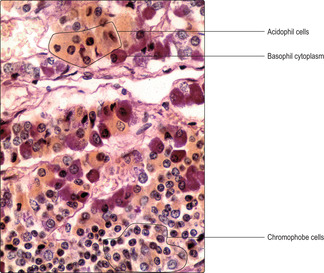
Fig. 14.2). The differences are due to variations in the pH of the hormones stored as cytoplasmic granules in the endocrine cells. The cytoplasm of chromophobes is virtually unstained with most dyes, and in some chromophobes the cytoplasm is sparse (
Fig. 14.3). Other methods of staining cells of the pars distalis emphasise the differences between the chemical composition of their cells by specifically staining the glycoproteins in basophils (
Fig. 14.3). Immunohistochemical methods using light and electron microscopy have now determined precisely which cells in the pars distalis produce the various anterior lobe hormones.
Hormones produced by the adenohypophysis are collectively termed trophic hormones in that they stimulate the release of other hormones from endocrine cells in endocrine glands or in other regions. In general, each endocrine cell in the pars distalis produces only one type of trophic hormone. Some acidophils produce the protein somatotrophin, which is also known as growth hormone (GH) as it is important in stimulating growth, particularly in bones. Other acidophils produce the protein prolactin (PrL), a hormone which has a major effect on milk production by the mammary glands and is also known as mammotrophin. All basophils produce glycoproteins; some produce TSH and others produce adrenocorticotrophic hormone (ACTH), which stimulate, respectively, the thyroid gland and the cells in the cortex of the adrenal glands (see below). Yet other basophils produce two trophic hormones per cell, both categorised as gonadotrophins as they affect endocrine secretion by cells in the gonads (ovaries and testes). The gonadotrophins are follicle-stimulating hormone (FSH) and luteinising hormone (LH). Further details of the function of FSH and LH are described in
Chapters 15 and
16.
Neurohypophysis
The neurohypophysis is described in three parts: the median eminence, the infundibulum and the pars nervosa (posterior lobe of the pituitary) (
Fig. 14.1).
■
Median eminence. This part of the hypothalamus contains:
■ neurons that secrete the hypothalamic releasing hormones which regulate the secretory activity of cells in the pars distalis of the pituitary
■ the primary capillary bed of the portal system which carries these hypothalamic hormones to the pars distalis
■ other hormone-producing neuronal cell bodies which have axons that extend along the infundibulum to the pars nervosa.
■ Infundibulum. This is a continuation of the hypothalamus along the pituitary stalk.
■
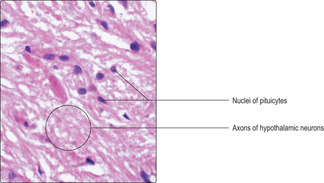
Pars nervosa. This is the region where axons from neurons in the median eminence terminate (
Fig. 14.4). Hormones synthesised by these neuronal cell bodies pass along their axons and are stored in granules in swollen axon terminals, known as Herring bodies, before release. These axons are supported by glial cells, called pituicytes. Two hormones are released from the pars nervosa. They are antidiuretic hormone, which acts on the kidneys, and oxytocin, which acts on mammary glands and the uterus. The effects of these hormones are described in
Chapters 13 and
16.
Thyroid gland
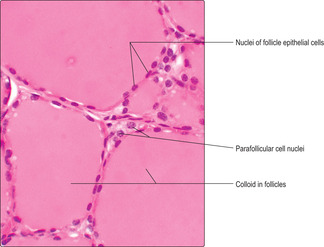
The thyroid gland lies anterior to the lower part of the larynx and upper trachea. It comprises two lobes that are connected across the mid-line by the isthmus. The majority of the endocrine-secreting cells of the thyroid are arranged as simple (single layered) epithelia surrounding roughly spherical spaces filled with colloid (
Fig. 14.5). The colloid is formed by thyroglobulin, an iodinated glycoprotein which is stored as the precursor of active thyroid hormones. These spheres, known as thyroid follicles, vary in size and virtually fill the thyroid gland. Scattered amongst the epithelial cells lining follicles, and in between some follicles, a minor cell population known as parafollicular (C) cells is present (
Fig. 14.5). Parafollicular cells secrete a peptide hormone, calcitonin, which lowers blood calcium levels.
Thyroxine is the major hormone secreted by thyroid epithelial cells. Thyroxine is derived from iodinated tyrosine molecules in thyroglobulin and is also known tetra-iodothyronine (T4). Thyroxine stimulates metabolism, particularly of carbohydrates and lipids, and promotes growth and development.
Thyroxine levels in blood are controlled by mechanisms involving TrH from the hypothalamus and TSH from the anterior lobe of the pituitary gland (
Fig. 14.1). A low level of thyroxine in blood stimulates the secretion of TrH and TSH. The action of TSH on thyroid epithelial cells stimulates the secretion of thyroxine into blood. When the level of thyroxine in blood is too high, negative feedback is activated and thyroxine, acting on the hypothalamus and the hypophysis, suppresses, respectively, the levels of TrH and TSH secreted (
Fig. 14.1). The consequence of this feedback is that secretion of thyroxine decreases and blood levels are lowered.
Parafollicular cells in the thyroid are larger than follicular epithelial cells (
Fig. 14.5). They secrete the peptide hormone calcitonin directly into capillaries. Calcitonin secretion is stimulated by high levels of calcium in blood and its actions help lower blood calcium. Calcitonin, along with parathyroid hormone (see below), maintains calcium homeostasis. Calcium is essential for many body functions, e.g. the conduction of nerve impulses and contraction of muscle. Regulating the amount of calcium in blood is essential for normal nerve and muscle function.
Clinical note
Iodine deficiency Sea water is rich in iodine salts and in some countries supplements of iodine salts are added to table salt. However, in some parts of the world, particularly those distant from a salty sea, there may be insufficient intake of iodine in the diet to provide adequate levels of thyroid hormones. In such cases, TSH levels in blood increase, the thyroid undergoes hypertrophy and a swelling in the neck, known as a goitre, develops.
Parathyroid glands
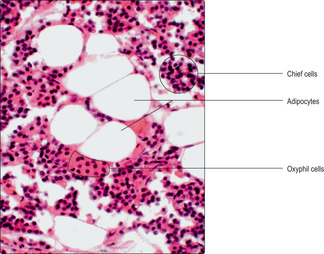
The parathyroid glands are small paired structures on the posterior surface of the thyroid gland. (There is usually more than one pair.) There are two major parenchymal cell types, chief cells and oxyphil cells (
Fig. 14.6). Chief cells are relatively small and make and secrete parathyroid hormone (PTH). Oxyphil cells are larger and stain with dyes such as eosin; their role is not known. In adults, adipocytes appear in the gland (
Fig. 14.6).
PTH assists in maintenance of calcium levels in the blood and in body fluids, in conjunction with calcitonin. Low levels of calcium in blood stimulate the secretion of PTH. Calcitonin and PTH have opposite effects and together affect the mineralisation of bone via the activity of osteoclasts (
Chapter 9).
Adrenal (suprarenal) glands
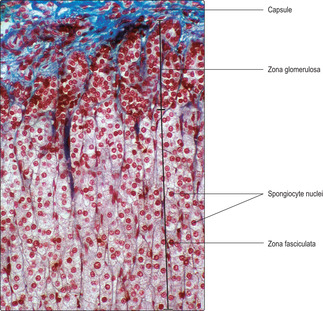
The paired adrenal glands are adjacent to the superior surfaces of the kidneys in humans and are known as suprarenal glands. Each gland is composed of an inner medulla surrounded by a much larger outer cortex. A connective tissue capsule surrounds each gland. The cortex and medulla develop from different embryological origins but both secrete hormones. The cortex is derived from embryonic mesoderm and the medulla from neural crest cells. Steroid hormones are produced and secreted by cortical cells, and vasoactive amines by cells in the medulla.
■
Zona fasciculata. This is the largest region and it occupies the mid-portion of the cortex. It is characterised by rows of endocrine cells (
Figs. 14.7 and
14.8), supported by reticulin fibres, lying alongside straight capillaries extending from capillaries in the zona glomerulosa. The endocrine cells in this zone are known as spongiocytes as their cytoplasm appears vacuolated in routine histological sections (
Figs 14.7 and
14.8). The apparently empty vacuoles contained lipids in life, e.g. cholesterol, stored as a precursor for the synthesis of hormones. The lipids were extracted during histological processing.
■
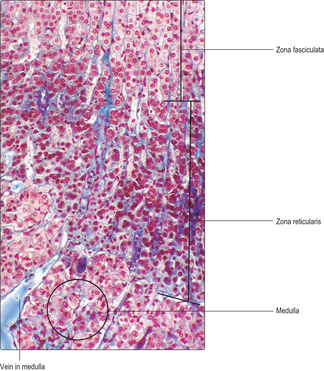
Zona reticularis. This is the inner region of the cortex and it surrounds the medulla. The endocrine cells in this region are relatively small (
Fig. 14.8) and generally the nucleus and cytoplasm are densely stained. These endocrine cells are distributed irregularly on a reticulin meshwork between capillaries which are continuous with those in the zona fasciculata.
Three groups of steroid hormone, known as corticosteroids, are produced from cholesterol by the endocrine cells in the adrenal cortex. The hormones are not stored in the cells but are synthesised when needed. The three groups of steroid hormone are distinguished by their functions.
■
Mineralocorticoids. The molecules in this group regulate water and salt concentration via actions on the distal convoluted tubules of the kidney (
Chapter 13). The major mineralocorticoid hormone is aldosterone and it is produced mainly by cells in the zona glomerulosa.
■ Sex hormones. These steroids are produced mainly by cells in the zona reticularis. Most of the molecules are male sex hormones of low activity.
The secretion of hormones by the adrenal cortex is under the regulatory control of ACTH from the anterior lobe of the pituitary gland. In turn, ACTH secretion is controlled by a releasing hormone (corticotrophin rH) from the hypothalamus. High levels of corticosteroids in blood suppress secretion of corticotrophin rH and ACTH. In addition, mineralocorticoid levels are affected by the intake of various mineral salts.
Adrenal medulla
The medulla of the adrenal gland is surrounded by the zona reticularis of the cortex. Medullary endocrine cells are modified postganglionic sympathetic neurons; they do not have axons. In general, medullary cells are large and in clusters or cords (
Fig. 14.8) near capillaries and are supported by reticulin fibres. The medulla receives blood from arterioles and from capillaries extending in from the zona reticularis. All blood from the cortex drains into veins in the medulla (
Fig. 14.8). There are two principal hormones synthesised by the endocrine cells in the medulla, adrenaline and noradrenaline; both are vasoactive amines. Each hormone is stored in small quantities in cytoplasmic granules. Release of hormones is stimulated by preganglionic nerves acting on the endocrine cells in the medulla. Under stressful conditions, much larger amounts of medullary hormones may be released.
Pancreas
The pancreas is a flattish organ in the abdominal cavity. The bulk of the pancreas is formed by exocrine cells and their secretions enter the gastrointestinal tract at the duodenum via the pancreatic duct (
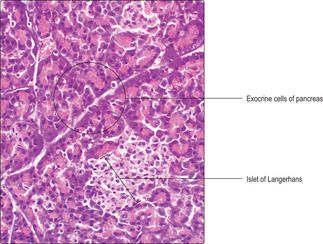
Chapter 12). The pancreas also contains endocrine cells in small clusters known as islets of Langerhans (
Fig. 14.9). There are about a million islets scattered throughout the pancreas.
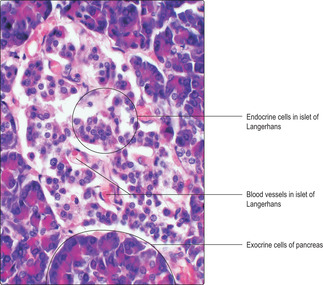
Islets of Langerhans generally are seen in histological sections as pale staining areas in comparison with surrounding exocrine cells (Figs 14.9 and
14.10). The endocrine cells lie close to capillaries (
Fig. 14.10) and several types have been identified. All the hormones produced are involved in regulating carbohydrate metabolism and other aspects of the functions of the digestive system. Immunohistochemical techniques have shown several cell types in each islet, each type secreting a different hormone. Most of the cells secrete insulin and are known as beta cells. Insulin, a polypeptide, reduces blood glucose levels. The next most populous cell type, alpha cells, secretes another polypeptide, glucagon, which elevates blood glucose levels. Other minority cell populations are involved in regulating gastrointestinal tract function.
The factors affecting endocrine secretion by the pancreas are numerous. One of the major factors affecting insulin secretion is the level of glucose in blood. In addition, sympathetic nerves inhibit insulin secretion and, conversely, parasympathetic nerves increase insulin secretion.
Pineal gland
The pineal gland is closely associated with the brain. Despite this proximity, and in contrast to the pituitary gland, there are no nerves which directly connect the pineal gland and the brain. The pineal gland responds to changes in light levels. It is concerned with the regulation of circadian rhythm, and is influenced by hormones from the gonads. The main cell types are pinealocytes and interstitial cells. Pinealocytes produce melatonin in the absence of light. The interstitial cells may be similar to glial cells.
Clinical note
All endocrine glands may secrete abnormally high, or low, levels of hormones. There are numerous causes. Malignant changes in endocrine cells can result in abnormal levels of hormone production. In other instances, abnormal levels of a particular pituitary trophic hormone, e.g. TSH, may be caused by inadequate levels of hormone production by other cells, e.g. inadequate levels of TrH from the hypothalamus. In some cases, an abnormal (auto)antibody may develop which binds to a particular circulating hormone and affects the function of that hormone. Some autoantibodies affect the thyroid gland and result in hypothyroidism manifest as Hashimoto’s disease.
The endocrine system
Hypothalamus and hypophysis (pituitary gland)
■ The hypothalamus is the part of the brain connected by a (pituitary) stalk to the hypophysis.
■ The hypophysis has a glandular portion (adenohypophysis) and neural portion (neurohypophysis).
■ Some hypothalamic hormones (peptides) affect the function of the adenohypophysis:
■ neurons in the hypothalamus secrete releasing hormones into capillaries in the hypothalamus which pass (in a portal circulation) along the stalk and stimulate the release of hormones from cells in the adenohypophysis
■ Other neurons in the hypothalamus produce hormones (peptides) which pass via their axons (along the stalk) to the neurohypophysis.
■ Feedback signals from the levels of hormones in blood affect the secretion of most hypothalamic and hypophyseal hormones.
Adenohypophysis
■ Chromophil and chromophobe cells are present:
■ acidophil cells secrete growth hormone or prolactin (proteins)
■ basophil cells secrete thyroid-stimulating hormone or adrenocorticotrophic hormone or follicle-stimulating hormone and luteinising hormone (glycoproteins).
■ Chromophobe function is not understood
Neurohypophysis
■ Hypothalamic hormones antidiuretic hormone (ADH) and oxytocin are stored in the ends of the axons (as Herring bodies):
■ ADH affects kidney tubules and decreases the amount of water excreted
■ oxytocin stimulates lactation and contraction of uterine smooth muscle.
Thyroid
■ A simple epithelium synthesises, secretes and stores thyroglobulin (an iodinated protein) as colloid in spherical follicles.
■ These epithelial cells also endocytose the thyroglobulin, convert it into active thyroid hormones (e.g. thyroxine) and secrete them into blood vessels:
■ thyroid hormones stimulate metabolism.
■ Parafollicular cells secrete the peptide hormone calcitonin directly into blood when blood calcium levels are high. Calcitonin lowers calcium levels in blood.
Parathyroid glands
■ Chief cells secrete parathyroid hormone when blood calcium is low, which raises calcium levels via its stimulation of the activity of osteoclasts. Oxyphil and adipose cells are also present.
Adrenal glands
■ Adrenal cortex synthesises, secretes steroids:
■ zona glomerulosa cells secrete mineralocorticoids which affect salt balance
■ zona fasciculata cells (spongiocytes) secrete glucocorticoids which regulate metabolism and are immunosuppressive
■ zona reticularis cells secrete, male sex hormones of low activity.
■ Adrenal medulla secretes vasoactive amines:
■ cells are modified sympathetic neurons which secrete adrenaline or noradrenaline involved in responses (fright, fight and flight) to stress.
Pancreas
■ Islets of Langerhans cells secrete hormones (peptides) which affect carbohydrate metabolism. Insulin is secreted by beta cells and it reduces blood glucose levels. Glucagon is secreted by alpha cells and it raises blood glucose levels.
Pineal gland
■ Responds to light, secretes melatonin and helps regulate circadian rhythm.