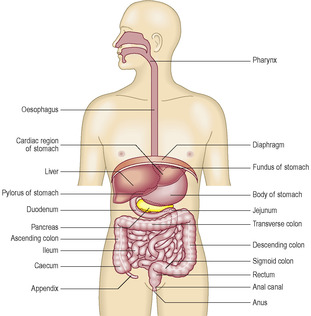
[level-membership-for-basic-science-category]The digestive system consists of the alimentary canal, which is a tube connecting the mouth and anus, and associated structures that facilitate digestion of ingested food and drink (Fig. 12.1). The alimentary canal comprises the mouth, oesophagus and gastrointestinal tract. The gastrointestinal tract comprises the stomach, duodenum, jejunum, ileum, colon, appendix, rectum and the upper part of the anal canal. The associated structures are the teeth, tongue, salivary glands, liver and pancreas. At the mouth, lips intervene between skin and the mouth and they are covered by a stratified squamous epithelium which is not keratinised. At the anus the stratified, keratinised squamous epithelium of skin extends a short distance into the anal canal. The upper part of the anal canal is lined by a stratified, squamous epithelium which is not keratinised.
The digestive system has two primary functions. It breaks down food and drink into small molecules such as glucose, amino acids, fatty acids and triglycerides, a process which involves enzymic activity and is known as digestion. The second primary function of the digestive system ensures the small molecules produced by digestion in the lumen of the alimentary canal enter the body ‘proper’ by being absorbed across the epithelium lining the alimentary canal and into the blood or lymphatic vessels. In addition, the digestive system absorbs water, minerals, vitamins and ions from the material in the lumen of the gastrointestinal tract. The terminal part of the tract functions as a store for components of food that have not been digested. The stored, undigested food and waste products of the body, e.g. from the breakdown of red blood cells, are emptied from the alimentary canal at defecation.
The structure of the alimentary canal is related to its functions and different aspects of function occur at different locations along the canal. The gastrointestinal tract itself is several metres in length in humans, which is a reflection of the space required to accomplish all aspects of its function and is related to the transit times required for these functions.
General structure of the alimentary canal
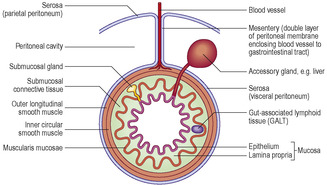
All the alimentary canal is lined by a mucosa (
Chapter 3), which consists of an epithelium, connective tissue lamina propria and, in many regions, a layer of smooth muscle forming the muscularis mucosae (
Fig. 12.2). The epithelium of the mucosa reflects the primary function(s) occurring in that region. However, in all regions the mucosa is important in forming a barrier between substances ingested, including microorganisms, and the internal environment of the body. In regions where the muscularis mucosae is present, contraction of this muscle moves and folds the mucosa, aiding contact between the contents of the lumen and the surface epithelial cells.
A submucosal layer of connective tissue, which supports nerves and blood and lymph vessels, attaches the mucosa in most parts of the alimentary canal to outer layers of muscle (the muscularis externa) (
Fig. 12.2). In most regions of the alimentary canal two layers of muscle are present in the muscularis externa. In the outer, longitudinal layer the long axis of each muscle cell lies roughly parallel to the length of the lumen. In the inner, circular layer the long axes of the muscle cells lie around the lumen. Contractions of the muscularis externa cause peristalsis, which provides the motive force transporting the luminal contents of the canal towards the anus. In the submucosal layer and the muscularis externa, parasympathetic neuronal cell bodies are present and some are involved in co-ordinating muscle activity.
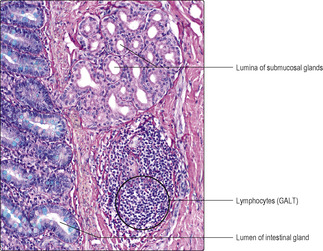
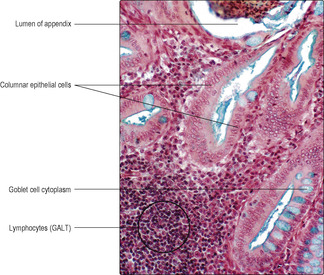
Specialised gut-associated lymphoid tissue (GALT) is present in the walls of the alimentary canal. In some regions, GALT forms large structures, e.g. tonsils; in other regions, only small clusters of lymphoid cells are present (
Fig. 12.2). These cells are part of the primary defence mechanisms against pathogens entering the body across the mucosa lining the gut tube.
Mouth
The mouth (oral cavity) contains the tongue and teeth. The epithelium of the mucosa lining the cheeks and tongue is able to resist the wear and tear in-volved in chewing food, and in most regions it is a stratified, squamous (non-keratinised) epithelium. This epithelial surface is moistened by secretions from small serous and mucous glands in the submucosal layer and from salivary glands (
Fig. 12.3) which drain their saliva into the oral cavity. Physical breakdown of food occurs in the mouth and it is mixed with salivary gland secretions which begin to break down large carbohydrate molecules.
Salivary glands
There are three, paired salivary glands, the parotid, submandibular and sublingual glands. Each gland is invested by a connective tissue capsule, and connective tissue forms trabeculae which penetrate the glands and provide support for the blood vessels and nerves supplying the glands and for the ducts draining them. Each gland has secretory cells arranged as compound, tubuloacinar, exocrine glands (
Chapter 3). The secretory acini drain into small ducts (lined by a simple cuboidal epithelium) which join together in a branching system; the larger ducts are lined by columnar epithelial cells, which may be stratified. The ducts drain the saliva into the oral cavity.
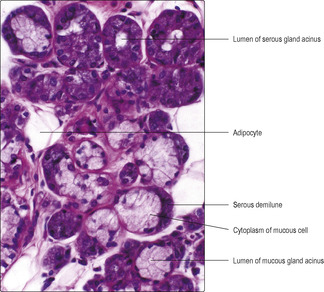
Two types of secretory cell, serous and mucous cells, are present in salivary gland acini (
Fig. 12.3) and, respectively, they produce a watery secretion containing proteins which function as enzymes and a viscous mucus in which large carbohydrate complexes are major components. In addition, myoepithelial cells wrap around the acini and their contractions help to expel the secretions. The secretions assist the process of digestion by moistening and lubricating the food, and by providing enzymes. The main enzyme is amylase, which breaks down large carbohydrate molecules. Saliva is also responsible for lubricating and cleansing the oral cavity. It contains bactericidal substances such as lysozyme secreted by serous cells and may contain immunoglobulin A secreted by plasma cells.
Submandibular glands
A submandibular gland lies beneath the mandible on each side, wrapped around the muscle which supports the tongue. Each gland drains via a duct that opens on the ipsilateral side of the ridge (the frenulum) on the undersurface of the tongue. Mucous glands are a prominent feature of submandibular glands, but serous cells are also present. The arrangement of the serous and mucous gland cells together in some acini is such that these are described as mucous glands with serous demilunes (
Fig. 12.3).
Sublingual glands
The sublingual glands are the smallest of the salivary glands. Mucous gland cells are predominant in sublingual glands and the ducts open directly into the floor of the oral cavity, rather than via a single duct system.
Tongue
The tongue is covered by a stratified, non-keratinised, squamous epithelium which on the undersurface is similar to the epithelium lining the mouth. The upper surface of the tongue is divided by a ‘V’-shaped groove into an anterior (two-thirds) and a posterior region developed from different parts of the embryo (see Mitchell B, Sharma R.
Embryology: An Illustrated Colour Text. Elsevier: 2004). The stratified squamous epithelium on the upper surface is studded by prominent projections. In the posterior region, many of the projections are due to aggregations of lymphocytes deep to the epithelium. In addition, three distinct types of projection known as papillae are also described, and all assist in macerating food and resisting abrasion:
■ Filiform papillae. These are numerous on the upper surface and are in parallel rows which converge towards the midline. The surface of these papillae is covered by a keratinised, stratified squamous epithelium.
■ Fungiform papillae. These are covered by a stratified (non-keratinised) squamous epithelium. Specialised clusters of sensory cells (taste buds) are present in the epithelium covering this type of papilla.
■ Circumvallate papillae. There are 8 to 12 large, circumvallate papillae visible to the unaided eye just anterior to the ‘V’-shaped groove of the tongue. They are also covered by a stratified (non-keratinised) squamous epithelium which contains taste buds.
Deep to the mucosa of the tongue there are numerous, small serous and mucous glands which secrete onto the surface of the epithelium. The major component of the inner part of the tongue is skeletal muscle (see
Fig. 5.7). The bundles of muscle cells are arranged in a complex three-dimensional meshwork and coordinated contraction of the muscle cells is important in chewing, swallowing and in sound production.
Pharynx and oesophagus
Swallowed food and drink pass via the pharynx to the oesophagus (gullet). The structure and function of the epithelium lining the component parts of the pharynx are described in
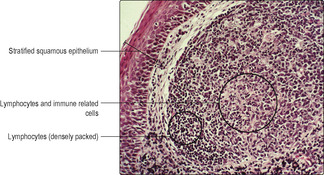
Chapter 11. Aggregations of lymphoid cells are a prominent feature of the walls of the pharynx and form tonsils (
Fig. 12.5). The lymphoid cells are important components of the immune mechanisms defending the alimentary canal (and the respiratory tract).
The major part of the oesophagus is in the thorax, but a small segment lies in the abdominal cavity and is continuous with the stomach. The structures in the walls of the oesophagus conform to the general plan for the gastrointestinal tract (
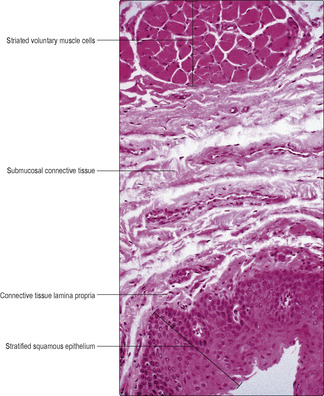
Fig. 12.2). In the thorax, the connective tissue of the adventitia binds the muscularis externa of the oesophagus to adjacent structures. The muscularis externa consists of an outer, approximately longitudinal layer and an inner circular layer. In the upper part of the muscularis externa, striated voluntary muscle (
Fig. 12.6) is present, whereas in the lower region it is replaced by smooth involuntary muscle. In the middle region there is a transition between the two types of muscle. Coordinated contraction of muscle cells in the different regions of the oesophagus ensures that swallowed food and drink normally pass to the stomach. In some regions, the submucosal tissue of the oesophagus contains serous and mucous glands in addition to blood vessels, nerves and aggregations of lymphoid cells. The epithelium of the mucosa lining the oesophagus is stratified and squamous (
Fig. 12.6) and is able to resist friction from swallowed food and chemical attack by swallowed drinks, e.g. alcohol.
The gastrointestinal tract
Stomach
The stomach is covered by a serosal (peritoneal) membrane. The surface squamous epithelial cells of the peritoneal membrane secrete small amounts of fluid (into the peritoneal cavity) and, as the stomach fills with food and drink, it moves relatively easily against adjacent structures. Within the connective tissue of the peritoneal membrane large accumulations of fat cells may be present. The muscularis externa of the stomach differs from the general plan in that it has three layers: an innermost oblique, a middle circular and an outer longitudinal layer. These muscle layers help prevent overdistension of the stomach and their contractions help mix the chyme and move it towards the small intestine. In the pyloric region of the stomach, at the gastroduodenal junction, the muscularis externa is thickened and functions as a sphincter. The sphincter is controlled by autonomic nerves and hormones which regulate the passage of chyme into the duodenum. In contrast, at the gastrooesophageal junction, stomach contents normally are not regurgitated into the oesophagus although a distinct muscular sphincter is not apparent.
The interior of the empty stomach is characterised by thick folds which run longitudinally. They are known as rugae and are formed by submucosal and mucosal layers. The rugae unfold as the stomach fills. They also help to channel chyme towards the pylorus and help to maintain a large interface between the contents of the stomach and the surface epithelium.
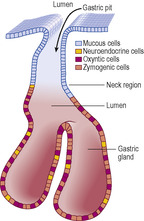
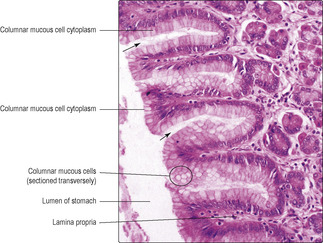
The epithelium of the gastric mucosa dips into the lamina propria and forms channels known as gastric pits (
Fig. 12.7). All the epithelial cells on the surface of all regions of the stomach and lining all the gastric pits are columnar and they secrete mucus (
Fig. 12.8). This mucus forms a thick protective layer which helps to ensure that the stomach cells are not damaged by gastric secretions. At the base of the pits, gastric glands extend as tubes deep into the lamina propria and reach the muscularis mucosae. The gastric glands secrete various substances which drain into the pits, and from there to the lumen of the stomach. In the cardiac and pyloric regions of the stomach gastric glands are coiled tubules lined by columnar cells which secrete mucus. Their appearance in routine H&E-stained sections resembles the mucous cells lining gastric pits. In the rest of the stomach (body and fundus) the gastric glands are straight tubules and the epithelium lining them contains several types of cell:
■
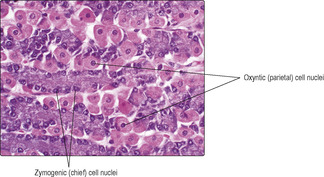
Zymogenic (chief) cells (
Fig. 12.9). These produce and secrete pepsinogen (and lipase, which digests lipids). Pepsinogen is inactive until it is converted by the acidity of the gastic juices, into pepsin, which breaks proteins down into smaller molecules. The release of gastric enzymes is stimulated by the vagus nerve, which is part of the parasympathetic nervous system.
■
Oxyntic (parietal) cells (
Fig. 12.9). These produce and secrete hydrochloric acid, which helps provide the optimum pH for enzymes secreted by zymogenic cells. The acid also destroys some ingested microorganisms. Secretion of acid is stimulated by the vagus nerve too. Oxyntic cells also secrete intrinsic factor, which is necessary for the absorption of vitamin B12, which is essential, in turn, for the production of red blood cells. Without intrinsic factor pernicious anaemia develops.
■ Stem cells. These are located amongst the mucous neck cells and, although relatively few in number, are able to undergo mitosis and give rise to replacement gastric epithelial cells of all types. Some offspring cells differentiate and migrate into their appropriate location within the epithelium; others remain as stem cells.
■ Neuroendocrine cells. These secrete a variety of hormones into the local environment and into blood vessels that modify the activity of other cells. For example, in the mucosa of the stomach some cells secrete gastrin, which stimulates contraction of the muscularis externa of the stomach and relaxation of the pyloric sphincter, thus moving stomach contents into the duodenum.
Small intestine
The small intestine has three parts: duodenum, jejunum and ileum. Digestion continues in the small intestine and involves digestive enzymes from secretory cells in the mucosal epithelium and, in the duodenum, in the submucosa. Secretions also enter the lumen of the duodenum from the pancreas and liver and these also aid digestion. The luminal contents, known as chyle, have a transit time along the small intestine of 3–4 hours and digestion is largely completed in that time. Most of the products of digestion, e.g. amino acids and monosaccharides, are absorbed across the epithelium lining the small intestine and pass into blood vessels; fatty acids and triglycerides pass into lacteals (small, blind-ended lymph vessels).
The wall of the small intestine conforms to the generalized structural plan of the gastrointestinal tract (
Fig. 12.2) though there are features which characterise each part. The epithelium of the mucosa lining the small intestine contains:
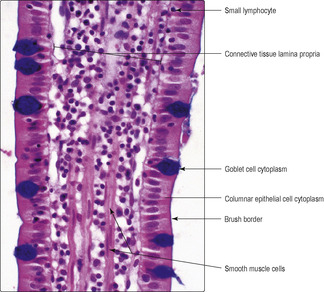
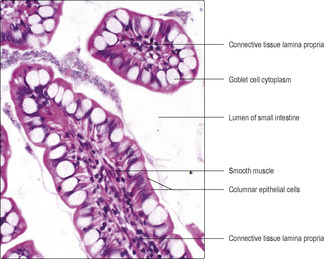
■ columnar epithelial cells (
Fig. 12.10), which are specialised for absorption of small molecules from the lumen (see below)
■ goblet cells, which make and secrete mucus that helps to protect the luminal surface (
Fig. 12.10)
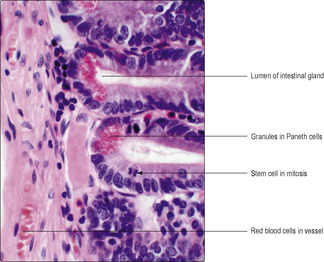
■ Paneth cells, which make, store and secrete lysozyme (
Fig. 12.11)
■ neuroendocrine cells, which secrete molecules that affect the function of other cells
■ lymphocytes, which may be between the epithelial cells (
Fig. 12.10).
Clinical note
Stomach ulcer Some stomach ulcers are the result of damage to epithelial cells lining the stomach by specific molecules, e.g. alcohol or aspirin. If epithelial cells are killed by such molecules the damage may be repaired by the normal proliferative activity of stem cells in the epithelium. However, if the damage is extensive and the production of replacement cells inadequate, the damage may extend into the connective tissue lamina propria and bleeding into the gastric lumen may occur.
There are several structural features of the small intestine that facilitate contact between luminal contents and the mucosal epithelium and thus aid digestion and absorption of the molecules produced by enzymic activities:
■ Transverse folds. These are visible in the duodenum, jejunum and first half of the ileum to the unaided eye. These folds are formed by the mucosa and submucosa and are known as plicae circulares.
■
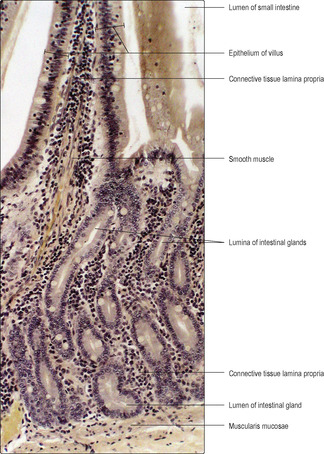
Villi (
Figs 12.12 and
12.13). These are present throughout the small intestine and are microscopic finger-like structures which project from the mucosa into the lumen (
Figs 12.12 and
12.13). They are covered by columnar epithelial absorptive cells and goblet cells (
Fig. 12.13) and have a core of connective tissue supporting blood vessels and lymphatic vessels (lacteals). A few smooth muscle cells are also present in villi (
Figs 12.10,
12.12 and
12.13) and their contractions move the villi amongst the chyle, thus aiding absorption.
■
Intestinal glands (crypts of Lieberkühn). These are tubes which dip into the mucosa as far as the muscularis mucosae (
Fig. 12.12). As well as columnar epithelial and goblet cells, Paneth cells (
Fig. 12.11), neuroendocrine cells and stem cells (
Fig. 12.11) are present in the epithelium of intestinal glands.
The life span of epithelial cells in the mucosa of the small intestine is less than a week; cells at the tip of villi die and are rapidly replaced. It is the mitotic activity (
Fig. 12.11) of stem cells in the epithelium at the base of intestinal glands which ensures that new cells are produced. Some of the new cells differentiate and become columnar epithelial cells, others become goblet cells, yet others remain as stem cells. The new columnar and goblet cells migrate from the intestinal glands along the epithelium and up to the tip of the villus where, in turn, they die.
The first part of the small intestine, the duodenum, is characterised by the presence of glands in the submucosal layer (
Fig. 12.14). These glands (Brunner’s glands) make and secrete alkaline mucus which helps neutralise the acidic chyme entering from the stomach. Secretions from neuroendocrine cells in the duodenum inhibit the action of gastrin and help control the flow of gastric contents to the duodenum.
Large intestine
The large intestine comprises the colon, appendix, rectum and the upper part of the anal canal (
Fig. 12.1). Its structure largely conforms to the general plan (
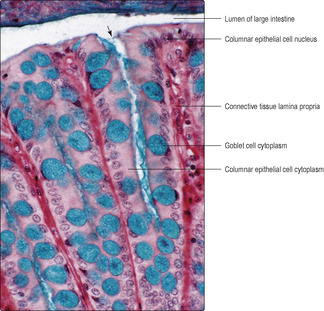
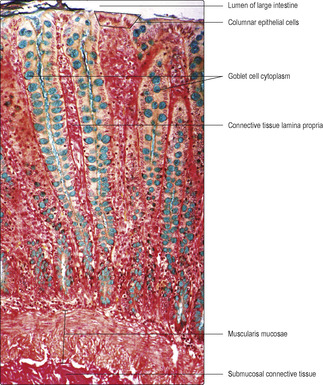
Fig. 12.2). The mucosa is similar along the length of the large intestine, and goblet cells (
Figs 12.15 and
12.16) are far more common than in the small intestine. The mucosal epithelium dips into the mucosa and forms intestinal glands which reach the muscularis mucosae (
Fig. 12.16). This arrangement increases the area of interface between the luminal contents and the epithelial cells. Columnar epithelial cells, absorbing mainly water, are present mainly on or near the luminal surface, and stem cells and a few neuroendocrine cells are also present in the epithelium. Villi and submucosal glands are not present in the large intestine.
The muscularis externa of the large intestine differs from the general plan in that although it has an inner circular layer its outer longitudinal layer is arranged in three bands (taeniae coli). Peristaltic contractions of the muscularis externa move the contents in the lumen towards the anus. Although the large intestine is much shorter than the small intestine, the transit time through the large intestine takes relatively longer at up to about 48 hours. During transit, water and salt absorption occurs and the remnants form feces. The mucus secreted by goblet cells in the epithelium is important in lubricating the movement of feces along the large intestine.
Liver
The liver is a large, highly vascular, glandular structure situated in the upper abdomen. It has significant roles in the function of the digestive system. It is covered by a connective tissue capsule and, in turn, by peritoneum. The cell type which forms the majority of the parenchyma of the liver is the hepatocyte (
Fig. 12.18). The liver receives blood drained from most of the gastrointestinal tract. This brings products of digestion, principally amino acids and mono-saccharides, directly from the gut. Hepatocytes synthesise a range of large molecules, some of which they store, e.g. glycogen; others they secrete into blood, e.g. albumin. Hepatocytes also break down absorbed harmful molecules (e.g. alcohol) and waste molecules (e.g. from the breakdown of haemoglobin). Some waste molecules are secreted by hepatocytes into bile (see below), which drains into the duodenum, from where they are eventually excreted in feces.
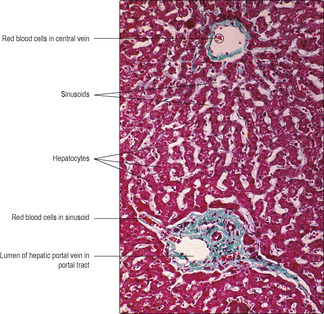
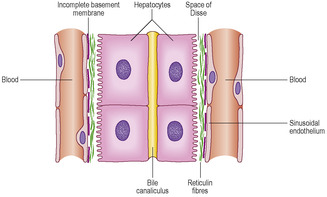
The liver receives blood from two sources: the hepatic artery, which supplies blood that is high in oxygen content, and the hepatic portal vein, which supplies blood that is low in oxygen content. The hepatic portal vein drains blood from the gastrointestinal tract and carries the molecules produced by digestion to the liver. These two sources of blood pass into liver sinusoids (wide vascular channels) lined by irregularly placed endothelial cells lying on an incomplete basement membrane. Non-migratory macrophages, known as Kupffer cells, are interspersed between the endothelial cells and they phagocytose effete red cells (and microorganisms if present in the blood). Columns of hepatocytes lie alongside the sinusoids separated from the basement membrane by a space (the space of Disse) (
Fig. 12.19). Blood passes along the sinusoids and some plasma passes between the endothelial cells into the space of Disse. The hepatocytes adjacent to the space of Disse take up the small molecules produced by digestion and synthesise larger molecules required by the body. Blood drains from sinusoids into small veins, known as central veins (
Fig. 12.18), which join together and leave the liver as the hepatic vein.
Between adjacent hepatocytes there are very small spaces, bile canaliculi (
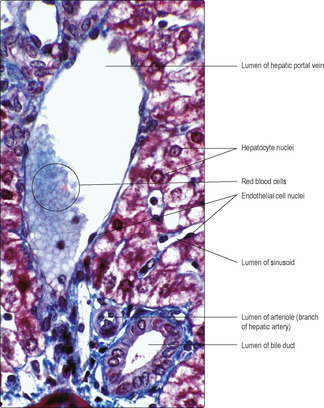
Fig. 12.19). These channels receive bile secreted by hepatocytes. The direction of flow in bile canaliculi is opposite to the blood flow in sinusoids. Bile canaliculi join together and drain into a network of epithelium-lined bile ducts. The bile ducts pass through the liver alongside branches of the hepatic arteries and hepatic portal veins in structures known as portal tracts (
Figs 12.18 and
12.20). The ducts join together and eventually bile drains into a single duct which transports bile, via the gall bladder, to the duodenum. Bile aids the digestion of fats as well as being the route by which the breakdown products from the destruction of red blood cells are excreted.
The classical functional unit of the liver is a hexagonal lobule enclosed by connective tissue. In humans, connective tissue is sparse between lobules but is present in portal tracts supporting branches of the hepatic artery, hepatic portal vein and bile duct (
Figs 12.18 and
12.20) and around central veins (
Fig. 12.18). A fine meshwork of reticulin fibres supports the sinusoids (see
Fig. 4.3). From a functional view, it is appropriate to consider the blood supply as the basis of the functional unit. Hepatocytes closest to where hepatic arteries and hepatic portal veins (in portal tracts) open into sinusoids are exposed to the highest concentrations of oxygen and molecules absorbed from the gut. These include the small molecules produced by digestion and any ingested toxic molecules or harmful microorganisms. Conversely, hepatocytes closest to central veins draining the sinusoids are exposed to lower levels of all these substances. Hepatocytes in different regions may appear similar but, in varying nutritional states and pathological conditions, their appearance may depend on their position in relation to blood supply.
Clinical notes
Chemotherapy Many chemotherapy drugs used to treat cancer target and kill cells in mitosis. As well as killing rapidly dividing cancer cells these drugs kill normal dividing cells such as those in epithelia lining the small (and large) intestine. Normally, intestinal epithelial cells are replaced every few days. After this type of chemotherapy there is impaired replacement of the columnar cells and goblet cells in the intestinal epithelia. As a result, absorption of nutrients and water is impaired and usually diarrhoea occurs.
Appendicitis The appendix is a blind-ended tube which may become inflamed due to stagnation and impaction of the contents of the gut. This results in appendicitis. If the inflammation destroys the wall of the appendix it spreads into the peritoneal cavity. As a result, microorganisms from the gut enter a large potential space where they can readily multiply: this may be fatal.
Gall bladder
Ducts draining bile from the liver eventually drain into a single bile duct which drains into the gall bladder. The gall bladder lies on the right, close to the liver, and stores and concentrates bile prior to its release. The gall bladder is lined by a simple columnar epithelium which absorbs fluid and concentrates the bile by up to 20 times. Smooth muscle forms the majority of the wall of the gall bladder and its contraction helps to expel stored bile. Parasympathetic nerves and molecules produced by neuroendocrine cells in the duodenum control the flow of bile in relation to the contents of the gastrointestinal tract. Bile expelled from the gall bladder enters the common bile duct, which drains into the duodenum.
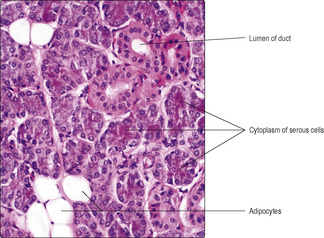
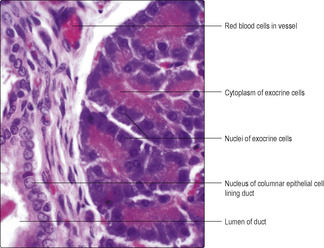
Pancreas
The pancreas lies in the abdominal cavity and functions as an exocrine and endocrine (
Chapter 14) gland. It has a thin connective tissue capsule and is divided into lobules by connective tissue septa. The exocrine cells are columnar epithelial cells (
Fig. 12.21) arranged as acini or tubules and they secrete bicarbonate ions (which help neutralise gastric acid) and pancreatic enzymes (proteinases, peptidases, lipases and amylases), which are inactive until they reach the duodenum. The enzymes are stored in zymogen granules in the apical portions of the columnar cells; the nucleus and rough endoplasmic reticulum are in the basal portions of the cells (
Fig. 12.21). The release of secretions is controlled by parasympathetic nerves and molecules secreted by neuroendocrine cells in the duodenum. A duct drains each acinus. Ducts join together within the lobules and eventually all join and form a single pancreatic duct. The duct system is lined by epithelial cells which, in the larger ducts, may be a double layer of columnar epithelial cells. The pancreatic duct joins the bile duct and bile and pancreatic juices drain into the duodenum.
Clinical note
The digestive system
■ The digestive system consists of the alimentary canal connecting the mouth to the anus and associated structures, e.g. salivary glands, liver and pancreas.
The alimentary canal
■ The alimentary canal has a general pattern of four layers:
■ a mucosa comprising an epithelium, basement membrane, lamina propria and a smooth muscle layer (the muscularis mucosae)
■ a submucosal layer supporting large blood vessels and nerves
■ a muscularis externa comprising an inner circular and an outer longitudinal layer of smooth muscle which contracts and moves the contents of the canal towards the anus
■ an outer covering which is a serosal (peritoneal) membrane in most regions.
Oesophagus
■ The oesophagus is lined by a stratified squamous epithelium and has serous and mucous glands in the submucosal layer.
■ It has striated voluntary muscle in the muscularis externa in regions close to the mouth and smooth muscle in lower regions.
■ It has an outer adventitia binding it to adjacent structures.
Stomach
■ It has three layers of smooth muscle in its wall and an outer serosal layer.
Small intestine
■ Epithelia covering villi and lining intestinal glands comprise columnar absorptive cells, goblet cells, stem cells and Paneth cells.
■ In the duodenum submucosal glands secrete alkaline mucus into the umen.
■ In the ileum lymphoid follicles are usually present.
Large intestine
■ A simple epithelium lines intestinal glands and contains absorptive columnar and goblet cells (no villi are present).
Structures associated with the digestive system
■ Salivary glands contain serous and mucous cells.
■ The tongue is covered by a stratified, squamous epithelium which, on its upper surface, has papillae and taste buds. Mucous and serous glands and lymphoid cells are deep to the epithelium and skeletal muscle forms the inner mass of the tongue.
■ The liver is formed mainly of hepatocytes:
■ it receives blood draining most parts of the alimentary canal and synthesises many new molecules and stores glycogen
■ it secretes bile which drains into the duodenum and aids digestion of fats
■ it destroys red blood cells and excretes some waste molecules in bile.
■ The pancreas is formed mainly by exocrine cells (endocrine cells are also present):
■ pancreatic enzymes (inactive when secreted) drain into the duodenum, where they begin to digest carbohydrates, proteins and lipids.
[/level-membership-for-basic-science-category][not-level-membership-for-basic-science-category]The digestive system consists of the alimentary canal, which is a tube connecting the mouth and anus, and associated structures that facilitate digestion of ingested food and drink (Fig. 12.1). The alimentary canal comprises the mouth, oesophagus and gastrointestinal tract. The gastrointestinal tract comprises the stomach, duodenum, jejunum, ileum, colon, appendix, rectum and the upper part of the anal canal. The associated structures are the teeth, tongue, salivary glands, liver and pancreas. At the mouth, lips intervene between skin and the mouth and they are covered by a stratified squamous epithelium which is not keratinised. At the anus the stratified, keratinised squamous epithelium of skin extends a short distance into the anal canal. The upper part of the anal canal is lined by a stratified, squamous epithelium which is not keratinised.
The digestive system has two primary functions. It breaks down food and drink into small molecules such as glucose, amino acids, fatty acids and triglycerides, a process which involves enzymic activity and is known as digestion. The second primary function of the digestive system ensures the small molecules produced by digestion in the lumen of the alimentary canal enter the body ‘proper’ by being absorbed across the epithelium lining the alimentary canal and into the blood or lymphatic vessels. In addition, the digestive system absorbs water, minerals, vitamins and ions from the material in the lumen of the gastrointestinal tract. The terminal part of the tract functions as a store for components of food that have not been digested. The stored, undigested food and waste products of the body, e.g. from the breakdown of red blood cells, are emptied from the alimentary canal at defecation.
The structure of the alimentary canal is related to its functions and different aspects of function occur at different locations along the canal. The gastrointestinal tract itself is several metres in length in humans, which is a reflection of the space required to accomplish all aspects of its function and is related to the transit times required for these functions.
General structure of the alimentary canal
All the alimentary canal is lined by a mucosa (
Chapter 3), which consists of an epithelium, connective tissue lamina propria and, in many regions, a layer of smooth muscle forming the muscularis mucosae (
Fig. 12.2). The epithelium of the mucosa reflects the primary function(s) occurring in that region. However, in all regions the mucosa is important in forming a barrier between substances ingested, including microorganisms, and the internal environment of the body. In regions where the muscularis mucosae is present, contraction of this muscle moves and folds the mucosa, aiding contact between the contents of the lumen and the surface epithelial cells.
A submucosal layer of connective tissue, which supports nerves and blood and lymph vessels, attaches the mucosa in most parts of the alimentary canal to outer layers of muscle (the muscularis externa) (
Fig. 12.2). In most regions of the alimentary canal two layers of muscle are present in the muscularis externa. In the outer, longitudinal layer the long axis of each muscle cell lies roughly parallel to the length of the lumen. In the inner, circular layer the long axes of the muscle cells lie around the lumen. Contractions of the muscularis externa cause peristalsis, which provides the motive force transporting the luminal contents of the canal towards the anus. In the submucosal layer and the muscularis externa, parasympathetic neuronal cell bodies are present and some are involved in co-ordinating muscle activity.
Specialised gut-associated lymphoid tissue (GALT) is present in the walls of the alimentary canal. In some regions, GALT forms large structures, e.g. tonsils; in other regions, only small clusters of lymphoid cells are present (
Fig. 12.2). These cells are part of the primary defence mechanisms against pathogens entering the body across the mucosa lining the gut tube.
Mouth
The mouth (oral cavity) contains the tongue and teeth. The epithelium of the mucosa lining the cheeks and tongue is able to resist the wear and tear in-volved in chewing food, and in most regions it is a stratified, squamous (non-keratinised) epithelium. This epithelial surface is moistened by secretions from small serous and mucous glands in the submucosal layer and from salivary glands (
Fig. 12.3) which drain their saliva into the oral cavity. Physical breakdown of food occurs in the mouth and it is mixed with salivary gland secretions which begin to break down large carbohydrate molecules.
Salivary glands
There are three, paired salivary glands, the parotid, submandibular and sublingual glands. Each gland is invested by a connective tissue capsule, and connective tissue forms trabeculae which penetrate the glands and provide support for the blood vessels and nerves supplying the glands and for the ducts draining them. Each gland has secretory cells arranged as compound, tubuloacinar, exocrine glands (
Chapter 3). The secretory acini drain into small ducts (lined by a simple cuboidal epithelium) which join together in a branching system; the larger ducts are lined by columnar epithelial cells, which may be stratified. The ducts drain the saliva into the oral cavity.
Two types of secretory cell, serous and mucous cells, are present in salivary gland acini (
Fig. 12.3) and, respectively, they produce a watery secretion containing proteins which function as enzymes and a viscous mucus in which large carbohydrate complexes are major components. In addition, myoepithelial cells wrap around the acini and their contractions help to expel the secretions. The secretions assist the process of digestion by moistening and lubricating the food, and by providing enzymes. The main enzyme is amylase, which breaks down large carbohydrate molecules. Saliva is also responsible for lubricating and cleansing the oral cavity. It contains bactericidal substances such as lysozyme secreted by serous cells and may contain immunoglobulin A secreted by plasma cells.
Submandibular glands
A submandibular gland lies beneath the mandible on each side, wrapped around the muscle which supports the tongue. Each gland drains via a duct that opens on the ipsilateral side of the ridge (the frenulum) on the undersurface of the tongue. Mucous glands are a prominent feature of submandibular glands, but serous cells are also present. The arrangement of the serous and mucous gland cells together in some acini is such that these are described as mucous glands with serous demilunes (
Fig. 12.3).
Sublingual glands
The sublingual glands are the smallest of the salivary glands. Mucous gland cells are predominant in sublingual glands and the ducts open directly into the floor of the oral cavity, rather than via a single duct system.
Tongue
The tongue is covered by a stratified, non-keratinised, squamous epithelium which on the undersurface is similar to the epithelium lining the mouth. The upper surface of the tongue is divided by a ‘V’-shaped groove into an anterior (two-thirds) and a posterior region developed from different parts of the embryo (see Mitchell B, Sharma R.
Embryology: An Illustrated Colour Text. Elsevier: 2004). The stratified squamous epithelium on the upper surface is studded by prominent projections. In the posterior region, many of the projections are due to aggregations of lymphocytes deep to the epithelium. In addition, three distinct types of projection known as papillae are also described, and all assist in macerating food and resisting abrasion:
■ Filiform papillae. These are numerous on the upper surface and are in parallel rows which converge towards the midline. The surface of these papillae is covered by a keratinised, stratified squamous epithelium.
■ Fungiform papillae. These are covered by a stratified (non-keratinised) squamous epithelium. Specialised clusters of sensory cells (taste buds) are present in the epithelium covering this type of papilla.
■ Circumvallate papillae. There are 8 to 12 large, circumvallate papillae visible to the unaided eye just anterior to the ‘V’-shaped groove of the tongue. They are also covered by a stratified (non-keratinised) squamous epithelium which contains taste buds.
Deep to the mucosa of the tongue there are numerous, small serous and mucous glands which secrete onto the surface of the epithelium. The major component of the inner part of the tongue is skeletal muscle (see
Fig. 5.7). The bundles of muscle cells are arranged in a complex three-dimensional meshwork and coordinated contraction of the muscle cells is important in chewing, swallowing and in sound production.
Pharynx and oesophagus
Swallowed food and drink pass via the pharynx to the oesophagus (gullet). The structure and function of the epithelium lining the component parts of the pharynx are described in
Chapter 11. Aggregations of lymphoid cells are a prominent feature of the walls of the pharynx and form tonsils (
Fig. 12.5). The lymphoid cells are important components of the immune mechanisms defending the alimentary canal (and the respiratory tract).
The major part of the oesophagus is in the thorax, but a small segment lies in the abdominal cavity and is continuous with the stomach. The structures in the walls of the oesophagus conform to the general plan for the gastrointestinal tract (
Fig. 12.2). In the thorax, the connective tissue of the adventitia binds the muscularis externa of the oesophagus to adjacent structures. The muscularis externa consists of an outer, approximately longitudinal layer and an inner circular layer. In the upper part of the muscularis externa, striated voluntary muscle (
Fig. 12.6) is present, whereas in the lower region it is replaced by smooth involuntary muscle. In the middle region there is a transition between the two types of muscle. Coordinated contraction of muscle cells in the different regions of the oesophagus ensures that swallowed food and drink normally pass to the stomach. In some regions, the submucosal tissue of the oesophagus contains serous and mucous glands in addition to blood vessels, nerves and aggregations of lymphoid cells. The epithelium of the mucosa lining the oesophagus is stratified and squamous (
Fig. 12.6) and is able to resist friction from swallowed food and chemical attack by swallowed drinks, e.g. alcohol.
The gastrointestinal tract
Stomach
Buy Membership for Basic Science Category to continue reading.
Learn more here
[/not-level-membership-for-basic-science-category]
Histology An Illustrated Colour Text