[level-membership-for-basic-science-category]Cells are the fundamental units of life. Textbooks may describe a ‘typical’ cell, but such a cell does not exist. Most cells are, to some extent, specialised in terms of their structure and function. Accordingly, the structural appearance of cells can provide information about their function. The term used to describe how cells are specialised is ‘differentiation’. Most cells have a nucleus which contains molecular programmes encoded in DNA (in chromosomes) that direct how a cell differentiates and what function(s) it performs. Many types of cell, even some which are differentiated, also have the ability to replicate themselves by a process of cell division known as mitosis (see below). Mitotic activity may continue in many types of cell throughout the life of the individ-ual. Cells undergoing successive rounds of mitosis are described as passing through a series of events known as the cell cycle (see below). Other cells may reach an end stage of differentiation and become unable to undergo mitosis and replicate themselves further. For example, shortly after birth nerve cells (neurons) cease division and replication.
Despite there being no ‘typical’ cell, cells share certain characteristics. All have a cell membrane which encloses cytoplasm; in most cells, the cytoplasm surrounds a nucleus. (Mature red blood cells in humans do not have a nucleus, nevertheless they live for about 100
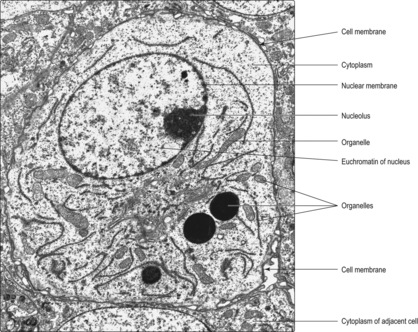
days.) Although the nucleus and cytoplasm may be clearly distinguished using a light microscope to examine cells in slices of tissue (Chapter 1), to resolve finer detail an electron microscope may be used. As the wavelength of electrons is much shorter than light waves, electron microscopy can resolve structures about 1000 times smaller than the light microscope can. Within cytoplasm, electron microscopy reveals a range of membrane-bound organelles and other structures (Fig. 2.1) which carry out a variety of functions during the life of a cell. The term ‘ultra-structure’ is used to describe structures revealed by electron microscopy. A brief survey of the ultrastructure of cells in relation to their function is given below.
Ultrastructure of cells and extracellular matrix
Cell membrane
The cell membrane is semi-permeable and allows inward and outward passage of selected substances. It also forms an essential barrier to the exterior and a boundary for the internal structure of the cell. It may be involved in attachments to adjacent cells (see below) and in recognition and communication within and outside the cell. Within, it may communicate with its cytoplasm and some signals may pass to the nucleus. Outside, it may communicate with other ‘self’ cells, both normal and abnormal (e.g. tumour cells or virally infected cells). Many cells interact, via their cell membrane, with microbes, and various molecules foreign to the body and a variety of cellular activities are stimulated or inhibited as a result.
Nucleus and nucleolus
The nucleus contains the vast majority of the DNA of the cell. The DNA is the hereditary material that has the genetic code expressed in a double strand of DNA in each chromosome. (Chromosomes contain proteins as well as DNA.) The number of chromosomes in a typical cell is species specific (humans have 46). Human chromosomes comprise 22 homologous pairs and a pair of sex chromosomes (two X chromosomes in females and an X and a Y chromosome in males). One of each pair of chromosomes is derived from the mother’s oocyte and the other from the father’s spermatozoon (see, respectively,
Chapters 16 and
15). The nucleus may also contain the structural and molecular mechanisms for the synthesis of RNA in one or more nucleoli.
The nucleus is enclosed by a nuclear membrane which is formed by two plasma membranes. In places, the nuclear membrane is perforated by pores which allow transport of material to and from the nucleus. The outer nuclear membrane is continuous in places with some membranes in the cytoplasm, and molecules (e.g. proteins and RNA) travel between nucleus and cytoplasm by this route.
The appearance of nuclei varies in relation to their function. Individual chromosomes are not apparent in cells unless the cell is dividing. The nuclear material, comprising DNA, proteins and RNA, is known as chromatin and two types are described, euchromatin and heterochromatin. Euchromatin appears less dense than heterochromatin (
Figs 2.1 and
2.2). The DNA molecules in euchromatin are uncoiled and are being used as coding for RNA synthesis which, in turn, directs protein synthesis in the cytoplasm. A large amount of euchromatin in a nucleus (
Fig. 2.1) indicates that a wide variety of RNA molecules are being made (and types of protein produced as a result). In contrast, in heterochromatin the DNA molecules are coiled and condensed and appear dense. The DNA in heterochromatin is mostly inactive, i.e. not directing the synthesis of RNA.
One or more nucleoli also appear as densely stained regions in the nuclei of cells actively synthesising proteins: their position may appear central or peripheral (
Fig. 2.1). RNA molecules synthesised in the nucleolus leave the nucleus, via pores in the nuclear membrane, and are involved in organising protein synthesis in the cytoplasm.
Cytoplasm
Cytoplasm comprises a fluid matrix, a cytoskeleton, various membrane-bound organelles and may include stored molecules.
■
Microtubules. Some microtubules are straight and long and allow the passage of substances within the cell. Many microtubules are polarised, i.e. they have specific ends with one end usually attached to a centrosome. Microtubules in some nerve cells transport molecules in a cytoplasmic process which extends several centimetres (
Chapter 6). Other microtubules assist in maintaining cell shape, compartmentalising the cytoplasm or facilitating movement of organelles within the cytoplasm. Microtubules are present in cilia, and in cells undergoing division (see below).
■
Intermediate filaments. The main role of this group of filaments involves resisting external stresses on the cytoplasm by their attachment to specific internal cell structures. They include the protein keratin in the epithelium of skin (
Chapter 7).
■
Thin (micro) filaments. These filaments are formed from the protein actin. Actin filaments are present in most cells and are involved in moving organelles within cells, in cell movement, and exocytosis and endocytosis (see below). One specific role of actin in muscle cells involves interaction with thicker protein filaments of myosin, which results in muscle contraction (
Chapter 5).
Cytoplasmic organelles
The main cytoplasmic organelles are mitochondria, endoplasmic reticulum, Golgi apparatus, vesicles and lysosomes. These organelles are involved in provision of energy for cellular processes, synthesis and secretion of protein, carbohydrate and lipid-based molecules, storage of proteins and degradation of waste material.
Mitochondria
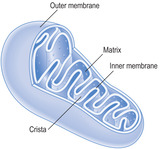
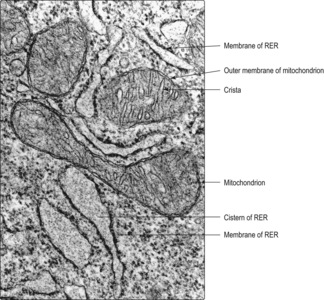
Mitochondria vary in shape but many are ovoid (
Figs 2.4 and
2.5). They are surrounded by an outer and inner membrane and provide most of the energy needs of the cell. The outer membrane is a typical plasma membrane, but the inner membrane has numerous in-foldings known as cristae. These folds increase the surface area inside each mitochondrion and provide a matrix in which metabolic processes occur. The matrix comprises a viscous fluid containing enzymes associated with the tricarboxylic acid (TCA) cycle. Mitochondria are involved in oxidative phosphorylation, which results in the production of adenosine triphosphate (ATP). ATP acts as a store of energy that is used for various cell activities. Mitochondria contain very small amounts of DNA in the matrix which code for some mitochondrial proteins.
Endoplasmic reticulum
Endoplasmic reticulum is a membrane system similar in appearance to the plasma membrane. It is double layered and encloses a space known as a cistern (
Fig. 2.5). In cells actively synthesising molecules, the cistern may be relatively wide. All membranes in the cytoplasm of a cell are probably linked and some link to the nuclear membrane. Two types of endoplasmic reticulum are described according to their appearance in electron micrographs.
■
Rough endoplasmic reticulum (RER). These membranes appear rough (
Fig. 2.5) as they are studded with ribosomes (see below). They are particularly prominent in cells synthesising proteins for export from the cell. Some proteins synthesised on RER are passed to the Golgi apparatus for further processing (see below).
■ Ribosomes are electron-dense structures containing RNA; some are attached to the membranes of RER (
Fig. 2.5), others lie free or form clusters in the cytoplasm (
Fig. 2.6). Specific proteins are synthesised by ribosomes from amino acids. The specificity is determined by coded RNA arriving from the DNA in the nucleus. In general, proteins produced on RER are for export and those produced by free ribosomes are for internal use.
Vesicles, endocytosis and exocytosis
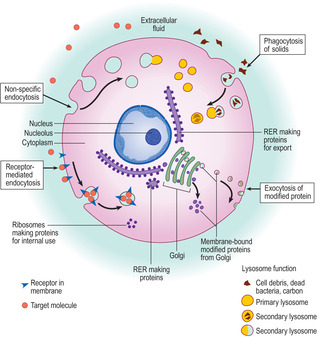
Membrane-bound vesicles are usually spherical. Many are involved in taking substances into and out of cells by the processes of endocytosis and exocytosis respectively (
Fig. 2.6). It is usual to distinguish two types of endocytosis: pinocytosis involves the uptake of fluid and phagocytosis the uptake of solids (but fluid may enter in the same vesicle). In addition, the process of endocytosis may be non-specific or specific. In non-specific endocytosis the cell membrane becomes invaginated and then encloses some extracellular material. Specific endocytosis involves receptor molecules on the cell membrane that are able to bind specific (target) molecules. In specific endocytosis only the part of the cell membrane with the receptor molecules (and their bound target molecules) invaginates and takes in the bound molecules; this process is known as receptor-mediated endocytosis (
Fig. 2.6). The fate of endocytotic vesicles varies; some may fuse with lysosomes (see below) or the contents may have a specific use within the cell. Exocytosis is the process by which the contents of membrane-bound vesicles are released from a cell. The membrane around the vesicle fuses with the cell membrane and the contents of the vesicle are released from the cell (
Fig. 2.6).
Golgi apparatus
The Golgi apparatus involves a stack of several parallel membranes (lying close to RER) and small membrane-bound vesicles (
Fig. 2.6). Newly synthesised proteins pass in vesicles from RER to the Golgi membranes. The proteins (in vesicles) then pass between the layers of Golgi membranes. During the time spent in the Golgi apparatus various carbohydrate molecules may be added to the newly synthesised proteins. Vesicles leaving the Golgi stack may move towards the cell membrane and discharge their contents by exocytosis or release them for internal use; others may remain stored in vesicles in the cell until they are needed.
Lysosomes
Lysosomes are membrane-bound organelles (Fig. 2.6) which are usually electron dense and thus appear dark in electron micrographs. They are formed via Golgi bodies and contain hydrolytic enzymes which are active at low (acidic) pH (e.g. acid phosphatases). Newly formed lysosomes are known as primary lysosomes and they may remain in a cell for some time before they become active. Lysosomes function by fusing with some endocytotic vesicles, other organelles or fragments of organelles, or other material; at this stage they become secondary lysosomes (
Fig. 2.6). Lysosomal enzymes are released onto the ingested material and break down most of the molecules they contained. The small molecules formed as a result may be reused by the cell; other substances which resist digestion (e.g. carbon) remain in the lysosome. The integrity of the membrane around lysosomes is important as it ensures that the hydrolytic lysosomal enzymes do not pass into the cytoplasm and destroy the cell itself.
Cytoplasmic processes
The cytoplasm of some cells forms specific processes which extend from the cells. These are microvilli, cilia and flagella.
■
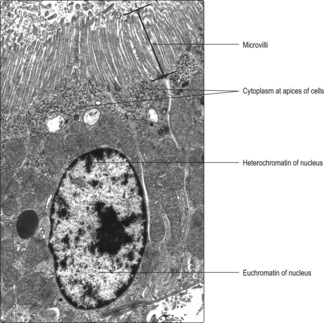
Microvilli (
Fig. 2.7) project as finger-like cytoplasmic extensions from many epithelial cells, e.g. from the apical surface of cells lining some gut tubes and others lining some tubules in the kidney. Actin microfilaments in microvilli help maintain their shape. The microvilli increase the area of cell membrane in contact with the contents of the tube and this aids absorption of molecules from the tube into the cell.
■
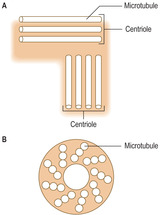
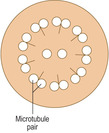
Cilia are cytoplasmic processes which extend from the cell membranes of some epithelial cells, e.g. cells in the epithelium lining tubes transporting air to the lungs. Cilia are usually wider and longer than microvilli. Cilia have microtubules extending along their length which have a symmetrical arrangement. The microtubules are in a 9 + 2 pattern, which is apparent when sectioned across their length (
Fig. 2.8). The tubules are involved in moving the cilia in a regular, synchronised beat which propels the material on the surface of the ciliated cells in a particular direction (
Chapter 11).
■
Flagella are similar to cilia and have symmetrically arranged microtubules but are much longer and wider than cilia. In humans, the only cells which have a flagellum are spermatozoa (the male gametes). Each spermatozoon is propelled by the beating of its flagellum as it travels the length of the female reproductive tract after copulation (
Chapters 15 and
16).
Molecules stored in cytoplasm
Carbohydrates and lipids are stored in cytoplasm; most are not enclosed by a membrane. Glycogen is stored in many cells and acts as an energy reserve. It appears as dense particles in electron micrographs, some of which are clustered together and may occupy extensive regions of the cytoplasm. Lipids are stored in some cells generally as spherical masses. Some stored lipids act as precursor molecules for the synthesis of steroid hormones, others provide energy reserves. Proteins and glycoproteins are usually stored in membrane-bound vesicles and their contents discharged by exocytosis when needed.
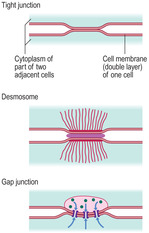
Intercellular junctions
There are three specialised ways in which adjacent cells are attached to each other (
Fig. 2.9).
■ Tight junctions (zona occludens). In these junctions a short length of the outer cell membrane layers of adjacent cells is fused. These junctions prevent molecules from passing in between cells.
■
Gap junctions. At these junctions membranes from adjacent cells are closely applied but they are not fused. Gap junctions allow the passage of ions and very small molecules from one cell to an adjacent cell. They are particularly important between heart muscle cells as ion movement between such cells is an essential part of coordinating contraction of heart muscle (
Chapter 10).
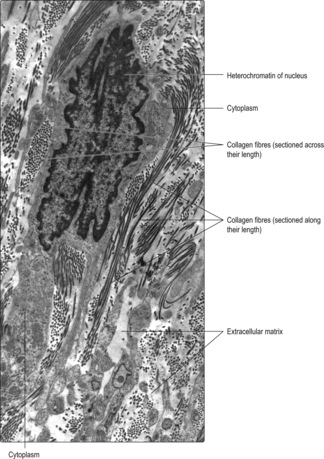
Extracellular matrix
Many cells are adjacent to extracellular matrix and some are entirely surrounded by it. Extracellular matrix varies in form and function in different parts of the body (
Chapter 4). Some proteins (e.g. collagen) form fibres in extracellular matrix, and their appearance depends on the angle at which they are sectioned (
Fig. 2.2).
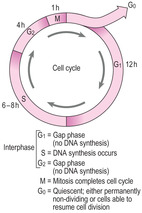
The cell cycle and cell division
The cell cycle is the series of stages through which a cell passes between successive rounds of the cell division process known as mitosis (
Fig. 2.10). The actual event of mitosis takes a relatively short time in the cycle, sometimes less than an hour. The stages a cell passes through before successive mitotic divisions are collectively known as interphase and take considerably longer (several hours or days). During interphase, events occur which may relate to the specific functions of the cell, e.g. protein synthesis and secretion. A cell in interphase must also undergo specific processes to replicate its DNA in preparation for mitosis.
The interphase of the cell cycle is divided into three stages (
Fig. 2.10). After mitosis, cells enter the G
1 phase, which is often the longest phase of the cycle. During G
1, cells synthesise many molecules, increase the volume of the cytoplasm and increase the number of organelles. These synthetic processes may be specific for a cell to carry out its functions as well as to produce molecules necessary for the next phase. At the end of G
1, the cells move into the next phase (the S phase) when they begin to synthesise new DNA. During the S phase the new DNA is assembled along the length of the existing double strand of DNA in each chromosome by the process of base pairing; this duplicates the existing DNA. At the end of the S phase, instead of a double strand of DNA in each chromosome there are four strands of DNA (arranged as two chromatids). When DNA replication is completed, the cells move into the G
2 phase, which is when the cell prepares for mitosis. The G
2 phase is usually much shorter than the G
1 phase.
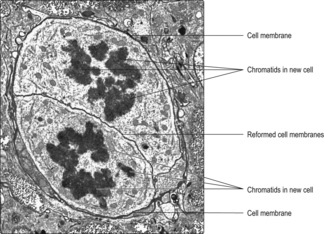
A cell which undergoes a mitotic cell division becomes two identical offspring cells with the same number of chromosomes containing the same amount and type of DNA as the original cell. Mitosis may be described in five stages.
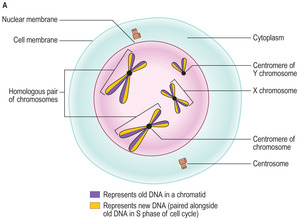
■
Prophase. During this stage the DNA in chromosomes becomes very coiled and condensed and appears as very dense chromatin. The nucleolus disappears and RNA production ceases. Towards the end of prophase (
Fig. 2.11A) chromosomes appear as two parallel chromatids joined at a point known as the centromere. Each chromatid contains an old strand of DNA and the newly synthesised copy strand. At this time, the centrosome of the cell has been duplicated and they move to opposite poles of the cell. Part of each centrosome begins to give rise to an array of microtubules which will form a mitotic spindle (see below).
■ Prometaphase. This stage begins with the disappearance of the nuclear membrane. During this phase chromosomes are still randomly arranged in the cytoplasm. A mitotic spindle forms and each chromosome begins to move towards the microtubules at the equator of the spindle.
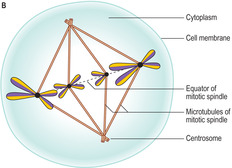
■
Metaphase. In this stage (
Fig. 2.11B) each chromosome attaches (by its centromere) at a separate point on the equator of the spindle, an arrangement described as forming a metaphase plate. Such plates are readily visible with the light microscope in actively dividing cells.
■
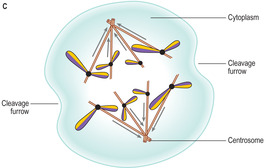
Anaphase. During anaphase the chromosomes split along their length separating the two chromatids which then move away from each other and towards opposite poles of the spindle (
Fig. 2.11C). The separation of the chromatids follows the track of the microtubules of the spindle. Towards the end of anaphase a depression begins to develop around the cytoplasm and it is known as the cleavage furrow.
On completion of the cell cycle, the two new cells may enter a G
1 phase and prepare for further rounds of the cycle. Some cells may suspend their progression around the cell cycle (usually at the G
1 stage) and enter a G
0 phase (
Fig. 2.10). Some cells in the G
0 phase may eventually re-enter the cycle and replicate themselves when, for example, they are needed to replace dead or damaged similar cells. Other cells halt their progression around the cycle, undergo differentiation and carry out specific functions before they die without having divided again.
Meiosis
Meiosis is a type of cell division in which the number of chromosomes (and DNA) in the offspring cells is half that in the parent cell. This occurs only in the formation of gametes, i.e. ova in females and spermatozoa in males. Meiosis ensures that on fertilisation the normal number of chromosomes (and their DNA content) is present in the fertilised ovum (the zygote). The process of meiosis ensures that there is a mixing of maternally and paternally derived chromosomes (and thus genes). This ensures diversity of the gene pool of a species. Further details of meiosis are described in Chapters 15 and
16.
■ A cell is the basic unit of life formed from cytoplasm and a nucleus (in most cells) enclosed by a cell membrane.
■ The nucleus contains DNA (in chromosomes), proteins and RNA.
■ In non-dividing cells, nuclear contents are described as chromatin:
■ heterochromatin is densely stained and is largely inactive
■ euchromatin is palely stained as the DNA is uncoiled and coding RNA to direct protein synthesis
■ a nucleolus is present in nuclei of cells producing RNA.
■ Cytoplasm contains organelles which carry out specific functions:
■ tubules and protein filaments give shape to cells and aid movement (of material in cells and of cells themselves)
■ mitochondria provide energy (ATP) for the cell
■ endoplasmic reticulum, which is formed from membranes surrounding a cistern, is involved in synthesis. RER has bound ribosomes and synthesises proteins mainly for export; SER is involved in lipid synthesis. Free ribosomes synthesise proteins mainly for internal use
■ membrane-bound vesicles are involved in uptake of substances into the cell (endocytosis) and export of substances (exocytosis)
■ the Golgi apparatus has membranes and vesicles and modifies synthesised proteins by adding, for example, carbohydrates
■ lysosomes are membrane bound and contain hydrolytic enzymes. Lysosomes fuse with endocytotic vesicles and digest their contents.
■ Cytoplasm in some cells forms projections, for example cilia and microvilli, and may store carbohydrate, lipids or proteins.
■ Specialised parts of cell membranes of adjacent cells form junctions which help to bind the cells together, prevent molecules moving in between the cells, or aid movement of ions between adjacent cells.
■ Cell division (mitosis) occurs in many types of cell. Prior to mitosis, cells synthesise copy strands of their DNA in the S phase of the cell cycle:
■ In the prophase stage of mitosis chromosomes condense and become visible as individual units. A spindle forms of microtubules. Gradually, the nuclear membrane disappears and (at metaphase) the chromosomes move to the equator of the spindle and attach to the microtubules. In the next phase (anaphase) the chromosomes split along their length and each half moves towards a pole of the spindle. Mitosis ends with telophase as the spindle disappears and the nuclear membrane reforms and encloses the uncoiling chromosomes. The cytoplasm of the original cell splits and two new cells are formed with each having the same number of chromosomes (and amount of DNA) as the original cell.
[/level-membership-for-basic-science-category][not-level-membership-for-basic-science-category]Cells are the fundamental units of life. Textbooks may describe a ‘typical’ cell, but such a cell does not exist. Most cells are, to some extent, specialised in terms of their structure and function. Accordingly, the structural appearance of cells can provide information about their function. The term used to describe how cells are specialised is ‘differentiation’. Most cells have a nucleus which contains molecular programmes encoded in DNA (in chromosomes) that direct how a cell differentiates and what function(s) it performs. Many types of cell, even some which are differentiated, also have the ability to replicate themselves by a process of cell division known as mitosis (see below). Mitotic activity may continue in many types of cell throughout the life of the individ-ual. Cells undergoing successive rounds of mitosis are described as passing through a series of events known as the cell cycle (see below). Other cells may reach an end stage of differentiation and become unable to undergo mitosis and replicate themselves further. For example, shortly after birth nerve cells (neurons) cease division and replication.
Despite there being no ‘typical’ cell, cells share certain characteristics. All have a cell membrane which encloses cytoplasm; in most cells, the cytoplasm surrounds a nucleus. (Mature red blood cells in humans do not have a nucleus, nevertheless they live for about 100
days.) Although the nucleus and cytoplasm may be clearly distinguished using a light microscope to examine cells in slices of tissue (Chapter 1), to resolve finer detail an electron microscope may be used. As the wavelength of electrons is much shorter than light waves, electron microscopy can resolve structures about 1000 times smaller than the light microscope can. Within cytoplasm, electron microscopy reveals a range of membrane-bound organelles and other structures (Fig. 2.1) which carry out a variety of functions during the life of a cell. The term ‘ultra-structure’ is used to describe structures revealed by electron microscopy. A brief survey of the ultrastructure of cells in relation to their function is given below.
Ultrastructure of cells and extracellular matrix
Cell membrane
The cell membrane is semi-permeable and allows inward and outward passage of selected substances. It also forms an essential barrier to the exterior and a boundary for the internal structure of the cell. It may be involved in attachments to adjacent cells (see below) and in recognition and communication within and outside the cell. Within, it may communicate with its cytoplasm and some signals may pass to the nucleus. Outside, it may communicate with other ‘self’ cells, both normal and abnormal (e.g. tumour cells or virally infected cells). Many cells interact, via their cell membrane, with microbes, and various molecules foreign to the body and a variety of cellular activities are stimulated or inhibited as a result.
Nucleus and nucleolus
The nucleus contains the vast majority of the DNA of the cell. The DNA is the hereditary material that has the genetic code expressed in a double strand of DNA in each chromosome. (Chromosomes contain proteins as well as DNA.) The number of chromosomes in a typical cell is species specific (humans have 46). Human chromosomes comprise 22 homologous pairs and a pair of sex chromosomes (two X chromosomes in females and an X and a Y chromosome in males). One of each pair of chromosomes is derived from the mother’s oocyte and the other from the father’s spermatozoon (see, respectively,
Chapters 16 and
15). The nucleus may also contain the structural and molecular mechanisms for the synthesis of RNA in one or more nucleoli.
The nucleus is enclosed by a nuclear membrane which is formed by two plasma membranes. In places, the nuclear membrane is perforated by pores which allow transport of material to and from the nucleus. The outer nuclear membrane is continuous in places with some membranes in the cytoplasm, and molecules (e.g. proteins and RNA) travel between nucleus and cytoplasm by this route.
The appearance of nuclei varies in relation to their function. Individual chromosomes are not apparent in cells unless the cell is dividing. The nuclear material, comprising DNA, proteins and RNA, is known as chromatin and two types are described, euchromatin and heterochromatin. Euchromatin appears less dense than heterochromatin (
Figs 2.1 and
2.2). The DNA molecules in euchromatin are uncoiled and are being used as coding for RNA synthesis which, in turn, directs protein synthesis in the cytoplasm. A large amount of euchromatin in a nucleus (
Fig. 2.1) indicates that a wide variety of RNA molecules are being made (and types of protein produced as a result). In contrast, in heterochromatin the DNA molecules are coiled and condensed and appear dense. The DNA in heterochromatin is mostly inactive, i.e. not directing the synthesis of RNA.
One or more nucleoli also appear as densely stained regions in the nuclei of cells actively synthesising proteins: their position may appear central or peripheral (
Fig. 2.1). RNA molecules synthesised in the nucleolus leave the nucleus, via pores in the nuclear membrane, and are involved in organising protein synthesis in the cytoplasm.
Cytoplasm
Cytoplasm comprises a fluid matrix, a cytoskeleton, various membrane-bound organelles and may include stored molecules.
■
Microtubules. Some microtubules are straight and long and allow the passage of substances within the cell. Many microtubules are polarised, i.e. they have specific ends with one end usually attached to a centrosome. Microtubules in some nerve cells transport molecules in a cytoplasmic process which extends several centimetres (
Chapter 6). Other microtubules assist in maintaining cell shape, compartmentalising the cytoplasm or facilitating movement of organelles within the cytoplasm. Microtubules are present in cilia, and in cells undergoing division (see below).
■
Intermediate filaments. The main role of this group of filaments involves resisting external stresses on the cytoplasm by their attachment to specific internal cell structures. They include the protein keratin in the epithelium of skin (
Chapter 7).
■
Thin (micro) filaments. These filaments are formed from the protein actin. Actin filaments are present in most cells and are involved in moving organelles within cells, in cell movement, and exocytosis and endocytosis (see below). One specific role of actin in muscle cells involves interaction with thicker protein filaments of myosin, which results in muscle contraction (
Chapter 5).
Cytoplasmic organelles
The main cytoplasmic organelles are mitochondria, endoplasmic reticulum, Golgi apparatus, vesicles and lysosomes. These organelles are involved in provision of energy for cellular processes, synthesis and secretion of protein, carbohydrate and lipid-based molecules, storage of proteins and degradation of waste material.
Mitochondria
Mitochondria vary in shape but many are ovoid (
Figs 2.4 and
2.5). They are surrounded by an outer and inner membrane and provide most of the energy needs of the cell. The outer membrane is a typical plasma membrane, but the inner membrane has numerous in-foldings known as cristae. These folds increase the surface area inside each mitochondrion and provide a matrix in which metabolic processes occur. The matrix comprises a viscous fluid containing enzymes associated with the tricarboxylic acid (TCA) cycle. Mitochondria are involved in oxidative phosphorylation, which results in the production of adenosine triphosphate (ATP). ATP acts as a store of energy that is used for various cell activities. Mitochondria contain very small amounts of DNA in the matrix which code for some mitochondrial proteins.
Buy Membership for Basic Science Category to continue reading.
Learn more here
[/not-level-membership-for-basic-science-category]
Histology An Illustrated Colour Text