27 Thalamus, epithalamus
Thalamus
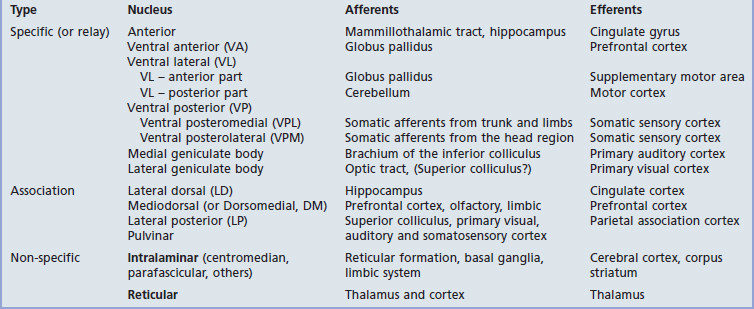
The thalamus is the largest nuclear mass in the entire nervous system. It is a prominent feature in MRI scans in each of the three planes in which slices are taken. The afferent and efferent connections of the main nuclear groups are listed in Table 27.1. The connections are so diverse that the thalamus cannot be said to have a unitary function.
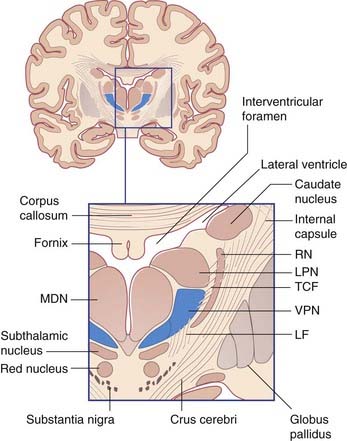
As noted in Chapter 2, the two thalami lie at the center of the brain. Their medial surfaces are usually linked across the third ventricle and their lateral surfaces are in contact with the posterior limb of the internal capsule. The upper surface of each occupies the floor of a lateral ventricle. The under-aspect receives sensory and cerebellar inputs as well as an upward continuum of the reticular formation.
Thalamic nuclei
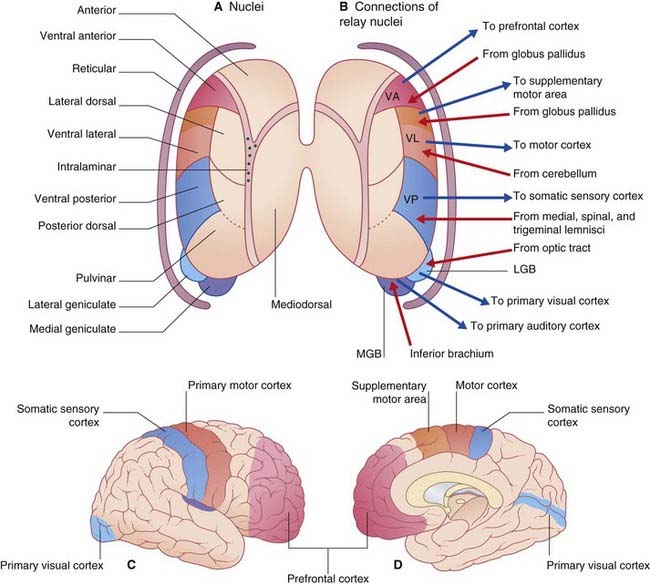
All thalamic nuclei except one (the reticular nucleus) have reciprocal excitatory connections with the cerebral cortex. The Y-shaped internal medullary lamina of white matter divides the thalamus into three large cell groups: medial dorsal, anterior, and lateral (Figure 27.1A). The lateral group comprises dorsal and ventral nuclear tiers. At the back of the thalamus are the medial and lateral geniculate bodies. The external medullary lamina separates the thalamus from the shell-like reticular nucleus.
Specific nuclei
The specific or relay nuclei are reciprocally connected to specific motor or sensory areas of the cerebral cortex. They comprise the nuclei of the ventral tier and the geniculate bodies (nuclei). Their afferent and efferent connections are indicated in Figure 27.1B.
The anterior nucleus receives the mammillothalamic tract and projects to the cingulate cortex. It is involved in a limbic circuit and has a function in relation to memory (Ch. 34).
The ventral posterior nucleus (VP) receives all of the fibers of the medial, spinal, and trigeminal lemnisci (Figure 27.2). It projects to the somatic sensory cortex (SI). A smaller projection is sent to the second somatic sensory area (SII) at the foot of the postcentral gyrus (see Ch. 29).
The VP is somatotopically arranged, as indicated in Figure 27.3. The portion of the nucleus devoted to the face and head is called the ventral posterior medial nucleus (VPM), that for the trunk and limbs the ventral posterior lateral nucleus (VPL). Modality segregation is a feature of both nuclei, with proprioceptive neurons most anterior, tactile neurons in the midregion, and nociceptive neurons at the back. The nociceptive region is sometimes called the posterior nucleus.
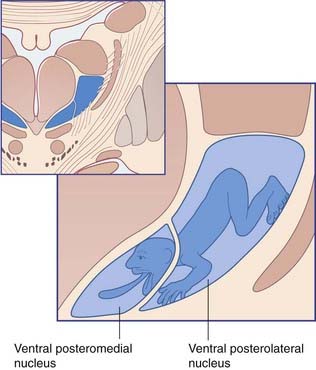
Figure 27.3 Somatic sensory map in the ventral posterior thalamic nucleus.
(Redrawn and modified from Ohye 1990, with permission.)
There is no evidence in the VP of an antinociceptive mechanism comparable to that found in the substantia gelatinosa region of the spinal cord and spinal trigeminal nucleus. An unexplained disorder, the thalamic syndrome, may follow a vascular lesion that disconnects the posterior thalamic nucleus from the somatic sensory cortex. In this condition, a period of complete sensory loss may occur on the contralateral side of the body, to be replaced by bouts of severe pain occurring either spontaneously or in response to tactile stimuli. (See also Ch. 35, central poststroke pain.)
The medial geniculate body (medial geniculate nucleus) is the thalamic nucleus of the auditory pathway. It receives the inferior brachium from the inferior colliculus (which carries auditory signals from both ears, Ch. 20), and it projects to the primary auditory cortex in the superior temporal gyrus.
The lateral geniculate body (lateral geniculate nucleus) is the principal thalamic nucleus for vision. It receives retinal inputs from both eyes by way of the optic tract, and it projects to the primary visual cortex in the occipital lobe. The visual pathways are described in Chapter 28.
Association nuclei
The association nuclei are reciprocally connected to the association areas of the cerebral cortex.
The lateral dorsal nucleus has reciprocal connections with the posterior part of the cingulate cortex, which is involved in functions related to memory (Ch. 34).
Non-specific nuclei
The intralaminar nuclei are contained within the internal medullary lamina of white matter. They can be regarded as a rostral continuation of the reticular formation of the midbrain (ascending reticular activating system in Ch. 24). They project widely to the cerebral cortex, as well as to the corpus striatum.
Afferents belonging to the ascending reticular activating system synapse in the intralaminar nuclei, also in the reticular nucleus and in the nucleus of Meynert in the basal forebrain (Ch. 34).
The thalamic reticular nucleus (TRN) is shaped like a shield around the front and lateral side of the thalamus. It is separated from the main thalamus by the external medullary lamina. All of the thalamocortical projections from the specific thalamic nuclei pass through TRN and give collateral branches to it (Figure 27.4). Fusiform neurons within the innermost lamina (VI) of the cerebral cortex project to the thalamic nuclei and also give off collaterals to TRN.
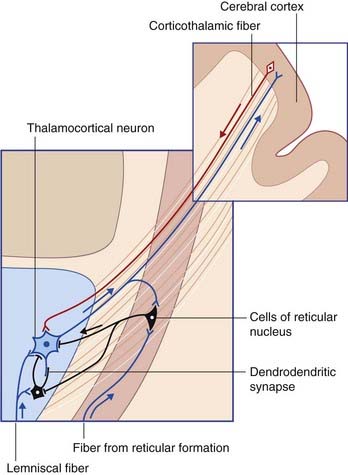
Oscillation
Sleep–wake cycles are described in Chapter 30.
Not represented in Table 27.1 are aminergic afferents passing to the ventral and intralaminar nuclei, from the midbrain raphe (serotonergic) and cerulean nucleus (noradrenergic). The proven value of tricyclic antidepressants in the therapy of chronic pain may be related to drug-induced prolongation of excitatory aminergic effects on thalamocortical neurons.
Thalamic peduncles
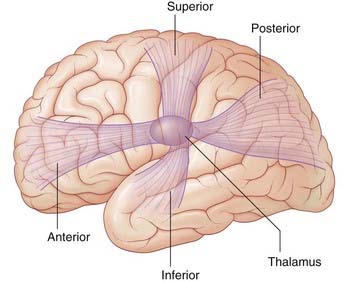
The reciprocal connections between the thalamus and the cerebral cortex travel in four thalamic peduncles, as shown in Figure 27.5. The anterior thalamic peduncle passes through the anterior limb of the internal capsule to reach the prefrontal cortex and cingulate gyrus. The superior thalamic peduncle passes through the posterior limb of the internal capsule to reach the premotor, motor, and somatic sensory cortex. The posterior thalamic peduncle passes through the retrolentiform part of the internal capsule to reach the occipital lobe and the posterior parts of the parietal and temporal lobes. The inferior thalamic peduncle passes below the lentiform nucleus to reach the anterior temporal and orbital cortex. Each of the four fans becomes incorporated into the corona radiata.
Epithalamus
The epithalamus includes the pineal gland (considered in Ch. 26); and the habenula and stria medullaris which are included with the limbic system in Chapter 34.
Core Information
Thalamus
Alitto HJ, Usry WM. Corticothalamic feedback and sensory processing. Curr Opin Neurobiol. 2003;13:440-445.
Benarroch E. The midline and intralaminar thalamic nuclei. Neurology. 2008;71:944-949.
Hall AJ, Lomber SG. Auditory cortex projections target the peripheral field representation of primary visual cortex. J Exp Neurol. 2008;190:413-430.
Ohye C. Thalamus. In: Paxinos G, editor. The human nervous system. ed 2. San Diego: Academic Press; 1990:439-468.
Pinault D. The thalamic reticular nucleus: structure, function and concept. Brain Res Rev. 2004;46:1-31.