[level-membership-for-basic-science-category]
CHAPTER 66 Small intestine
DUODENUM
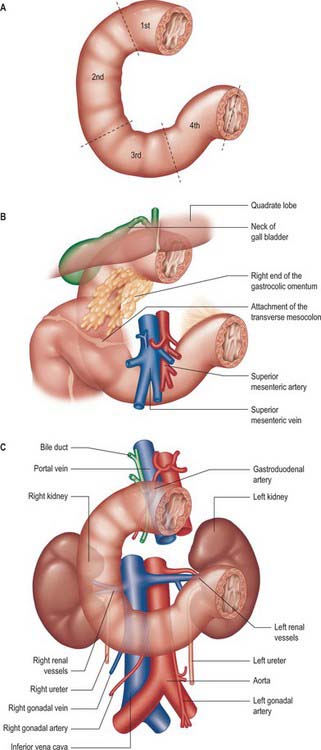
The adult duodenum is 20–25 cm long and is the shortest, widest and most predictably placed part of the small intestine. It is only partially covered by peritoneum, although the extent of the peritoneal covering varies along its length: the proximal 2.5 cm is intraperitoneal, and the remainder is retroperitoneal. The duodenum forms an elongated ‘C’ that lies between the level of the first and third lumbar vertebrae in the supine position. The lower ‘limb’ of the C extends further to the left of the midline than the upper limb. The head and uncinate process of the pancreas lie within the concavity of the duodenum which is ‘draped’ over the prominence formed from the lumbar spine, the duodenum therefore bends in an antero-posterior direction as well as following the form of a ‘C’. The duodenum lies entirely above the level of the umbilicus and is described as having four parts (Figs 66.1, 66.2).
FIRST (SUPERIOR) PART
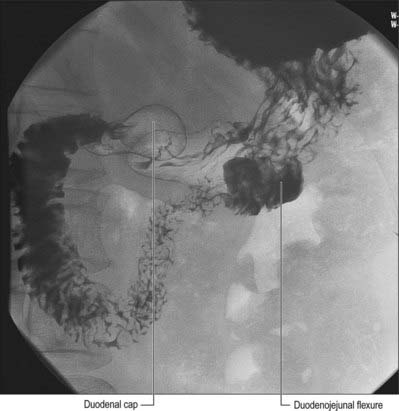
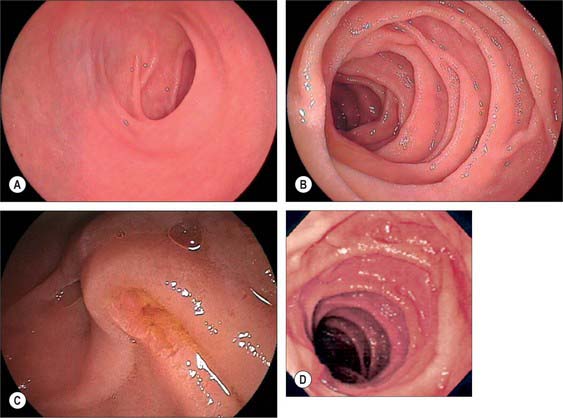
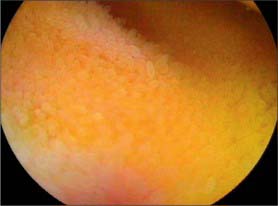
The first, and most mobile, part of the duodenum is about 5 cm long. It starts as a continuation of the duodenal end of the pylorus and ends at the superior duodenal flexure. Peritoneum covers the anterior and superior part of its posterior aspect close to the pylorus, where the duodenum forms part of the anterior wall of the epiploic foramen. Here the lesser omentum is attached to its upper border and the greater omentum to its lower border. The first 2 or 3 cm have a bland internal mucosal appearance and readily distend on insufflation during endoscopy. This part is frequently referred to as the duodenal ‘cap’. It has a triangular, homogeneous appearance during contrast radiology, shows the same pattern of internal rugae as the pylorus, and is often visible on plain radiographs of the abdomen as an isolated triangular gas shadow to the right of the first or second lumbar vertebra. The duodenum next passes superiorly, posteriorly and laterally for 5 cm before curving sharply inferiorly into the superior duodenal flexure: it rapidly becomes more retroperitoneal during this part of its course, until peritoneum only covers its anterior aspect. From the end of the duodenal cap, its internal appearance is characterized by extensive, deep mucosal folds that involve up to half of the circumference of the lumen. Even during endoscopic insufflation, these folds are pronounced (Fig. 66.3) and they are readily seen on contrast radiographs. The section from the duodenal cap to the superior duodenal flexure lies posterior and inferior to the quadrate lobe of the liver.
SECOND (DESCENDING) PART
The second part lies anterior to the hilum of the right kidney, the right renal vessels, the edge of the inferior vena cava and the right psoas major. The head of the pancreas and the common bile duct are medial and the hepatic flexure is above and lateral. A small part of the pancreatic head is sometimes embedded in the medial duodenal wall, and pancreatic ‘rests’ in the duodenal wall may produce small filling defects on contrast radiology. The internal appearance is similar to that of the distal portion of the first part of the duodenum, with pronounced mucosal folds (Fig. 66.3). The common bile duct and pancreatic duct enter the medial wall obliquely and usually unite to form the common hepatopancreatic ampulla. The narrow distal end opens on the summit of the major duodenal papilla (ampulla of Vater), which is situated on the posteromedial wall of the second part, 8–10 cm distal to the pylorus. There are variants of the ductal anatomy. The most common is a second, accessory pancreatic duct which may open 2 cm above the major papilla on a minor duodenal papilla. Peptic ulceration of the second part of the duodenum is less common than that of the first part, and tends to occur on the anterior or lateral wall.
FOURTH (ASCENDING) PART
The fourth part of the duodenum is 2.5 cm long. It starts just to the left of the aorta, runs superiorly and laterally to the level of the upper border of the second lumbar vertebra, then turns sharply anteroinferiorly at the duodenojejunal flexure to become continuous with the jejunum. The main trunk of the inferior mesenteric vein lies either posterior to the duodenojejunal flexure or beneath the adjacent peritoneal fold. (The duodenojejunal flexure is a useful landmark to locate the vein radiologically or surgically.) The aorta, left sympathetic trunk, left psoas major, left renal and left gonadal vessels are all posterior relations, the left kidney and left ureter are posterolateral, and the upper part of the root of the small bowel mesentery, the left lateral transverse mesocolon and transverse colon, which separate it from the stomach, are anterior. The peritoneum of the root of the small bowel mesentery continues over its anterior surface. The lower border of the body of the pancreas is superior.
At its left lateral end, the fourth part becomes progressively covered in peritoneum on its superior and inferior surfaces, such that it is suspended from the retroperitoneum by a double fold of peritoneum, the ligament of Treitz, at the start of the duodenojejunal flexure. The ligament of Treitz may contain a small slip of muscle, the suspensory muscle of the duodenum. When present, the suspensory muscle contains skeletal muscle fibres that run from the left crus of the diaphragm to connective tissue around the coeliac axis, and smooth muscle fibres that run from the coeliac axis: its function is unknown. It is an important landmark in the radiological diagnosis of incomplete rotation and malrotation of the small intestine (see Ch. 73). The ligament of Treitz does not form part of the duodenal mesentery because the vascular supply to the fourth part of the duodenum continues to enter its wall from the posteromedial aspect.
VASCULAR SUPPLY AND LYMPHATIC DRAINAGE
Arteries
The main vessels supplying the duodenum are the superior and inferior pancreaticoduodenal arteries. The first and second parts also receive contributions from several sources including the right gastric, supraduodenal, right gastroepiploic, hepatic and gastroduodenal arteries. Branches of the superior pancreaticoduodenal artery may contribute to the supply of the pyloric canal, with a minor degree of anastomosis to the gastric arteries in the muscular layer across the pyloroduodenal junction (Fig. 66.4).
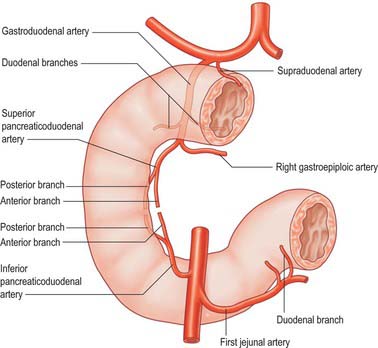
Superior pancreaticoduodenal arteries
The superior pancreaticoduodenal artery is usually double. The anterior artery is a terminal branch of the gastroduodenal artery and descends in the anterior groove between the second part of the duodenum and the head of the pancreas. It supplies branches to the first and second parts of the duodenum and to the head of the pancreas, and anastomoses with the anterior division of the inferior pancreaticoduodenal artery. The posterior artery is usually a separate branch of the gastroduodenal artery and is given off at the upper border of the first part of the duodenum. It descends to the right, anterior to the portal vein and common bile duct as the latter lies behind the first part of the duodenum. It then runs behind the head of the pancreas, crosses posterior to the common bile duct (which is embedded in the head of the pancreas), enters the duodenal wall and anastomoses with the posterior division of the inferior pancreaticoduodenal artery. The posterior artery supplies branches to the head of the pancreas, the first and second parts of the duodenum, and several branches to the lowest part of the common bile duct.
JEJUNUM
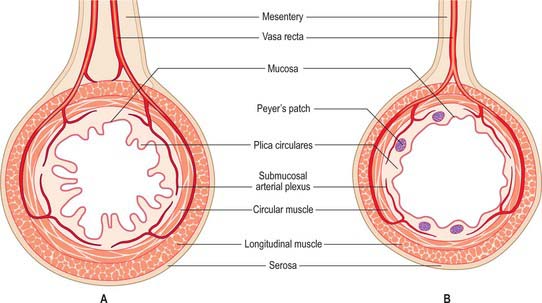
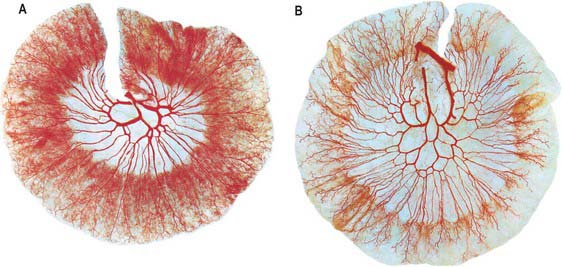
The jejunum has a median external diameter of 4 cm and an internal diameter of 2.5 cm. It has a thicker wall than the ileum, and an extensive arterial blood supply that makes it appear redder than the ileum in the living. The plicae circulares are most pronounced in the proximal jejunum, where they are more numerous and deeper than elsewhere in the small bowel (Fig. 66.5A). They frequently ‘branch’ around the lumen and may appear to be ranged one on top of another, giving the jejunum a characteristic appearance during single contrast radiography (Fig. 66.6). Lymphoid aggregates are almost absent from the proximal jejunum, but are present distally, although they are fewer in number and smaller than in the ileum. Where present, they are usually discoid in shape and impalpable.
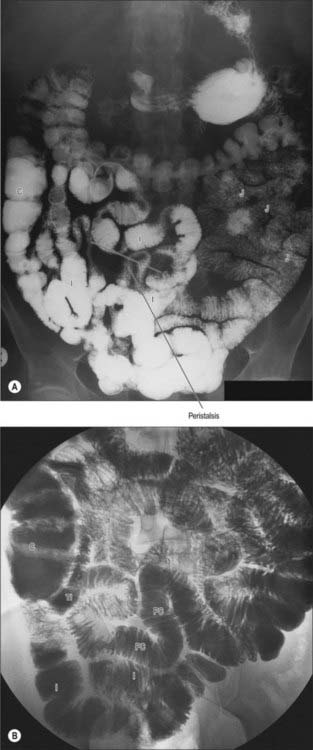
In the supine position the jejunum usually occupies the upper left infracolic compartment, extending down to the umbilical region. The first one or two loops often occupy a recess between the left part of the transverse mesocolon and the left kidney. On supine radiological examination, the jejunal loops are characteristically situated in the upper abdomen, to the left of the midline, whereas the ileal loops tend to lie in the lower right part of the abdomen and pelvis. This distribution can be reversed during paralytic ileus or small bowel obstruction due to rotation around the mesenteric attachment following bowel distension.
ILEUM
The ileum has a median external diameter of 3.5 cm and an internal diameter of 2 cm, and tends to have a thinner wall than the jejunum (Fig. 66.5B). The plicae circulares become progressively less obvious in the distal mucosa in the ileum; they tend to be single and flatter with less pronounced crests (Figs 66.6, 66.7), and the mucosa of the terminal ileum immediately proximal to the ileocaecal valve may appear almost flat. Lymphoid aggregates are larger and more numerous than in the jejunum and may be easily palpable in the terminal ileum. They are most prominent in early childhood and, when enlarged in viral infections, may form the apex of an ileo-ileal intussusception. They become less prominent prior to puberty, and attain adult morphology by late teenage years. In the supine position, the ileum lies mainly in the hypogastric region and right iliac fossa. The terminal ileum frequently lies in the pelvis, whence it ascends over the right psoas major and right iliac vessels, to end by opening into the ileocaecal valve in the right iliac fossa.
VASCULAR SUPPLY AND LYMPHATIC DRAINAGE
Arteries
Branches from the superior mesenteric artery supply the jejunum and ileum. The arteries divide as they approach the mesenteric border, giving off numerous branches which extend between the serosal and muscular layers, supplying the muscle and forming an intricate sub-mucosal plexus that supplies the glands and villi. Although there is a profuse anastomotic network of arteries within the mesentery, anastomoses between the terminal branches close to the intestinal wall are few (Fig. 66.8). The intramural and submucosal arterial networks are not of large calibre, which means that flow along lengths of small bowel is limited to a few centimetres, and division or occlusion of several consecutive vasa recta may produce segmental, focal ischaemia. Division of more proximal vessels allows flow to continue in the arcades for moderately long lengths of intestine.
Superior mesenteric artery
The superior mesenteric artery originates from the aorta 1 cm below the coeliac trunk, at the level of the intervertebral disk between the first and second lumbar vertebrae (Figs 66.9, 66.10). The angle of its origin from the aorta is acute, which makes cannulation via the transfemoral route somewhat difficult. The artery runs inferiorly and anteriorly, anterior to the uncinate process of the pancreas and the third part of the duodenum, and posterior to the splenic vein and the body of the pancreas. The left renal vein lies behind it and separates it from the aorta (Fig. 66.11). The artery crosses anterior to the inferior vena cava, right ureter and right psoas major as it descends in the root of the small bowel mesentery. Its calibre progressively decreases as successive branches are given off to loops of jejunum and ileum, and its terminal branch anastomoses with the ileocolic artery.
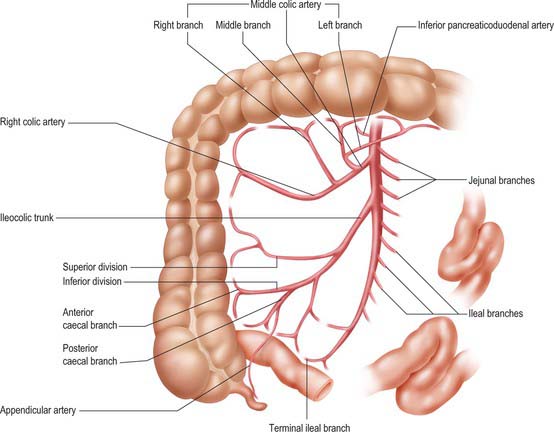
Fig. 66.9 The superior mesenteric artery and its branches. The outlines of representative ileal and jejunal loops, appendix, caecum, ascending and transverse colon are shown for reference. Only the origin of jejunal and ileal branches are shown. For details of arcades see Fig. 66.8.
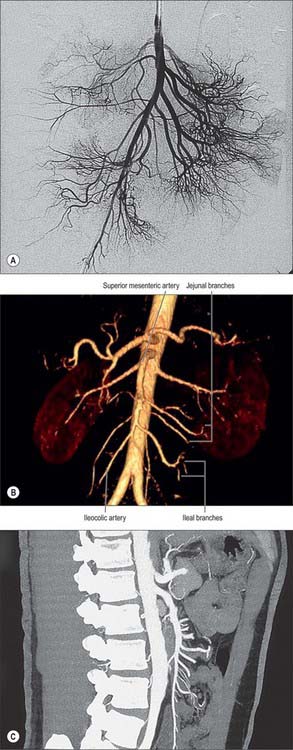
There are usually five to ten jejunal branches which arise from the left side of the upper portion of the superior mesenteric artery (Figs 66.9, 66.10). They are distributed to the jejunum as a series of short arcades which form a single (occasionally double) tier of anastomotic arcs before giving rise to multiple vasa recta (Fig. 66.8). These vessels run almost parallel in the mesentery and are distributed alternately to opposite aspects of its wall where the two series of vessels form distinct ‘leaves’ within the mesentery. Small twigs supply regional lymph nodes and other structures in the mesentery.
Ileal branches are more numerous than the jejunal branches but smaller in calibre. They arise from the left and anterior aspects of the superior mesenteric artery. The length of the mesentery is greater in the ileum and the branches form three, four or sometimes five tiers of arcs within the mesentery before giving rise to multiple vasa recta that run directly towards the ileal wall. The ileal branches run parallel in the mesentery and are distributed to alternate aspects of the ileum. They are longer and smaller than similar jejunal vessels, particularly in the distal ileum, and do not form such definite parallel ‘leaves’ of vessels. The vascular supply in the last loop of the terminal ileum is limited. There are usually only two separate arcades, peri-serosal and in the mid-zone of the mesentery. They each receive a contribution from the ileal branch of the ileocolic artery and the last ileal branch of the superior mesenteric artery and are often larger in calibre than the mid-ileal vessels. Other than this connection, few if any vessels connect the ileocolic and superior mesenteric arteries, which makes surgical exposure of the ileocolic artery up to its origin relatively simple.
Veins
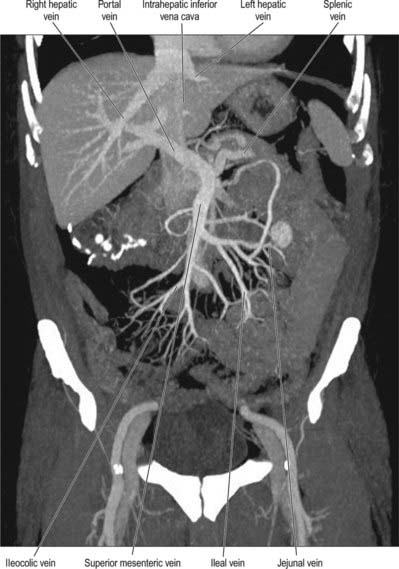
Superior mesenteric vein
The superior mesenteric vein drains the small intestine, caecum, ascending and transverse parts of the colon (Fig. 66.12; see Fig. 60.8). It is formed in the right lower mesentery of the small bowel by the union of tributaries from the terminal ileum, caecum and vermiform appendix. It ascends in the mesentery to the right of the superior mesenteric artery, passes anterior to the right ureter, inferior vena cava, third part of the duodenum and uncinate process of the pancreas, and finally joins the splenic vein behind the neck of the pancreas to form the portal vein.
INNERVATION
The ileum and jejunum are innervated by parasympathetic and sympathetic fibres via the superior mesenteric plexus (see Figs 60.2–60.4). Preganglionic sympathetic axons originate from neurones in the intermediolateral grey matter of the mid-thoracic spinal segments and travel in the greater and lesser splanchnic nerves to the superior mesenteric ganglion where they synapse. Postganglionic axons accompany the superior mesenteric artery into the mesentery and are distributed along branches of the artery. The sympathetic nerves are vasoconstrictor to the vasculature and inhibitory to the musculature of the jejunum and ileum.
MICROSTRUCTURE
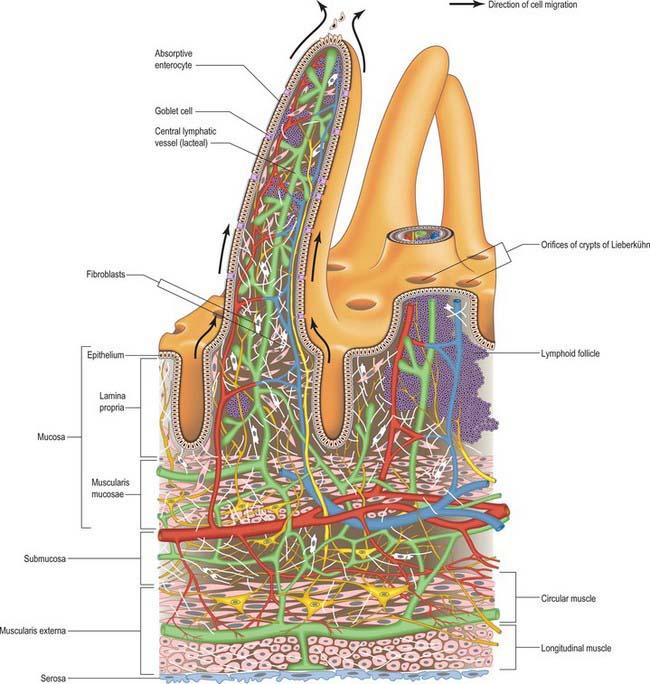
The intestinal wall is composed of mucosa, submucosa, muscularis externa and serosa or adventitia. The mucosa is thick and very vascular in the proximal small intestine, but thinner and less vascular distally. In places it is ridged by the underlying submucosa to form circular folds, plicae circulares, which protrude into the lumen; mucosal finger- or leaf-like intestinal villi cover the whole surface (Fig. 66.13). There are numerous simple tubular intestinal glands or crypts between the bases of the villi, and additional submucosal glands in the duodenum. The submucosa contains aggregates of lymphoid tissue, most numerous in the ileum.
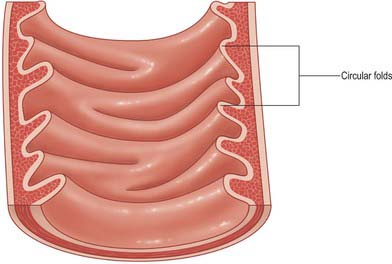
Fig. 66.13 Internal aspect of a representative sample of the proximal jejunum, showing circular folds.
Circular folds
Other than in the first part of the duodenum, large crescentic folds of mucosa project into the lumen of the small intestine, lying either transversely or slightly obliquely to its long axis (Fig. 66.14). Unlike gastric folds they are not lost during physiological distension of the intestine. Most extend round half or two-thirds of the luminal circumference, some are complete circles, some bifurcate and join adjacent folds, yet others are spiral but extend only once (occasionally two or three times) round the lumen. Larger folds are up to 8 mm deep at their broadest, but most are smaller than this, and larger folds often alternate with smaller ones. Folds begin to appear 2.5–5 cm beyond the pylorus. Distal to the major duodenal papilla they are large and close together, as they also are in the proximal half of the jejunum. From here to midway along the ileum they diminish, and they disappear almost completely in the distal ileum, which accounts for the thinness of this part of the intestinal wall. The circular folds slow the passage of the intestinal contents and increase the absorptive surface. They are visible in contrast and occasionally plain radiographs.
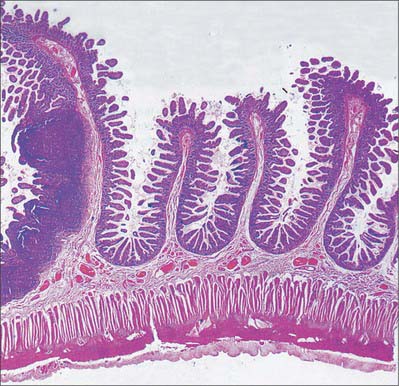
Intestinal villi
Intestinal villi are highly vascular projections of the mucosal surface, just visible to the naked eye (Figs 66.15–66.17). They cover the entire intestinal mucosa, increase the surface area of the lumen about eight-fold, and give it a velvety texture. Villi are large and numerous in the duodenum and jejunum, and smaller and fewer in the ileum. In the first part of the duodenum they appear as broad ridges, become tall and foliate in the distal duodenum and proximal jejunum, and then gradually shorten to a finger-like form in the distal jejunum and ileum. Villi vary in density from 10 to 40 per square millimetre and from 0.5–1.0 mm in height (Fig. 66.15). Infoldings of the mucosa dip down from the base of the villi for a short distance into the lamina propria. These crypts (of Lieberkühn), are most prominent in the proximal small intestine (Fig. 66.16).
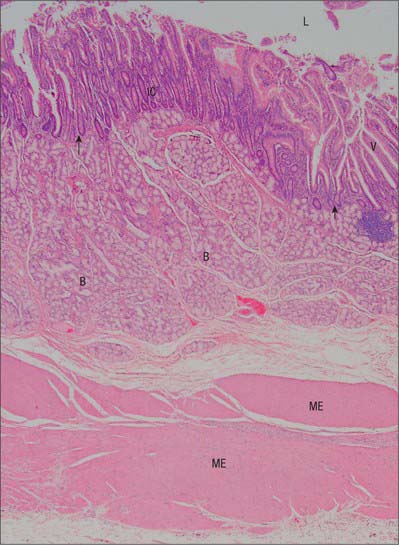
Mucosa
The mucosa consists of epithelium, lamina propria and muscularis mucosae.
Epithelium
A single-layered epithelium covers the intestinal villi (Figs 66.15, 66.16) and also lines the intestinal glands (crypts) that open between the bases of villi. Two types of cell, enterocytes and goblet cells, cover the surfaces of the villi, whereas microfold cells (M cells) are restricted to the dome epithelium that covers localized accumulations of lymphoid tissues. These cell types are all in contact basally with a basal lamina to which they adhere, and all are derived from a common stem cell in the intestinal crypts.
The apical cell surface is resistant to protease attack because microvilli possess a specialized glycoprotein-rich surface coat (glycocalyx) which, with an overlying layer of mucus, protects the epithelium against pancreatic enzymes in the intestinal lumen. The cell coat also contains a number of digestive enzymes as integral membrane proteins. These include enzymes that degrade disaccharides and oligopeptides prior to absorption. Further details of the structure of microvilli are given on page 18).
Microfold cells are present where the epithelium covers lymphoid aggregates in the intestinal wall (p. 74). They are cuboidal or flattened in shape and have long, widely spaced microfolds rather than microvilli on their apical surfaces. They sample luminal antigens by endocytosis, and transport antigen to lymphocytes lying within intercellular pockets formed by deep invaginations of the M-cell basolateral plasma membranes. See page 78 for details of antigen processing and presentation.
Intestinal glands or crypts
Intestinal glands or crypts (of Lieberkühn) are tubular pits that open into the lumen throughout the intestinal mucosa via small circular apertures between the bases of the villi (Figs 66.14–66.16). Their thin walls are composed of columnar enterocytes supplemented by mucous cells, Paneth cells, stem cells and neuroendocrine cells. They are separated by a basement membrane from a rich capillary plexus within the lamina propria.
Paneth cells are numerous in the deeper parts of the intestinal crypts, particularly in the duodenum. They are rich in zinc and contain large acidophilic granules that stain strongly with eosin or phosphotungstic haematoxylin. Paneth cells secrete lysozyme, a highly specific antibacterial enzyme, and other defensive proteins (defensins) such as tumour necrosis factor alpha (TNF-α), which protect the intestinal luminal surface.
Several types of neuroendocrine cell are scattered among the walls of the intestinal crypts, and less commonly over the villi. They secrete bioactive peptides, such as gastrin, cholecystokinin and secretin, basally into the surrounding lamina propria. Crypt neuroendocrine cells are derived from stem cells, which also give rise to enterocytes and other epithelial elements. For further details of the dispersed neuroendocrine system, see page Chapter 60.
Lamina propria
The lamina propria is composed of connective tissue and provides mechanical support for the epithelium. It has a rich vascular plexus, receives nutrients absorbed by the enterocytes, and forms the cores of the villi. It contains lymphoid tissue, fibroblasts and connective tissue extracellular matrix fibres, smooth muscle cells, eosinophils, macrophages, mast cells, capillaries, lymphatic vessels and unmyelinated nerve fibres. Plasma cells are numerous. Lymphocytes may be clustered in solitary or aggregated follicles, some of which extend through the muscularis mucosae into the submucosa.
Submucosa
Submucosal (Brunner’s) glands
Submucosal glands are limited to the submucosa of the duodenum and often referred to as ‘Brunner’s glands’ (see Fig. 66.15). Their ducts traverse the muscularis mucosae to enter the bases of the mucosal crypts. They are largest and most numerous near the pylorus and form an almost complete layer in the proximal half of the descending duodenum, thereafter they gradually diminish in number and disappear at the duodenojejunal junction. They are small, branched tubuloacinar glands: each has several secretory acini lined by low columnar epithelial cells that produce an alkaline (pH 9) mucoid secretion which effectively neutralizes acidic chyme from the stomach. Many neuroendocrine cells are present among the acinar cells.
Didierlaurent A, Simonet M, Sira J-C. Innate and acquired plasticity of the intestinal immune system. Cell Mol Life Sci. 2006;62:1285-1287.
Jackson JE. Vascular anatomy of the gastrointestinal tract. In: Butler P, Mitchell AWM, Ellis H. Applied Radiological Anatomy. Cambridge: Cambridge University Press, 1999.
Kadir S. Atlas of Normal and Variant Angiographic Anatomy. Philadelphia: WB Saunders, 1991.
Kraehenbuhl JP, Neutra MR. Molecular and cellular basis of immune protection of mucosal surfaces. Physiol Rev. 1992;72:853-879.
Underhill BML. Intestinal length in man. Br Med J. 1955;2:1243-1246.
[/level-membership-for-basic-science-category][not-level-membership-for-basic-science-category]
CHAPTER 66 Small intestine
DUODENUM
The adult duodenum is 20–25 cm long and is the shortest, widest and most predictably placed part of the small intestine. It is only partially covered by peritoneum, although the extent of the peritoneal covering varies along its length: the proximal 2.5 cm is intraperitoneal, and the remainder is retroperitoneal. The duodenum forms an elongated ‘C’ that lies between the level of the first and third lumbar vertebrae in the supine position. The lower ‘limb’ of the C extends further to the left of the midline than the upper limb. The head and uncinate process of the pancreas lie within the concavity of the duodenum which is ‘draped’ over the prominence formed from the lumbar spine, the duodenum therefore bends in an antero-posterior direction as well as following the form of a ‘C’. The duodenum lies entirely above the level of the umbilicus and is described as having four parts (Figs 66.1, 66.2).
FIRST (SUPERIOR) PART
The first, and most mobile, part of the duodenum is about 5 cm long. It starts as a continuation of the duodenal end of the pylorus and ends at the superior duodenal flexure. Peritoneum covers the anterior and superior part of its posterior aspect close to the pylorus, where the duodenum forms part of the anterior wall of the epiploic foramen. Here the lesser omentum is attached to its upper border and the greater omentum to its lower border. The first 2 or 3 cm have a bland internal mucosal appearance and readily distend on insufflation during endoscopy. This part is frequently referred to as the duodenal ‘cap’. It has a triangular, homogeneous appearance during contrast radiology, shows the same pattern of internal rugae as the pylorus, and is often visible on plain radiographs of the abdomen as an isolated triangular gas shadow to the right of the first or second lumbar vertebra. The duodenum next passes superiorly, posteriorly and laterally for 5 cm before curving sharply inferiorly into the superior duodenal flexure: it rapidly becomes more retroperitoneal during this part of its course, until peritoneum only covers its anterior aspect. From the end of the duodenal cap, its internal appearance is characterized by extensive, deep mucosal folds that involve up to half of the circumference of the lumen. Even during endoscopic insufflation, these folds are pronounced (Fig. 66.3) and they are readily seen on contrast radiographs. The section from the duodenal cap to the superior duodenal flexure lies posterior and inferior to the quadrate lobe of the liver.
SECOND (DESCENDING) PART
The second part lies anterior to the hilum of the right kidney, the right renal vessels, the edge of the inferior vena cava and the right psoas major. The head of the pancreas and the common bile duct are medial and the hepatic flexure is above and lateral. A small part of the pancreatic head is sometimes embedded in the medial duodenal wall, and pancreatic ‘rests’ in the duodenal wall may produce small filling defects on contrast radiology. The internal appearance is similar to that of the distal portion of the first part of the duodenum, with pronounced mucosal folds (Fig. 66.3). The common bile duct and pancreatic duct enter the medial wall obliquely and usually unite to form the common hepatopancreatic ampulla. The narrow distal end opens on the summit of the major duodenal papilla (ampulla of Vater), which is situated on the posteromedial wall of the second part, 8–10 cm distal to the pylorus. There are variants of the ductal anatomy. The most common is a second, accessory pancreatic duct which may open 2 cm above the major papilla on a minor duodenal papilla. Peptic ulceration of the second part of the duodenum is less common than that of the first part, and tends to occur on the anterior or lateral wall.
FOURTH (ASCENDING) PART
The fourth part of the duodenum is 2.5 cm long. It starts just to the left of the aorta, runs superiorly and laterally to the level of the upper border of the second lumbar vertebra, then turns sharply anteroinferiorly at the duodenojejunal flexure to become continuous with the jejunum. The main trunk of the inferior mesenteric vein lies either posterior to the duodenojejunal flexure or beneath the adjacent peritoneal fold. (The duodenojejunal flexure is a useful landmark to locate the vein radiologically or surgically.) The aorta, left sympathetic trunk, left psoas major, left renal and left gonadal vessels are all posterior relations, the left kidney and left ureter are posterolateral, and the upper part of the root of the small bowel mesentery, the left lateral transverse mesocolon and transverse colon, which separate it from the stomach, are anterior. The peritoneum of the root of the small bowel mesentery continues over its anterior surface. The lower border of the body of the pancreas is superior.
At its left lateral end, the fourth part becomes progressively covered in peritoneum on its superior and inferior surfaces, such that it is suspended from the retroperitoneum by a double fold of peritoneum, the ligament of Treitz, at the start of the duodenojejunal flexure. The ligament of Treitz may contain a small slip of muscle, the suspensory muscle of the duodenum. When present, the suspensory muscle contains skeletal muscle fibres that run from the left crus of the diaphragm to connective tissue around the coeliac axis, and smooth muscle fibres that run from the coeliac axis: its function is unknown. It is an important landmark in the radiological diagnosis of incomplete rotation and malrotation of the small intestine (see Ch. 73). The ligament of Treitz does not form part of the duodenal mesentery because the vascular supply to the fourth part of the duodenum continues to enter its wall from the posteromedial aspect.
VASCULAR SUPPLY AND LYMPHATIC DRAINAGE
Arteries
The main vessels supplying the duodenum are the superior and inferior pancreaticoduodenal arteries. The first and second parts also receive contributions from several sources including the right gastric, supraduodenal, right gastroepiploic, hepatic and gastroduodenal arteries. Branches of the superior pancreaticoduodenal artery may contribute to the supply of the pyloric canal, with a minor degree of anastomosis to the gastric arteries in the muscular layer across the pyloroduodenal junction (Fig. 66.4).
Superior pancreaticoduodenal arteries
The superior pancreaticoduodenal artery is usually double. The anterior artery is a terminal branch of the gastroduodenal artery and descends in the anterior groove between the second part of the duodenum and the head of the pancreas. It supplies branches to the first and second parts of the duodenum and to the head of the pancreas, and anastomoses with the anterior division of the inferior pancreaticoduodenal artery. The posterior artery is usually a separate branch of the gastroduodenal artery and is given off at the upper border of the first part of the duodenum. It descends to the right, anterior to the portal vein and common bile duct as the latter lies behind the first part of the duodenum. It then runs behind the head of the pancreas, crosses posterior to the common bile duct (which is embedded in the head of the pancreas), enters the duodenal wall and anastomoses with the posterior division of the inferior pancreaticoduodenal artery. The posterior artery supplies branches to the head of the pancreas, the first and second parts of the duodenum, and several branches to the lowest part of the common bile duct.
