9 Sensory Receptors and the Peripheral Nervous System
Receptors Encode the Nature, Location, Intensity, and Duration of Stimuli
The job of sensory receptors collectively is to produce electrical signals that represent all relevant aspects of stimuli—what kind of stimulus, where it is, how intense, when it starts and stops. Sometimes a single receptor can do all of this, but often one or more populations of receptors are required.
Each Sensory Receptor Has an Adequate Stimulus, Allowing It to Encode the Nature of a Stimulus
Sensory receptors transduce (“lead across”) some aspect of the external or internal environment into a graded electrical signal (a receptor potential). Each receptor is more sensitive to one kind of stimulus, called its adequate stimulus, than to others. Hence, there are chemoreceptors, photoreceptors, thermoreceptors, and mechanoreceptors, and the identity of the particular receptors responding to a stimulus provides some initial information about the nature of that stimulus. Individual types of receptors within these broad classes are usually more finely tuned to particular aspects of a stimulus category, providing even more information about the nature of a stimulus. For example, although all the mechanoreceptors of the inner ear are very similar to each other, some are set up to respond best to sound vibrations, others to the position of the head (see Chapter 14).
Receptor Potentials Encode the Intensity and Duration of Stimuli
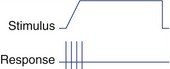
To a first approximation, receptors encode the intensity and duration of stimuli by the size and duration of the receptor potentials they produce (Fig. 9-1). There’s actually a little more to it than this, though, because in some systems increasing intensity is signaled by recruiting additional, less sensitive, receptors (e.g., rods for dim light and cones for bright light). In addition, some receptors signal only the beginning and end of a stimulus and do not respond to maintained stimuli.
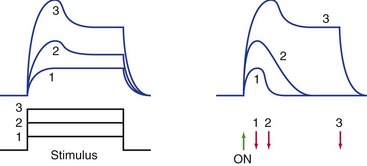
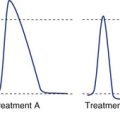
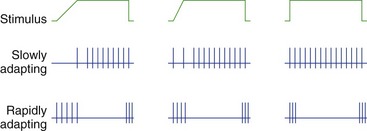
Most Sensory Receptors Adapt to Maintained Stimuli, Some More Rapidly Than Others
Most receptors, unlike the two shown in Fig. 9-2, are actually somewhere between the extremes of slowly and rapidly adapting. The response may be exaggerated at the beginning (or end) of a stimulus, but maintained to some extent throughout the stimulus (see Fig. 9-1).
Sensory Receptors All Share Some Organizational Features
Sensory Receptors Use Ionotropic and Metabotropic Mechanisms to Produce Receptor Potentials
The transduction mechanisms used by sensory receptors are gratifyingly similar to the mechanisms used in the production of postsynaptic potentials (Fig. 9-3). Some are depolarizing, others hyperpolarizing; some use direct alteration of ion channels, others use G protein–coupled mechanisms (Table 9-1). Many of the receptor molecules used by sensory receptors are actually closely related to postsynaptic receptor molecules, but are simply set up so that they respond to a stimulus rather than to a neurotransmitter.
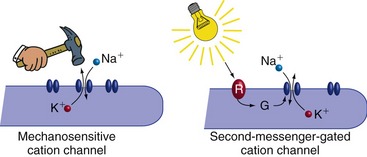
Table 9-1 Transduction mechanisms used by different kinds of sensory receptors
| Stimulus-Gated Channels | G Protein–Coupled Mechanisms |
|---|---|
| Most somatosensory receptors | Some pain receptors Photoreceptors |
| Hair cells (CN VIII) | Olfactory receptors |
| Some taste receptors | Some taste receptors |
| Some visceral receptors | Some visceral receptors |
All Sensory Receptors Produce Receptor Potentials, but Some Do Not Produce Action Potentials
Receptor potentials are not propagated actively; rather, like postsynaptic potentials, they decay over a short distance. Therefore, receptors that signal over long distances must generate action potentials as well as receptor potentials (Fig. 9-4). An example is a receptor that signals something touching your big toe. It produces a depolarizing receptor potential in response to touch, but the receptor potential itself dies out near the receptive ending. But action potentials are initiated at a nearby trigger zone and get conducted all the way into the CNS. In mammals, receptors with long axons convey information about somatic sensation (touch, pain, etc.), visceral sensation, and smell, and all produce depolarizing receptor potentials.
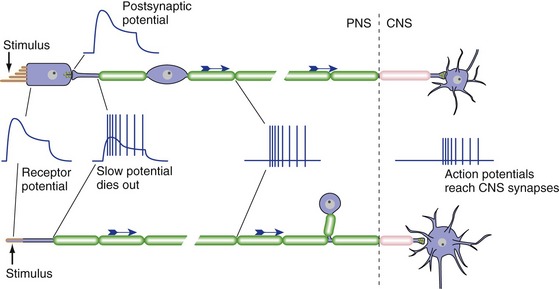
In contrast, receptors that signal over short distances (less than a mm or so) don’t need to produce action potentials. Instead, they synapse on the peripheral processes of primary afferent neurons, whose cell bodies lie in peripheral ganglia. The receptor potential changes the rate at which the receptor releases transmitter, and this in turn changes the rate at which the primary afferent sends action potentials into the CNS. A receptor with a short axon or no axon can depolarize (and release more transmitter) or hyperpolarize (and release less transmitter) in response to a stimulus; some can do both, depending on the specifics of the stimulus. Examples of short receptors are taste receptor cells, photoreceptors, and the mechanoreceptive hair cells of the inner ear.
Somatosensory Receptors Detect Mechanical, Chemical, or Thermal Changes
Receptors in Muscles and Joints Detect Muscle Status and Limb Position
Muscle spindles are receptor organs composed of small muscle fibers (called intrafusal fibers, meaning “inside the spindle”) enclosed in a spindle-shaped capsule (THB6 Figure 9-14, p. 216). The spindles are embedded in skeletal muscles and oriented so that they are stretched by anything that stretches the muscle. Sensory endings attached to the intrafusal fibers produce depolarizing receptor potentials when the spindles are stretched (Fig. 9-5). The ends of intrafusal fibers are contractile and receive inputs from small (gamma) motor neurons. Contraction of this part of an intrafusal fiber does not contribute significantly to the strength of a muscle. Rather, it regulates the length of the central, stretch-sensitive portion of the intrafusal fiber and thereby regulates its sensitivity to externally applied stretch (THB6 Figure 9-15, p. 217).
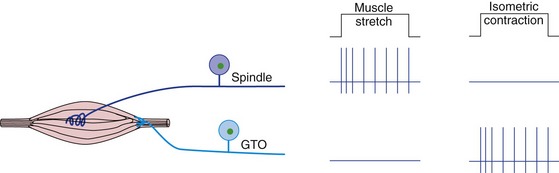
Golgi tendon organs are networks of sensory endings interspersed among the collagen fibers of tendons (THB6 Figure 9-16, p. 218). Tension in a tendon compresses the sensory endings and causes depolarizing receptor potentials. Passive muscle stretch does not generate much tension in a tendon, but muscle contraction against a load does (see Fig. 9-5).
Visceral Structures Contain a Variety of Receptive Endings
Visceral structures also receive a wealth of receptive endings, but much less is known about them than about somatosensory and other receptors. Collectively they form the afferent components of a vast network of connections mediating the homeostatic and drive-related behaviors discussed in Chapter 23.
Peripheral Nerves Convey Information to and from the CNS
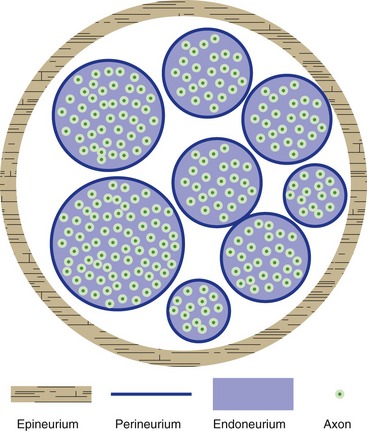
The dura mater surrounding the CNS continues as the epineurium of peripheral nerves (Fig. 9-6; see THB6 Figure 9-19, p. 222). This is a substantial covering around each nerve, conferring considerable mechanical strength. The arachnoid continues as the perineurium covering individual nerve fascicles. Just as in the case of the arachnoid, perineurial cells are interconnected by tight junctions and form a diffusion barrier between the inside and the outside of a nerve fascicle, part of a continuation of the blood-brain barrier system as a blood-nerve barrier (capillaries inside nerve fascicles are zipped up by tight junctions, helping to complete the barrier). The endoneurium is the loose background connective tissue within nerve fascicles.
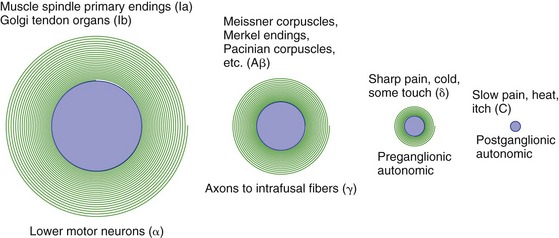
The Diameter of a Nerve Fiber Is Correlated with Its Function
Large-diameter axons have large cell bodies and thick myelin sheaths, and they conduct action potentials rapidly. Small-diameter axons have smaller cell bodies, have thin myelin sheaths or are unmyelinated, and conduct action potentials slowly. Large-diameter sensory axons end peripherally in muscle spindles, Golgi tendon organs, and joint receptors. The sensory axons that end peripherally as the cutaneous receptors used for discriminative tactile sense (precise judgments of shape and position) are not quite as large. Small-diameter axons end peripherally as receptors for pain, temperature, simple sensation of touch, and assorted visceral receptors. Major categories of somatic afferent fibers in peripheral nerves, and their relative sizes, are indicated in Fig. 9-7 (see THB6 Table 9-3, p. 224, for more details).