[level-membership-for-basic-science-category]muscle tissue
Specific terms have been introduced to identify particular parts of muscle cells (which are also known as muscle fibres). The muscle cell membrane is known as the sarcolemma, the cytoplasm as the sarcoplasm and the smooth endoplasmic reticulum as the sarcoplasmic reticulum.
Types of muscle tissue
There are three types of muscle tissue classified by location and the arrangement of the contractile myofilaments in the muscle cells:
■ skeletal muscle contraction is involved, mainly, in moving the skeleton
■ cardiac muscle contraction moves blood through the heart
■ smooth muscle in the walls of many tubes and hollow organs (e.g. in some blood vessels and the urinary bladder) controls the flow of substances along or out of such structures.
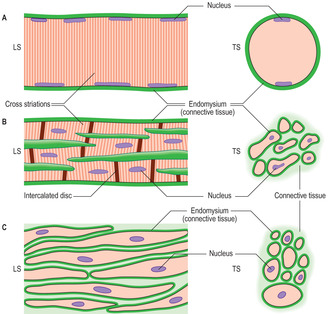
Skeletal and cardiac muscle cells are referred to as striped or striated muscle as the contractile myofilaments are arranged in a repeating pattern in the sarcoplasm and this is manifested as transverse dark and light striations across the length of the muscle cells (Figs 5.1A,B and 5.2). The precise arrangement of the contractile myofilaments are revealed only by using an electron microscope (see below). Although smooth muscle cells contain actin and myosin myofilaments, they are not arranged in a regular pattern and smooth muscle cells do not display striations (Fig. 5.1C).
Other cell types with the ability to contract have also been detected using techniques which identify contractile proteins in their cytoplasm. These types include:
■ myoepithelial cells, which surround acini and ducts of some exocrine glands; their contraction aids the expulsion of secretions
■ myofibroblasts, which occur in many regions of connective tissue; they aid repair by secreting collagen and undergoing contraction during scar formation
■ pericytes, which surround capillaries; after injury they undergo mitosis and replace damaged fibroblasts and smooth muscle cells.
Muscle cells and connective tissue
Confusingly, muscle cells are also referred to as muscle fibres, probably because some are long, thin structures which measure several centimetres in length. (In contrast, connective tissue fibres, although long and thin, are protein molecules, not cells.)
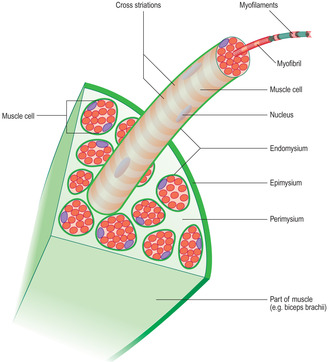
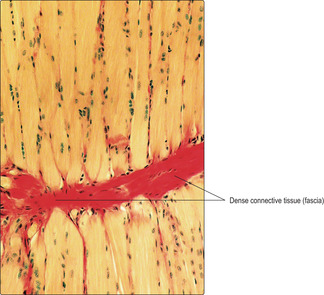
The connective tissue associated with muscle cells consists of three covering layers (Figs 5.1 and 5.3). Each muscle cell is covered by, and bound to, fine connective tissue fibres which form an external (basal) lamina known as an endomysium. Several muscle cells are held together as bundles by connective tissue fibres known as the perimysium. The connective tissue covering a muscle itself, e.g. the biceps brachii, is called the epimysium; this layer is also known as deep fascia.
Contraction of a muscle, such as biceps brachii, involves the contractile proteins in individual muscle cells moving and shortening their overall length. This movement generates a force of contraction and it is transmitted to the sarcolemma and the external lamina. The laminae around adjacent muscle cells are connected to each other and to the other layers of connective tissue. In this way the force of muscle cell contraction is transmitted to all the connective tissue layers. Such contractile forces from muscle cells, applied to connective tissue fibres, is then transferred to other structures such as bone e.g. the humerus or soft tissue, e.g. the eyeball.
It is important to appreciate that when muscles stop contracting they passively relax and the contractile proteins resume their original position but this does not exert a force. Muscles can only contract (pull), never push!
Skeletal muscle
As its name suggests this type of muscle is associated with the skeletal system, and it is responsible for movements of bones and joints. Skeletal muscle is also known as voluntary muscle as its contraction is usually under conscious control via its nerve supply. Skeletal muscle gives characteristic form to the body’s contour (along with adipose tissue). Although gross anatomy is beyond the scope of this book, it is important to appreciate that the attachment sites of a skeletal muscle to the skeleton, and the position of the muscle in relation to a joint, will determine the line of pull of the muscle and the type of movement that occurs at the joint as the muscle contracts.
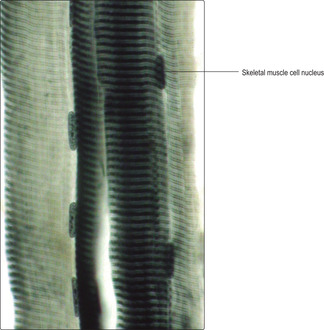
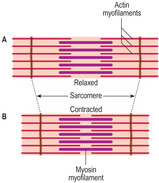
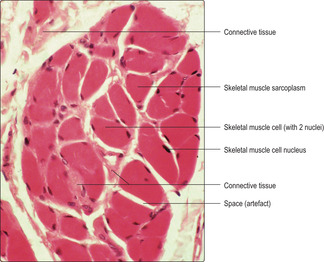
Skeletal muscle cells are multinucleate, long and cylindrical in shape. They form by fusion of several cells (myoblasts) during development in utero. The nuclei of skeletal muscle cells are at the periphery of the sarcoplasm (Figs 5.1A and 5.2), a feature which allows them to be distinguished from other types of muscle cell. The contractile myofilaments (actin and myosin) within muscle cells are arranged as bundles (myofibrils) with their long axes along the length of the cells (Fig. 5.3) and each cell contains hundreds of myofibrils. The contractile proteins in the myofibrils overlap each other (Fig. 5.4A) and form a functional unit (a sarcomere). Aligned in this way the myofibrils refract light differently, hence the striped appearance (Fig. 5.2). The dark bands are known as A bands and the I bands are light.
In histological sections of skeletal muscle cells which have been sectioned along their length, transverse striations, reflecting the arrangement of the contractile proteins, are apparent (Fig. 5.2). If skeletal muscle cells are not sectioned along their length the striations are not apparent, but it is possible to recognise the type of muscle cell by the peripheral position of the nuclei (Figs 5.1A and 5.5). Numerous blood capillaries (Fig. 5.6) can be demonstrated in the connective tissue layers around muscle cells. The capillaries ensure a good blood supply, which provides the oxygen and nutrients necessary for muscle contraction.
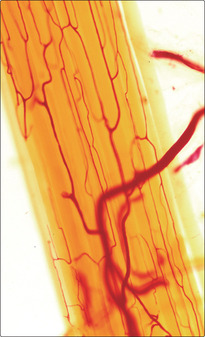
Many skeletal muscles are anchored to bone by tendons (Chapter 4). The collagen fibres covering the muscle cells are continuous with the collagen of the tendon. In turn, bundles of collagen, known as Sharpey’s fibres, pass into the bone and anchor the tendon. Tendons take on a variety of forms, such that the attachment may be focal or broad. In some muscles the connective tissue surrounding the muscle cells attaches to a collagen (fascial) layer (Fig. 5.7), rather than to bone.
Variation occurs in the size of muscle cells, the number of mitochondria they contain and the enzymes present, as well as their content of myoglobin. Myoglobin, an oxygen-carrying protein, resembles haemoglobin and gives muscle a reddish colour. Such red muscle cells are predominant in muscles in which sustained contraction is required, e.g. to maintain posture. In contrast, in muscles where fast contraction and spurts of activity occur myoglobin is sparse and the muscles appear whitish to the naked eye.
Ultrastructure of skeletal muscle and contraction
Contractile proteins occupy much of the sarcoplasm of muscle cells, and the thick myosin and thin actin myofilaments can be identified in cells sectioned longitudinally and transversely (Figs 5.8 and 5.9 respectively). Numerous mitochondria (Fig. 5.9) are present in muscle cells and myoglobin and glycogen may also be present; all aid in providing energy for contraction. An extensive membranous meshwork connecting the sarcolemma and the sarcoplasmic reticulum (the t tubule system) is present (Fig. 5.9). This meshwork has a role in moving calcium ions into the sarcoplasm, which is a process that is an essential trigger for muscle contraction.

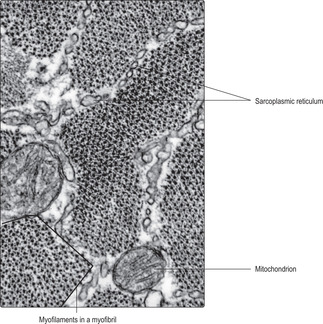
The functional unit of the skeletal muscle cell is known as a sarcomere (Fig. 5.4) and it consists of thin (actin) and thick (myosin) myofilaments and other proteins which anchor them in parallel arrays and to the sarcolemma. Contraction involves the parallel actin and myosin proteins sliding along each other (Fig. 5.4), thus shortening the cell overall. This is known as the sliding filament mechanism of contraction and is an energy-dependent process.
Cardiac muscle
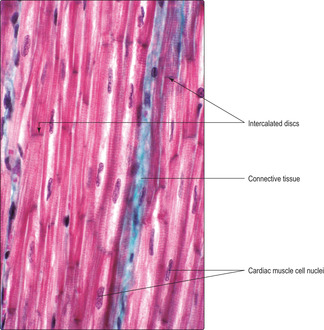
This type of muscle, like skeletal muscle, appears striated (Figs 5.1B and 5.10) owing to the arrangement of the actin and myosin proteins in the sarcoplasm. Unlike skeletal muscle, cardiac muscle contraction is not under conscious control. Cardiac muscle cells are cylindrical and much shorter than skeletal muscle cells. Each cardiac muscle cell has a single, centrally placed nucleus and some cells branch (Fig. 5.1B). Specialised cell junctions (Chapter 2) link cardiac muscle cells end to end and these regions, called intercalated discs, are visible in histological sections (Fig. 5.10). Desmosomes and zonulae adherens anchor the muscle cells together at intercalated discs. In addition, there are gap junctions at these discs which ensure the excitation stimulating contraction spreads rapidly between muscle cells and results in their synchronised contraction. The mechanism of contraction is similar to that of skeletal muscle. The connective tissue associated with cardiac muscle (Fig. 5.10) is arranged in layers similar to those of skeletal muscle and it transmits the forces of the muscle contraction. The arrangement of connective tissue fibres forms a fibrous ‘skeleton’ for the heart. This arrangement helps to ensure that the contraction of heart muscle cells constricts the chambers of the heart in synchrony and it also anchors the heart valves and aids their function (Chapter 10).
Smooth muscle
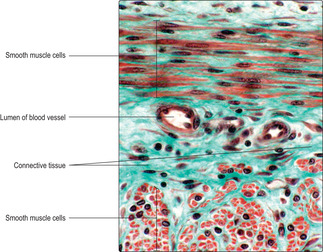
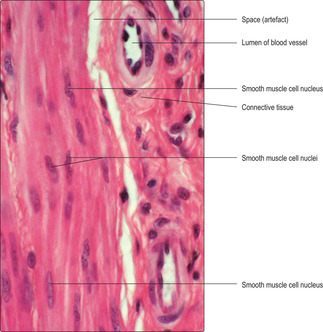
Smooth muscle cells are fusiform in shape and tapered at each end with a centrally placed nucleus (Fig. 5.1C). Smooth muscle cells are usually grouped together and form sheets or bundles, and their appearance in histological sections depends on the angle at which they are sectioned (Figs 5.1C and 5.11) and their state of contraction (Fig. 5.12). Contraction of smooth muscle cells is not under conscious control and they are described as involuntary muscle cells. The contractile proteins in smooth muscle cells are arranged irregularly and some are linked with adherens-type junctions at intervals on the inner surface of the cell membrane. Each muscle cell is attached to an external lamina of connective tissue (endomysium) (Fig. 5.1C) and muscle cells are bundled together by layers of connective tissue (Fig. 5.11). Between some muscle cells the external lamina is incomplete and gap junctions link adjacent smooth muscle cells. The presence of gap junctions aids the spread of the excitatory stimulus for contraction and helps groups of cells to contract as a single unit. When smooth muscle cells contract they take on a corkscrew-like shape, which may be apparent as a change in shape of the nucleus (Fig. 5.12), and the force is transmitted via their external laminae to other surrounding connective tissue.
Muscle contraction and nerve supply
Skeletal muscle contraction is dependent on its nerve supply. Indeed, if the nerves supplying skeletal muscles are severed the muscle cells do not contract and they waste away (atrophy). Cardiac and smooth muscle cells have an intrinsic ability to contract in the absence of nerve supply. However, they are supplied by nerves which control their activity but they are not normally under conscious control. Further details of the nerve supply to muscles is given in Chapter 6.
Clinical note
Muscle hypertrophy, hyperplasia and repair Adult skeletal muscle is able to undergo hypertrophy: the muscle cells become larger as they respond to exercise by forming more contractile proteins. Although skeletal muscle cells are not able to undergo mitosis, if very minor damage occurs some repair may occur. This is due to a minor population of undifferentiated cells (known as satellite cells) in skeletal muscle which can differentiate into skeletal muscle cells.
Cardiac muscle cells can also increase in volume by hypertrophy, and this can occur in response to high blood pressure (hypertension). In contrast to skeletal muscle, there are no stem cells in cardiac muscle and so there is no replacement of dead cardiac muscle cells. After a heart attack (myocardial infarction), cardiac muscle cells die because of loss of their blood supply and they are not replaced. Fibroblasts in the connective tissue of the heart proliferate and secrete collagen and a fibrous scar takes the place of the dead muscle cells. The scar may alter the contraction of the normal heart muscle with harmful consequences.
Smooth muscle cells can undergo hypertrophy (increase in cell size) and hyperplasia (increase in cell number). These processes are particularly important in ensuring uterine muscle is able to function during pregnancy and childbirth.
Muscle tissue
■ skeletal, cardiac and smooth.
■ Myoepithelial cells, myofibroblasts and pericytes also have the ability to contract.
■ Intracytoplasmic proteins (mainly the myofilaments actin and myosin) in muscle cells (and the above cell types) are responsible for contraction.
■ Connective tissue adjacent to muscle cells transmits the force of contraction to other structures.
Skeletal muscle
■ Cells are multinucleate and nuclei are adjacent to the sarcolemma (muscle cell membrane).
■ Cells are long, cylindrical and display transverse striations reflecting the linear arrangement of the myofilaments in the sarcoplasm (cytoplasm).
■ Contraction moves bones at joints and/or stabilises joints.
■ Contraction is (mostly) under conscious control via the nerve supply.
Cardiac muscle
■ Cells are cylindrical, relatively short, and have a single, centrally placed nucleus.
■ Cells display transverse striations corresponding to the linear arrangement of myofilaments.
■ Cells have intercalated discs showing the end-to-end connection of these cells.
■ Contraction is under the (unconscious) control of nerves.
■ Contractions, transmitted to connective tissue in the heart, move blood through the heart in a synchronised manner.
Smooth muscle
■ Cells are fusiform in shape and a nucleus is in the centre of each cell.
■ Cells have few randomly arranged myofilaments.
■ Cells occur in bundles or layers around many tubes.
■ Contraction is under (unconscious) control of nerves.
[/level-membership-for-basic-science-category][not-level-membership-for-basic-science-category]muscle tissue
Specific terms have been introduced to identify particular parts of muscle cells (which are also known as muscle fibres). The muscle cell membrane is known as the sarcolemma, the cytoplasm as the sarcoplasm and the smooth endoplasmic reticulum as the sarcoplasmic reticulum.
Types of muscle tissue
There are three types of muscle tissue classified by location and the arrangement of the contractile myofilaments in the muscle cells:
■ skeletal muscle contraction is involved, mainly, in moving the skeleton
■ cardiac muscle contraction moves blood through the heart
■ smooth muscle in the walls of many tubes and hollow organs (e.g. in some blood vessels and the urinary bladder) controls the flow of substances along or out of such structures.
Skeletal and cardiac muscle cells are referred to as striped or striated muscle as the contractile myofilaments are arranged in a repeating pattern in the sarcoplasm and this is manifested as transverse dark and light striations across the length of the muscle cells (Figs 5.1A,B and 5.2). The precise arrangement of the contractile myofilaments are revealed only by using an electron microscope (see below). Although smooth muscle cells contain actin and myosin myofilaments, they are not arranged in a regular pattern and smooth muscle cells do not display striations (Fig. 5.1C).
Other cell types with the ability to contract have also been detected using techniques which identify contractile proteins in their cytoplasm. These types include:
■ myoepithelial cells, which surround acini and ducts of some exocrine glands; their contraction aids the expulsion of secretions
■ myofibroblasts, which occur in many regions of connective tissue; they aid repair by secreting collagen and undergoing contraction during scar formation
■ pericytes, which surround capillaries; after injury they undergo mitosis and replace damaged fibroblasts and smooth muscle cells.
Muscle cells and connective tissue
Confusingly, muscle cells are also referred to as muscle fibres, probably because some are long, thin structures which measure several centimetres in length. (In contrast, connective tissue fibres, although long and thin, are protein molecules, not cells.)
The connective tissue associated with muscle cells consists of three covering layers (Figs 5.1 and 5.3). Each muscle cell is covered by, and bound to, fine connective tissue fibres which form an external (basal) lamina known as an endomysium. Several muscle cells are held together as bundles by connective tissue fibres known as the perimysium. The connective tissue covering a muscle itself, e.g. the biceps brachii, is called the epimysium; this layer is also known as deep fascia.
Contraction of a muscle, such as biceps brachii, involves the contractile proteins in individual muscle cells moving and shortening their overall length. This movement generates a force of contraction and it is transmitted to the sarcolemma and the external lamina. The laminae around adjacent muscle cells are connected to each other and to the other layers of connective tissue. In this way the force of muscle cell contraction is transmitted to all the connective tissue layers. Such contractile forces from muscle cells, applied to connective tissue fibres, is then transferred to other structures such as bone e.g. the humerus or soft tissue, e.g. the eyeball.
It is important to appreciate that when muscles stop contracting they passively relax and the contractile proteins resume their original position but this does not exert a force. Muscles can only contract (pull), never push!
Skeletal muscle
As its name suggests this type of muscle is associated with the skeletal system, and it is responsible for movements of bones and joints. Skeletal muscle is also known as voluntary muscle as its contraction is usually under conscious control via its nerve supply. Skeletal muscle gives characteristic form to the body’s contour (along with adipose tissue). Although gross anatomy is beyond the scope of this book, it is important to appreciate that the attachment sites of a skeletal muscle to the skeleton, and the position of the muscle in relation to a joint, will determine the line of pull of the muscle and the type of movement that occurs at the joint as the muscle contracts.
Skeletal muscle cells are multinucleate, long and cylindrical in shape. They form by fusion of several cells (myoblasts) during development in utero. The nuclei of skeletal muscle cells are at the periphery of the sarcoplasm (Figs 5.1A and 5.2), a feature which allows them to be distinguished from other types of muscle cell. The contractile myofilaments (actin and myosin) within muscle cells are arranged as bundles (myofibrils) with their long axes along the length of the cells (Fig. 5.3) and each cell contains hundreds of myofibrils. The contractile proteins in the myofibrils overlap each other (Fig. 5.4A) and form a functional unit (a sarcomere). Aligned in this way the myofibrils refract light differently, hence the striped appearance (Fig. 5.2). The dark bands are known as A bands and the I bands are light.
Buy Membership for Basic Science Category to continue reading. Learn more here
[/not-level-membership-for-basic-science-category]
