CHAPTER 33 Pharynx
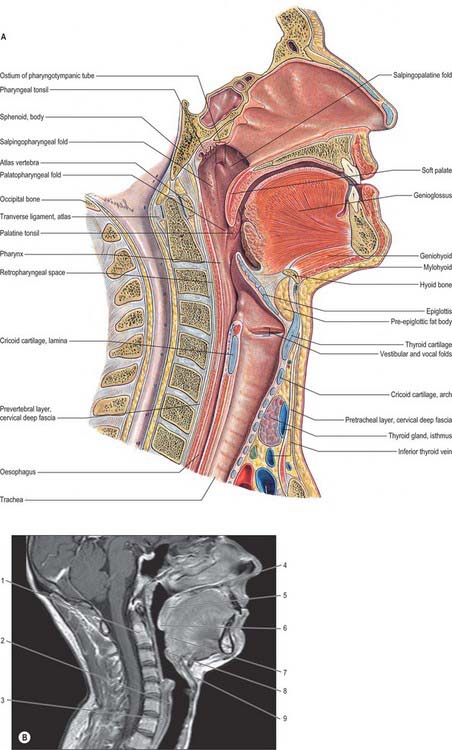
The pharynx is a 12–14 cm long musculomembranous tube shaped like an inverted cone. It extends from the cranial base to the lower border of the cricoid cartilage (the level of the sixth cervical vertebra), where it becomes continuous with the oesophagus. The width of the pharynx varies constantly because it is dependent on muscle tone, especially of the constrictors: at rest the pharyngo-oesophageal junction is closed as a result of tonic closure of the upper oesophageal sphincter, and during sleep muscle tone is low and the dimensions of the pharynx are markedly decreased (which may give rise to snoring and sleep apnoea). The pharynx is limited above by the posterior part of the body of the sphenoid and the basilar part of the occipital bone, and it is continuous with the oesophagus below. Behind, it is separated from the cervical part of the vertebral column and the prevertebral fascia which covers longus colli and longus capitis by loose connective tissue in the retropharyngeal space above and the retrovisceral space below.
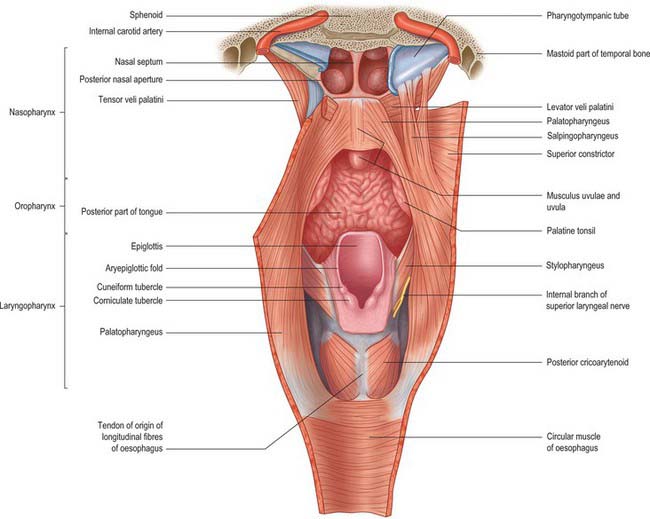
The pharynx lies behind, and communicates with, the nasal, oral and laryngeal cavities via the nasopharynx, oropharynx and laryngopharynx respectively (Figs 33.1, 33.2). Its lining mucosa is continuous with that lining the pharyngotympanic tubes, nasal cavity, mouth and larynx.
NASOPHARYNX
BOUNDARIES
The nasopharynx lies above the soft palate and behind the posterior nares, which allow free respiratory passage between the nasal cavities and the nasopharynx (Figs 33.1, 33.2). The nasal septum separates the two posterior nares, each of which measures approximately 25 mm vertically and 12 mm transversely. Just within these openings lie the posterior ends of the inferior and middle nasal conchae. The nasopharynx has a roof, a posterior wall, two lateral walls and a floor. These are rigid (except for the floor, which can be raised or lowered by the soft palate) and the cavity of the nasopharynx is therefore never obliterated by muscle action, unlike the cavities of the oro- and laryngopharynx. The nasal and oral parts of the pharynx communicate through the pharyngeal isthmus which lies between the posterior border of the soft palate and the posterior pharyngeal wall. Elevation of the soft palate and constriction of the palatopharyngeal sphincter close the isthmus during swallowing.
The lateral walls of the nasopharynx display a number of important surface features. On either side each receives the opening of the pharyngotympanic tube (also termed the auditory or Eustachian tube), situated 10–12 mm behind and a little below the level of the posterior end of the inferior nasal concha (Fig. 33.2). The tubal aperture is approximately triangular in shape, and is bounded above and behind by the tubal elevation which consists of mucosa overlying the protruding pharyngeal end of the cartilage of the pharyngotympanic tube. A vertical mucosal fold, the salpingopharyngeal fold, descends from the tubal elevation behind the aperture (Fig. 33.2) and covers salpingopharyngeus in the wall of the pharynx (see Fig. 33.4); a smaller salpingopalatine fold extends from the anterosuperior angle of the tubal elevation to the soft palate in front of the aperture. As levator veli palatini enters the soft palate it produces an elevation of the mucosa immediately around the tubal opening (see Fig. 33.4). A small variable mass of lymphoid tissue, the tubal tonsil, lies in the mucosa immediately behind the opening of the pharyngotympanic tube, and further behind the tubal elevation there is a variable depression in the lateral wall, the lateral pharyngeal recess (fossa of Rosenmüller). The floor of the nasopharynx is formed by the nasal, upper surface of the soft palate.
MICROSTRUCTURE OF NON-TONSILLAR NASOPHARYNX
The nasopharyngeal epithelium anteriorly is ciliated, pseudostratified respiratory in type with goblet cells (see Fig. 2.2). The ducts of mucosal and submucosal seromucous glands open onto its surface. Posteriorly, the respiratory epithelium changes to non-keratinized stratified squamous epithelium which continues into the oropharynx and laryngopharynx. The transitional zone between the two types of epithelium consists of columnar epithelium with short microvilli instead of cilia. Superiorly, this zone meets the nasal septum; laterally, it crosses the orifice of the pharyngotympanic tube; and it passes posteriorly at the union of the soft palate and the lateral wall. Numerous mucous glands surround the tubal orifices.
ADENOID OR PHARYNGEAL TONSIL
The adenoid, or pharyngeal tonsil, is a median mass of mucosa-associated lymphoid tissue (MALT) situated in the roof and posterior wall of the nasopharynx (Fig. 33.3). At its maximal size (during the early years of life) it is shaped like a truncated pyramid, often with a vertically oriented median cleft, so that its apex points towards the nasal septum and its base to the junction of the roof and posterior wall of the nasopharynx.
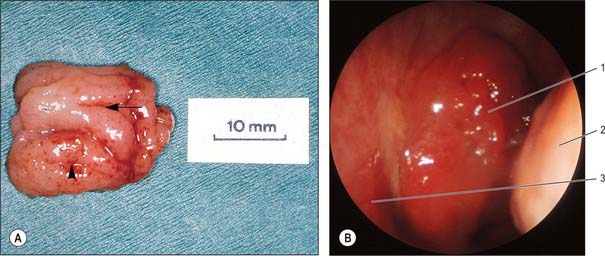
The free surface of the pharyngeal tonsil is marked by folds that radiate forwards and laterally from a median blind recess, the pharyngeal bursa (bursa of Luschka), which extends backwards and up. The recess is present in the fetus and the young, but only occasionally present in the adult, and marks the rostral end of the embryological notochord. The number and position of the folds and of the deep fissures which separate them vary. A median fold may pass forwards from the pharyngeal bursa towards the nasal septum, or instead a fissure may extend forwards from the bursa, dividing the nasopharyngeal tonsil into two distinct halves which reflect its paired developmental origins (Fig. 33.3).
The prenatal origins of the nasopharyngeal tonsil are described on page 604. After birth it initially grows rapidly, but usually undergoes a degree of involution and atrophy from the age of 8–10 years (although hypoplasia may still occur in adults up to the seventh decade). Relative to the volume of the nasopharynx, the size of the tonsil is largest at 5 years, which may account for the frequency of nasal breathing problems in preschool children, and the incidence of adenoidectomy in this age group.
Vascular supply and lymphatic drainage
The arterial supply of the pharyngeal tonsil is derived from the ascending pharyngeal and ascending palatine arteries, the tonsillar branches of the facial artery, the pharyngeal branch of the maxillary artery and the artery of the pterygoid canal (see Figs 33.5, 33.6). In addition, a nutrient or emissary vessel to the neighbouring bone, the basisphenoid artery, which is a branch of the inferior hypophysial arteries, supplies the bed of the pharyngeal tonsil and is a possible cause of persistent postadenoidectomy haemorrhage in some patients.
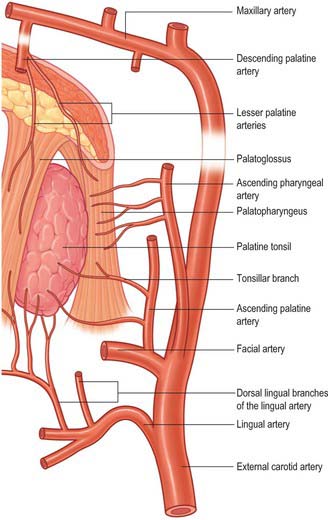
Numerous communicating veins drain the pharyngeal tonsil into the internal submucous and external pharyngeal venous plexuses. They emerge from the deep lateral surface of the tonsil and join the external palatine (paratonsillar) veins, and pierce the superior constrictor either to join the pharyngeal venous plexus, or to unite to form a single vessel that enters the facial or internal jugular vein; they may also connect with the pterygoid venous plexus.
PHARYNGOTYMPANIC TUBE
The pharyngotympanic tube (see Fig. 36.7) connects the tympanic cavity to the nasopharynx and allows the passage of air between these spaces in order to equalize the air pressure on both aspects of the tympanic membrane. It is about 36 mm long and descends anteromedially from the tympanic cavity to the nasopharynx at an angle of approximately 45° with the sagittal plane and 30° with the horizontal (these angles increase with age and elongation of the skull base). It is formed partly by cartilage and fibrous tissue and partly by bone.
The cartilaginous part, which is approximately 24 mm long, is formed by a triangular plate of cartilage, the greater part of which is in the posteromedial wall of the tube. Its apex is attached by fibrous tissue to the circumference of the jagged rim of the bony part of the tube, and its base is directly under the mucosa of the lateral nasopharyngeal wall, forming a tubal elevation (torus tubarius; Eustachian cushion) behind the pharyngeal orifice of the tube (Fig. 33.2; see Fig. 30.1). The upper part of the cartilage is bent laterally and downwards, producing a broad medial lamina and narrow lateral lamina. In transverse section it is hook-like and incomplete below and laterally, where the canal is composed of fibrous tissue. The cartilage is fixed to the cranial base in the groove between the petrous part of the temporal bone and the greater wing of the sphenoid, and ends near the root of the medial pterygoid plate. The cartilaginous and bony parts of the tube are not in the same plane, the former descending a little more steeply than the latter. The diameter of the tube is greatest at the pharyngeal orifice, least at the junction of the two parts (the isthmus), and increases again towards the tympanic cavity.
The pharyngotympanic tube is innervated by filaments from the tympanic plexus and from the pharyngeal branch of the pterygopalatine ganglion. The tympanic plexus ramifies on the promontory in the middle ear cavity and is formed by the tympanic branch of the glossopharyngeal nerve and caroticotympanic nerves of sympathetic origin (see also Ch. 36).
OROPHARYNX
BOUNDARIES
The oropharynx extends from below the soft palate to the upper border of the epiglottis (Figs 33.1, 33.2). It opens into the mouth through the oropharyngeal isthmus, demarcated by the palatoglossal arch, and faces the pharyngeal aspect of the tongue. Its lateral wall consists of the palatopharyngeal arch and palatine tonsil (see Fig. 30.3). Posteriorly, it is level with the bodies of the second, and upper part of the third, cervical vertebrae (Fig. 33.2).
SOFT PALATE
The soft palate is a mobile flap suspended from the posterior border of the hard palate, sloping down and back between the oral and nasal parts of the pharynx (Figs 33.2, 33.4). The boundary between the hard and soft palate is readily palpable and may be distinguished by a change in colour, the soft palate being a darker red with a yellowish tint. The soft palate is a thick fold of mucosa enclosing an aponeurosis, muscular tissue, vessels, nerves, lymphoid tissue and mucous glands; almost half its thickness is represented by numerous mucous glands which lie between the muscles and the oral surface of the soft palate. The latter is covered by a stratified squamous epithelium, while the nasal surface is covered with a ciliated columnar epithelium. In most individuals, two small pits, the fovea palatini, may be seen, one on each side of the midline: they represent the orifices of ducts from some of the minor mucous glands of the palate. In its usual relaxed and pendant position, the anterior (oral) surface of the soft palate is concave, and has a median raphe. The posterior aspect is convex and continuous with the nasal floor, the anterosuperior border is attached to the posterior margin of the hard palate, and the sides blend with the pharyngeal wall. The inferior border is free, and hangs between the mouth and pharynx. A median conical process, the uvula, projects downwards from its posterior border (see Fig. 30.3). Taste buds are found on the oral aspect of the soft palate.
The anterior third of the soft palate contains little muscle and consists mainly of the palatine aponeurosis. This region is less mobile and more horizontal than the rest of the soft palate and is the chief area acted upon by tensor veli palatini.
Palatoglossal and palatopharyngeal arches
The lateral wall of the oropharynx presents two prominent folds, the palatoglossal and palatopharyngeal folds (anterior and posterior pillars of the fauces respectively) (see Fig. 30.3). The palatoglossal arch, the anterior fold, runs from the soft palate to the side of the tongue and contains palatoglossus. The palatopharyngeal arch, the posterior fold, projects more medially and passes from the soft palate to merge with the lateral wall of the pharynx; it contains palatopharyngeus. A triangular tonsillar fossa (tonsillar sinus), lies on each side of the oropharynx between the diverging palatopharyngeal and palatoglossal arches, and contains the palatine tonsil.
Uvulopalatopharyngoplasty
The pharyngeal airway is kept patent in the patient who is awake by the combined dilating action of genioglossus, tensor veli palatini, geniohyoid and stylohyoid, which act to counter the negative pressure generated in the lumen of the pharynx during inspiration. The tone in the muscles is reduced during sleep, and is also affected by alcohol and other sedatives, hypothyroidism and a variety of neurological disorders. If the dilator muscle tone is insufficient, the walls of the pharynx may become apposed. Intermittent pharyngeal obstruction may cause snoring, and complete obstruction may cause apnoea, hypoxia and hypercarbia which lead to arousal and sleep disturbance.
PALATINE TONSIL
The right and left palatine tonsils form part of the circumpharyngeal lymphoid ring. Each tonsil is an ovoid mass of lymphoid tissue situated in the lateral wall of the oropharynx (Fig. 33.5; see Figs 30.3, 30.5). Size varies according to age, individuality and pathological status (tonsils may be hypertrophied and/or inflamed). It is therefore difficult to define the normal appearance of the palatine tonsil. For the first 5 or 6 years of life the tonsils increase rapidly in size. They usually reach a maximum at puberty, when they average 20–25 mm in vertical, and 10–15 mm in transverse, diameters, and they project conspicuously into the oropharynx. Tonsillar involution begins at puberty, when the reactive lymphoid tissue begins to atrophy, and by old age only a little tonsillar lymphoid tissue remains.
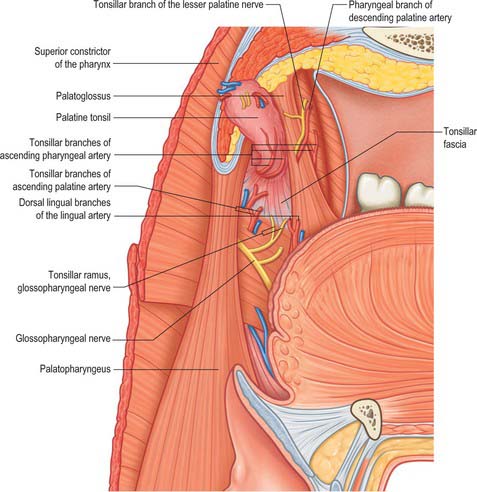
The lateral or deep surface of the tonsil spreads downwards, upwards and forwards. Inferiorly, it invades the dorsum of the tongue, superiorly, the soft palate, and, anteriorly, it may extend for some distance under the palatoglossal arch. This deep, lateral aspect is covered by a layer of fibrous tissue, the tonsillar hemicapsule. The latter is separable with ease for most of its extent from the underlying muscular wall of the pharynx, which is formed here by the superior constrictor, and sometimes by the anterior fibres of palatopharyngeus, with styloglossus on its lateral side (Fig. 33.5). Anteroinferiorly, the hemicapsule adheres to the side of the tongue and to palatoglossus and palatopharyngeus. In this region, the tonsillar artery, a branch of the facial artery, pierces the superior constrictor to enter the tonsil, accompanied by venae comitantes (Fig. 33.5). An important and sometimes large vein, the external palatine or paratonsillar vein, descends from the soft palate lateral to the tonsillar hemicapsule before piercing the pharyngeal wall. Haemorrhage from this vessel from the upper angle of the tonsillar fossa may complicate tonsillectomy. The muscular wall of the tonsillar fossa separates the tonsil from the ascending palatine artery, and, occasionally, from the tortuous facial artery itself, which may lie near the pharyngeal wall at the lower tonsillar level. The glossopharyngeal nerve lies immediately lateral to the muscular wall of the tonsillar fossa (Figs 33.4A, 33.5): it is at risk if the wall is pierced, and is commonly temporarily affected by oedema following tonsillectomy. The internal carotid artery lies approximately 25 mm behind and lateral to the tonsil. In some individuals the styloid process may be elongated and deviate towards the tonsillar bed.
Vascular supply and lymphatic drainage
The arterial blood supply to the palatine tonsil is derived from branches of the external carotid artery (Figs 33.5, 33.6). Three arteries enter the tonsil at its lower pole. The largest is the tonsillar artery, which is a branch of the facial, or sometimes the ascending palatine, artery. It ascends between medial pterygoid and styloglossus, perforates the superior constrictor at the upper border of styloglossus, and ramifies in the tonsil and posterior lingual musculature (Figs 35.5, 33.6). The other arteries found at the lower pole are the dorsal lingual branches of the lingual artery, which enter anteriorly, and a branch from the ascending palatine artery, which enters posteriorly to supply the lower part of the palatine tonsil. The upper pole of the tonsil also receives branches from the ascending pharyngeal artery, which enter the tonsil posteriorly, and from the descending palatine artery and its branches, the greater and lesser palatine arteries. All of these arteries enter the deep surface of the tonsil, branch within the connective tissue septa, narrow to become arterioles and then give off capillary loops into the follicles, interfollicular areas and the cavities within the base of the reticulated epithelium. The capillaries rejoin to form venules, many with high endothelia, and the veins return within the septal tissues to the hemicapsule as tributaries of the pharyngeal drainage. The tonsillar artery and its venae comitantes often lie within the palatoglossal fold, and may haemorrhage if this fold is damaged during surgery.
Innervation
The tonsillar region is innervated by tonsillar branches of the maxillary and glossopharyngeal nerves (Fig. 33.5). The fibres from the maxillary nerve pass through, but do not synapse in, the pterygopalatine ganglion; they are distributed through the lesser palatine nerves and form a plexus (the circulus tonsillaris) around the tonsil together with the tonsillar branches of the glossopharyngeal nerve. Nerve fibres from this plexus are also distributed to the soft palate and the region of the oropharyngeal isthmus. The tympanic branch of the glossopharyngeal nerve supplies the mucous membrane lining the tympanic cavity. Infection, malignancy and postoperative inflammation of the tonsil and tonsillar fossa may therefore be accompanied by pain referred to the ear.
LARYNGOPHARYNX
BOUNDARIES
The laryngopharynx is situated behind the entire length of the larynx (known clinically as the hypopharynx) and extends from the superior border of the epiglottis, where it is delineated from the oropharynx by the lateral glossoepiglottic folds, to the inferior border of the cricoid cartilage, where it becomes continuous with the oesophagus (Figs 33.1, 33.2). The laryngeal inlet lies in the upper part of its incomplete anterior wall, and the posterior surfaces of the arytenoid and cricoid cartilages lie below this opening.
The obliquely sloping inlet of the larynx lies in the anterior part of the laryngopharynx and is bounded above by the epiglottis, below by the arytenoid cartilages of the larynx, and laterally by the aryepiglottic folds (Fig. 33.1). Below the inlet, the anterior wall of the laryngopharynx is formed by the posterior surface of the cricoid cartilage.
PHARYNGEAL FASCIA
The two named layers of fascia in the pharynx are the pharyngobasilar and buccopharyngeal fascia. The fibrous layer that supports the pharyngeal mucosa is thickened above the superior constrictor to form the pharyngobasilar fascia (see Fig. 33.9). It is attached to the basilar part of the occipital bone and the petrous part of the temporal bone medial to the pharyngotympanic tube, and to the posterior border of the medial pterygoid plate and the pterygomandibular raphe. Inferiorly, it diminishes in thickness, but is strengthened posteriorly by a fibrous band attached to the pharyngeal tubercle of the occipital bone which descends as the median pharyngeal raphe of the constrictors. This fibrous layer is really the internal epimysial covering of the muscles and their aponeurotic attachment to the base of the skull. The thinner, external part of the epimysium is the buccopharyngeal fascia, which covers the superior constrictor and passes forwards over the pterygomandibular raphe to cover buccinator.
PHARYNGEAL TISSUE SPACES
Tissue spaces between the layers of cervical fascia are described in Chapter 28: tissue spaces around the larynx are described in Chapter 34.
MUSCLES OF THE SOFT PALATE AND PHARYNX
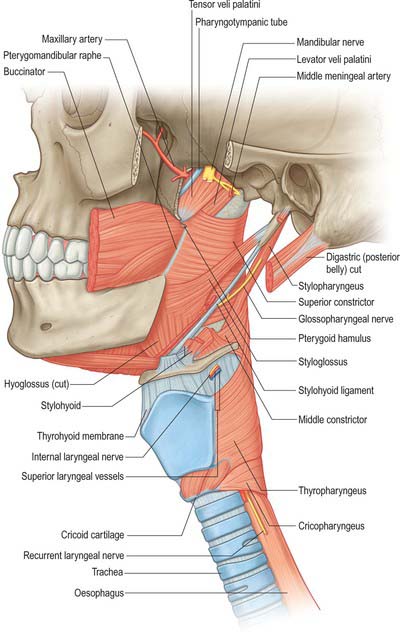
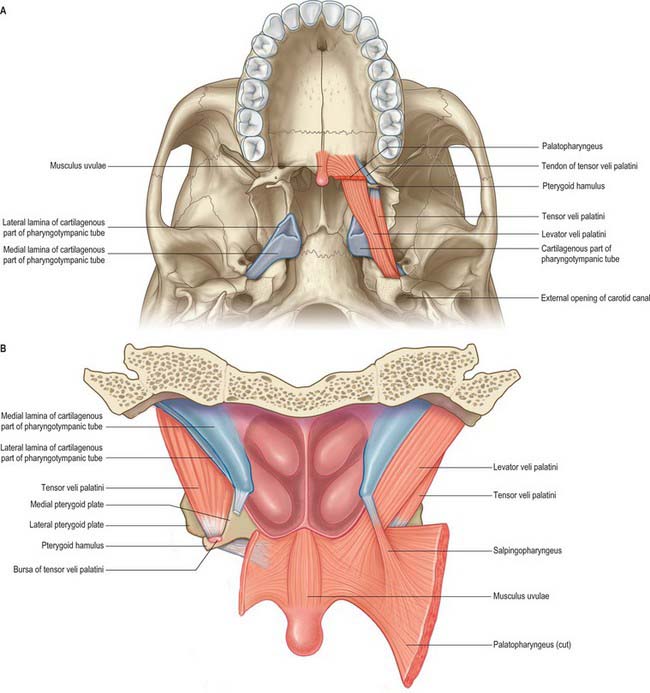
Levator veli palatini
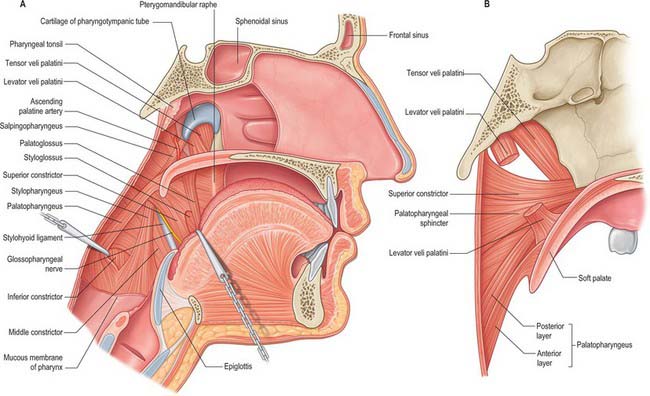
Levator veli palatini arises by a small tendon from a quadrilateral roughened area on the medial end of the inferior surface of the petrous part of the temporal bone, in front of the lower opening of the carotid canal (Figs 33.4, 33.7, 33.8, 33.9). Additional fibres arise from the inferior aspect of the cartilaginous part of the pharyngotympanic tube and from the vaginal process of the sphenoid bone. At its origin the muscle is inferior rather than medial to the pharyngotympanic tube and only crosses medial to it at the level of the medial pterygoid plate. It passes medial to the upper margin of the superior constrictor and anterior to salpingopharyngeus. Its fibres spread in the medial third of the soft palate between the two strands of palatopharyngeus to attach to the upper surface of the palatine aponeurosis as far as the midline, where they interlace with those of the contralateral muscle. Thus, the two levator muscles form a sling above and just behind the palatine aponeurosis.
Tensor veli palatini
Tensor veli palatini arises from the scaphoid fossa of the pterygoid process and posteriorly from the medial aspect of the spine of the sphenoid bone (Figs 33.4, 33.7, 33.8, 33.9; see Fig. 30.6). Between these two sites it is attached to the anterolateral membranous wall of the pharyngotympanic tube (including its narrow isthmus where the cartilaginous medial two-thirds meets the bony lateral one-third). Some fibres may be continuous with those of tensor tympani. Inferiorly, the fibres converge on a delicate tendon that turns medially around the pterygoid hamulus to pass through the attachment of buccinator to the palatine aponeurosis and the osseous surface behind the palatine crest on the horizontal plate of the palatine bone. There is a small bursa between the tendon and the pterygoid hamulus.
The muscle is thin and triangular and lieslateral to the medial pterygoid plate, pharyngotympanic tube and levator veli palatini. Its lateral surface contacts the upper and anterior part of medial pterygoid, the mandibular, auriculotemporal and chorda tympani nerves, the otic ganglion and the middle meningeal artery.
Palatoglossus
Palatoglossus is narrower at its middle than at its ends (Fig. 33.4). Together with its overlying mucosa it forms the palatoglossal arch or fold (see Fig. 30.3). It arises from the oral surface of the palatine aponeurosis where it is continuous with its contralateral fellow. It extends forwards, downwards and laterally in front of the palatine tonsil to the side of the tongue. Some of its fibres spread over the dorsum of the tongue, others pass deeply into its substance to intermingle with fibres of the intrinsic transverse muscle.
Palatopharyngeus
Palatopharyngeus and its overlying mucosa form the palatopharyngeal arch (see Fig. 30.3). Within the soft palate, palatopharyngeus is composed of two fasciculi that are attached to the upper surface of the palatine aponeurosis; they lie in the same plane but are separated from each other by levator veli palatini (Figs 33.4, 33.8). The thicker, anterior fasciculus arises from the posterior border of the hard palate as well as the palatine aponeurosis, where some fibres interdigitate across the midline. The posterior fasciculus is in contact with the mucosa of the pharyngeal aspect of the palate, and joins the posterior band of the contralateral muscle in the midline. The two layers unite at the posterolateral border of the soft palate, and are joined by fibres of salpingopharyngeus. Passing laterally and downwards behind the tonsil, palatopharyngeus descends posteromedial to and in close contact with stylopharyngeus, to be attached with it to the posterior border of the thyroid cartilage. Some fibres end on the side of the pharynx, attached to pharyngeal fibrous tissue, and others cross the midline posteriorly, decussating with those of the contralateral muscle. Palatopharyngeus thus forms an incomplete internal longitudinal muscular layer in the wall of the pharynx.
Passavant’s muscle (palatopharyngeal sphincter)
The existence of Passavant’s muscle remains controversial. It has been described as a part of the superior constrictor and palatopharyngeus muscles. An alternative view holds that it is a distinct palatine muscle that arises from the anterior and lateral parts of the upper surface of the palatine aponeurosis, lies lateral to levator veli palatini, blends internally with the upper border of the superior constrictor, and encircles the pharynx as a sphincter-like muscle (Figs 33.4, 33.8). Whatever its origin, when it contracts, it forms a ridge (Passavant’s ridge) when the soft palate is elevated. The change from columnar, ciliated, ‘respiratory’ epithelium to stratified, squamous epithelium that takes place on the superior aspect of the soft palate occurs along the line of attachment of the palatopharyngeal sphincter to the palate. The muscle is hypertrophied in cases of complete cleft palate.
Musculus uvulae
Musculus uvulae arises from the posterior nasal spine of the palatine bone and the superior surface of the palatine aponeurosis, and lies between the two laminae of the aponeurosis (Figs 33.1, 33.8). It runs posteriorly above the sling formed by levator veli palatini and inserts beneath the mucosa of the uvula. The two sides of the muscle are united along most of its length.
Salpingopharyngeus
Salpingopharyngeus arises from the inferior part of the cartilage of the pharyngotympanic tube near its pharyngeal opening and passes downwards within the salpingopharyngeal fold to blend with palatopharyngeus (Figs 33.1, 33.4A, 33.8).
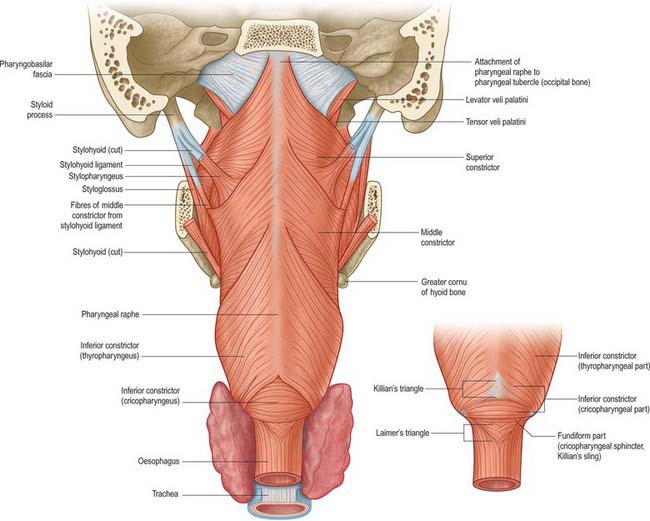
Stylopharyngeus
Stylopharyngeus is a long slender muscle, cylindrical above and flat below. It arises from the medial side of the base of the styloid process, descends along the side of the pharynx, and passes between the superior and middle constrictors to spread out beneath the mucous membrane (Figs 33.4, 33.9; see Fig. 30.6). Some fibres merge into the constrictors and the lateral glossoepiglottic fold, while others join fibres of palatopharyngeus and are attached to the posterior border of the thyroid cartilage. The glossopharyngeal nerve curves round the posterior border and the lateral side of stylopharyngeus, and passes between the superior and middle constrictors to reach the tongue.
Superior constrictor
The superior constrictor is a quadrilateral sheet of muscle and is thinner than the other two constrictors. It is attached anteriorly to the pterygoid hamulus (and sometimes to the adjoining posterior margin of the medial pterygoid plate), the posterior border of the pterygomandibular raphe, the posterior end of the mylohyoid line of the mandible, and, by a few fibres, to the side of the tongue (Figs 33.1, 33.4, 33.5, 33.7, 33.9; see Fig. 30.6). The fibres curve back into a median pharyngeal raphe which is attached superiorly to the pharyngeal tubercle on the basilar part of the occipital bone.
The upper border of the superior constrictor is separated from the cranial base by a crescentic interval which contains levator veli palatini, the pharyngotympanic tube and an upward projection of pharyngobasilar fascia. The lower border is separated from the middle constrictor by stylopharyngeus and the glossopharyngeal nerve (Fig. 33.8). Anteriorly, the pterygomandibular raphe separates the superior constrictor from buccinator, and, posteriorly, the superior constrictor lies on the prevertebral muscles and fascia, from which it is separated by the retropharyngeal space. The ascending pharyngeal artery, pharyngeal venous plexus, glossopharyngeal and lingual nerves, styloglossus, middle constrictor, medial pterygoid, stylopharyngeus and the stylohyoid ligament all lie laterally, and palatopharyngeus, the tonsillar capsule and the pharyngobasilar fascia lie internally.
Middle constrictor
The middle constrictor is a fan-shaped sheet attached anteriorly to the lesser cornu of the hyoid and the lower part of the stylohyoid ligament (the chondropharyngeal part of the muscle), and to the whole of the upper border of the greater cornu of the hyoid (the ceratopharyngeal part) (Figs 33.4A, 33.7, 33.9; see Fig. 30.6). The lower fibres descend deep to the inferior constrictor to reach the lower end of the pharynx; the middle fibres pass transversely and the superior fibres ascend and overlap the superior constrictor. All fibres insert posteriorly into the median pharyngeal raphe.
Inferior constrictor
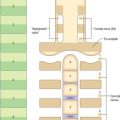
The inferior constrictor is the thickest of the three constrictor muscles, and is usually described in two parts, thyropharyngeus and cricopharyngeus (Figs 33.4A, 33.7, 33.9; see Fig. 30.6). Thyropharyngeus arises from the oblique line of the thyroid lamina, a strip of the lamina behind this, and by a small slip from the inferior cornu. Some additional fibres arise from a tendinous cord that loops over cricothyroid. Cricopharyngeus arises from the side of the cricoid cartilage between the attachment of cricothyroid and the articular facet for the inferior thyroid cornu. Some authors have described cricopharyngeus as consisting of a superficial upper oblique portion, the pars oblique, and a lower, deeper, transverse portion, the pars fundiformis. The upper part attaches to the median raphe while the lower part forms a circular band that lacks a median raphe. The area demarcated by the pars oblique and pars fundiformis of cricopharyngeus is termed Killian’s dehiscence (or Killian’s triangle). A second triangular area, Laimer’s triangle, can be identified beneath cricopharyngeus between the longitudinal fibres of the oesophagus as they pass laterally on either side to attach to the cricoid cartilage: only the circular muscle of the oesophagus forms the wall here. Both triangles are postulated to be sites of weakness in the wall of the pharynx and oesophagus, and are therefore areas where diverticula could potentially form. Both cricopharyngeus and thyropharyngeus spread posteromedially to join the contralateral muscle. Thyropharyngeus is inserted into the median pharyngeal raphe, and its upper fibres ascend obliquely to overlap the middle constrictor; however, cricopharyngeus blends with the circular oesophageal fibres around the narrowest part of the pharynx.
Cricopharyngeus contains about 40% of endomysial connective tissue, much of which is elastic, but it lacks muscle spindles. It contains both slow-twitch type I and fast-twitch type II fibres, a structural arrangement that underpins the various functions of the upper oesophageal sphincter, i.e. maintaining constant basal tone, yet being able to relax and contract rapidly during swallowing, belching and vomiting. The tonic activity of cricopharyngeus between swallows prevents influx of air during inspiration and tracheobronchial aspiration and pharyngeal reflux of oesophageal contents during oesophageal peristalsis. For further reading, see Lang and Shaker (2000).
ANATOMY OF SWALLOWING (DEGLUTITION)
ORAL PREPARATORY PHASE
In the oral preparatory phase, the essential action is chewing: the mandible is moved by the action of the jaw elevators and depressors (see Ch. 31), and the food is reduced by the grinding action of the teeth and simultaneously mixed with saliva. The lips are maintained as a tight labial seal by the contraction of orbicularis oris: buccinator performs a similar function for the cheeks. In this way, the sulci are closed, the vestibule normally remains empty, and any food that enters the vestibule is returned to the oral cavity proper. Buccinator also keeps the cheeks taut, ensuring that they are kept clear of the occlusal surfaces of the teeth during chewing: loss of the nerve supply to buccinator as a result of damage to the facial nerve results in painful and repeated lacerations of the cheeks.
The soft palate is depressed during this phase and premature spillage of food is common. Spillage occurs because the soft palate is not in continuous contact with the posterior part of the tongue, as was once thought (Hiiemae & Palmer 1999). Bolus formation appears to involve several cycles of food being transported from the anterior to the posterior part of the tongue through the palatoglossal and palatopharyngeal arches until a bolus accumulates on the oropharyngeal surface of the tongue (retrolingual loading). Throughout this phase, the lateral and rotatory tongue movements that deliver the food to the teeth for grinding and reduction are crucial for normal bolus formation. If effective tongue movements do not occur, chewing will be compromised. The end of this phase of swallowing is marked by the tongue holding the bolus of food that has been formed against the hard palate in readiness for transport to the posterior part of the oral cavity.
PHARYNGEAL PHASE
The airway is sealed at the laryngeal inlet by closure of the glottis (see Ch. 34), and the epiglottis is retroflexed over the laryngeal aditus as a result of passive pressure from the base of the tongue and active contraction of the aryepiglottic muscles. The conventional view that laryngeal closure during swallowing occurs from inferior to superior, i.e the vocal folds adduct first and the epiglottis covers the arytenoids and glottis last, has been challenged by studies using simultaneous electromyography and fibreoptic endoscopic evaluation of swallowing, FEES, which have reported that the aryepiglottic folds close before vocal fold adduction during a swallow. It is probably reasonable to assume that the sequence of events that close the glottis may alter according to the type of swallow and consistency of the bolus. What is not in dispute is that the hyoid bone and larynx are raised and pulled anteriorly by the suprahyoid muscles and the longitudinal muscles of the pharynx, so that the laryngeal inlet is brought forward under the bulge of the posterior tongue, i.e. out of the path of the bolus. This action helps expand the hypopharyngeal space and relax the upper oesophageal sphincter, which is also raised by several centimetres. The bolus passes over the reflected anterior surface of the epiglottis and is swept through the laryngopharynx to the upper oesophageal sphincter.
Gag reflex
Traditionally, the stimulus for triggering a swallow has been regarded as contact with the posterior wall of the pharynx, since this is usually where the gag reflex is triggered (see p. 283 and Fig. 19.12). However, it has become clear that many regions of the oropharynx, when appropriately stimulated by the presence of food or liquid, are capable of triggering a swallow, although some regions are more sensitive than others, e.g. the area over the palatoglossal arches. Moreover, there appears to be little relationship between a functioning gag reflex and the ability to swallow normally. Individuals with a reduced or absent gag reflex can swallow safely; conversely, the presence of a brisk and clear gag reflex is not always associated with the ability to swallow normally.
OESOPHAGEAL PHASE
The third, or oesophageal, stage begins after the relaxation of the upper oesophageal sphincter has allowed the bolus to enter the oesophagus. Sequential waves of contractions of the oesophageal musculature subsequently propel the bolus down to the lower oesophageal sphincter, which opens momentarily to admit the bolus to the stomach. The oesophageal phase of swallowing is much more variable than the other phases, and lasts between 8 and 20 seconds.
SWALLOWING PATTERN GENERATOR
The patterning and timing of striated muscle contraction during swallowing are generated at a brainstem level in a network of neural circuits which collectively form a central swallowing pattern generator (SPG). Experimental neurophysiological and tracer studies in animals have shown that the SPG includes several brainstem motor nuclei and two groups of interneurones in the dorsal and ventral medulla, a dorsal swallowing group (DSG) and a ventral swallowing group (VSG). The DSG is located in the nucleus of the solitary tract and possibly in the adjacent reticular formation, and contains neurones that generate the swallowing pattern, probably triggered by convergent information from both cortical and peripheral inputs. The afferent feedback from the branches of the superior laryngeal nerve which innervate the valleculae, epiglottis and supraglottic part of the larynx, and which relay through the nucleus of the solitary tract, facilitates laryngeal closure during swallowing (Jafari et al 2003). The VSG is in the ventrolateral medulla above the nucleus ambiguus and contains neurones that act as ‘switches’, distributing the swallowing drive to motoneurone pools in the trigeminal, facial and hypoglossal nuclei and in the nucleus ambiguus.
Supramedullary influence on swallowing
Techniques including transcranial magnetic stimulation, cortical evoked potentials, positron emission tomography and functional magnetic resonance imaging (fMRI) have all been exploited in the study of volitional swallowing in humans. Thus far, these studies have shown that numerous cortical and subcortical regions are recruited in swallowing, although, as yet, the functional contributions made by each region have not been established, and it is not known how they influence the SPG. The most consistent cortical activation occurs in the primary motor and somatosensory cortices. Perhaps not surprisingly, the greatest area of fMRI activation in the primary motor cortex occurs over the portion of the precentral gyrus where the face, tongue and pharynx are represented: there is evidence for functional asymmetry between the right and left oral sensorimotor cortices (Martin et al 2004). The activity of the SPG is coordinated with other medullary reflexes, e.g. it is difficult to elicit a swallow when the cortical masticatory centres are stimulated.
SWALLOWING IN THE NEONATE
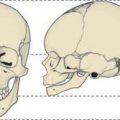
In the adult, the tip of the epiglottis is significantly lower than the inferior edge of the soft palate. In the neonate, the larynx is high in the neck and the epiglottis may extend above the soft palate so that the laryngeal airway is in direct continuity with the posterior nares (Fig. 33.10): a potential space is therefore formed between the soft palate above, the epiglottis behind and the tongue anteroinferiorly. In other mammals with an oropharyngeal anatomy similar to that of the human infant, up to 14 cycles of tongue movement or oral phases cause the accumulation of food in this space. Subsequent emptying of the space is a single event followed by movement of the bolus down the oesophagus. The ratio of accumulation cycles to swallow events in the human neonate is approximately 1.5 : 1, which is lower than in other mammals, but still implies some temporary accumulation. In the case of a liquid bolus, accumulated material may be passed laterally to the epiglottis through the piriform fossae rather than over the flexed epiglottis, although it is not known whether this happens in the human infant.
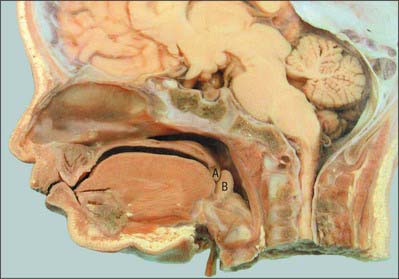
Swallow safety is critical at all ages. In the neonate, where coordination of suckling, swallowing and breathing is not fully developed, the potential risk of airway obstruction and/or aspiration of ingested milk or other material during swallowing is reduced because the intranarial larynx prevents the bolus from entering the larynx before and after a swallow (see Ch. 34). The change towards adult anatomy and coordination of the phases of swallowing starts a few months after birth. Differential growth in length of the human pharynx causes the larynx to take up its low adult position and the epiglottis to lose contact with the soft palate: the larynx reaches its final position around the time of puberty. The adult anatomy does not allow any significant accumulation of food to occur immediately anterior to the epiglottis, which means that the transport of food through the fauces has to bear a 1 : 1 relationship to pharyngeal and oesophageal transit. Moreover, the lowered larynx compromises the previously protected airway: the hyoid and larynx are therefore raised and pulled forwards during a swallow in order to minimize the risk of deglutitive aspiration.
Bluestone CD. Anatomy and physiology of the Eustachian tube. Cummings CW, et al, editors. Otolaryngology: Head and Neck Surgery, 3rd edn., vol 4. St Louis: Mosby, 1998;3003-3025.
Bonnington A, Mahon M, Whitmore I. A histological study of the cricopharyngeus muscle in man. J Anat. 1988;156:27-37.
Brownlow H, Whitmore I, Willan PLT. A quantitative study of the histochemical and morphometric characteristics of the human cricopharyngeus muscle. J Anat. 1989;166:67-75.
Freelander E. Blood supply of the human levator and tensor veli palatini muscles. Clin Anat. 1992;5:34-44.
Graney DO, Retruzzelli GJ, Myers EW. Anatomy. Cummings CW, et al, editors. Otolaryngology: Head and Neck Surgery, 3rd edn., vol 2. St Louis: Mosby, 1998;1327-1348.
Gray SD, Pinborough-Zimmerman J. Velopharyngeal incompetence. In: Cummings CW, et al. Otolaryngology: Head and Neck Surgery, vol 5: Pediatric Otolaryngology. 3rd edn. St Louis: Mosby; 1998:174-187.
Harris ML, Julyan P, Kulkarni B, et al. Mapping metabolic brain activation during human volitional swallowing: a positron emission tomography study using [18F]fluorodeoxyglucose. J Cereb Blood Flow Metab. 2005;25:520-526.
Hiiemae KM, Palmer JB. Food transport and bolus formation during complete feeding sequences on foods of different initial consistency. Dysphagia. 1999;14:31-42.
Jafari S, Prince RA, Kim DY, Paydarfar D. Sensory regulation of swallowing and airway protection: a role for the internal superior laryngeal nerve in humans. J Physiol. 2003;550:287-304.
Lang IM, Shaker R. An overview of the upper esophageal sphincter. Curr Gastroenterol Rep. 2000;2:185-190.
Martin RE, MacIntosh BJ, Smith RC, et al. Cerebral areas processing swallowing and tongue movement are overlapping but distinct: a functional magnetic resonance imaging study. J Neurophysiol. 2004;92:2428-2443.
Sade J, editor. Basic Aspects of the Eustachian Tube and Middle Ear Disease. Geneva: Kugler & Ghedini, 1989.
Thexton A. Some aspects of swallowing. In: Harris M, Edgar M, Meghji S. Clinical Oral Science. Oxford: Wright; 1998:150-166.
Thexton AJ, Crompton AW. The control of swallowing. Linden RWA, editor. The Scientific Basis of Eating. Taste and Smell, Salivation, Mastication and Swallowing and their Dysfunctions. Frontiers of Oral Biology Series vol 9. Basel: Karger; 1998:168-222.
Wood-Jones I. The nature of the soft palate. J Anat. 1940;77:147-170.
Describes the structure of the soft palate and its movements during swallowing..