[level-membership-for-basic-science-category]
18 Overview of Motor Systems
The firing rates of our motor neurons, and therefore the states of contraction of our muscles, are determined by multiple influences. Simple reflex arcs like the stretch reflex and more complex motor programs like the basic pattern generator for walking are built into the spinal cord and brainstem. These reflex arcs and motor programs, as well as the motor neurons themselves, are in turn influenced by various descending pathways. Finally, activity in the descending pathways is modulated by other cortical areas, the basal ganglia (see Chapter 19), and the cerebellum (see Chapter 20).
Each Lower Motor Neuron Innervates a Group of Muscle Fibers, Forming a Motor Unit
Each lower motor neuron innervates a fraction of the muscle fibers in one muscle. The combination of a lower motor neuron and all the muscle fibers it innervates is a motor unit (Fig. 18-1). Although there is a range of motor unit sizes in every muscle, their average size varies in a predictable way—those involving finely controlled muscles (e.g., extraocular muscles) contain very few muscle fibers, and those involving less finely controlled muscles may have hundreds of muscle fibers.
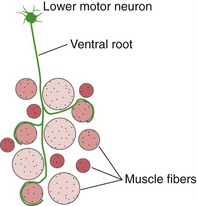
Lower Motor Neurons Are Arranged Systematically
The cell bodies of lower motor neurons are arranged systematically in the anterior horn (just as things like body parts and retinal areas are represented systematically in pathways and cortical areas). At any given spinal level, motor neurons for more proximal muscles are located medial to those for more distal muscles, and motor neurons for flexors are located dorsal to those for extensors (Fig. 18-2).
There Are Three Kinds of Muscle Fibers and Three Kinds of Motor Units
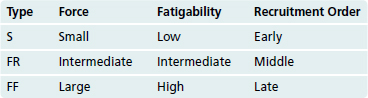
We use most muscles for multiple purposes that require different forces of contraction, from relatively weak contractions used in holding positions for long periods of time (e.g., standing) to powerful contractions that cannot be sustained for very long (e.g., sprinting, jumping). Corresponding to this, there are three different kinds of muscle fibers: slow fibers (S) that produce little force but do not fatigue much, fast fibers (FF) that produce a lot of force but fatigue quickly, and intermediate fibers (FR) with intermediate properties. All the muscle fibers innervated by a given motor neuron are of the same type, so there are also three kinds of motor units (Table 18-1).
Motor Control Systems Involve Both Hierarchical and Parallel Connections
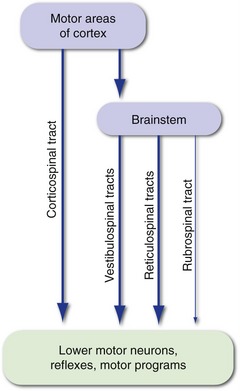
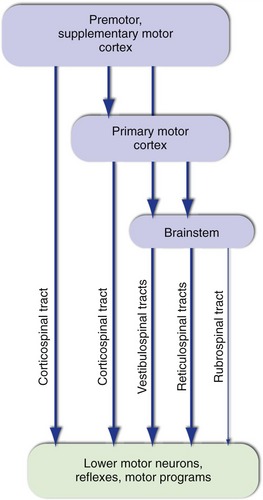
The firing rates of lower motor neurons are influenced most directly by local connections and by inputs descending from more rostral levels of the CNS (i.e., upper motor neurons). Local connections include those that mediate simple reflexes, as well as those of central pattern generators or motor programs, connections that provide the basic timing signals for rhythmic activities like walking and breathing. Parallel descending inputs from several sources influence movement both directly, by synapsing on lower motor neurons, and indirectly, by synapsing on the interneurons of reflex circuits and motor programs (Fig. 18-3). The principal sources of descending inputs are the cerebral cortex (corticospinal tract), vestibular nuclei (vestibulospinal tracts), and reticular formation (reticulospinal tracts); there is also a descending projection from the red nucleus (rubrospinal tract), but this is small and relatively unimportant in humans.
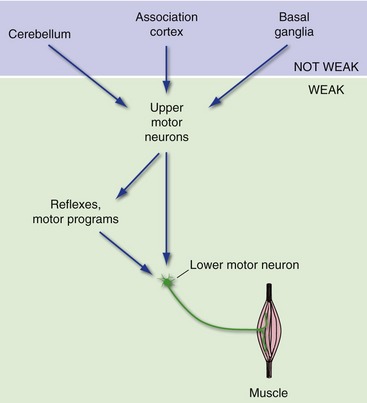
The basal ganglia, cerebellum, and certain areas of association cortex are also important in the production of movement but have little or no direct access to lower motor neurons (Fig.18-4). Rather, they affect movement by modulating the activity of upper motor neurons, particularly those in the cerebral cortex. This has important clinical consequences. Damage to motor cortex, lower motor neurons, or muscle results in weakness. In contrast, damage to the basal ganglia, cerebellum, or movement-related association cortex typically causes a variety of movement abnormalities (e.g., incoordination, involuntary movements), but not prominent weakness, because both upper and lower motor neurons are still intact.
The Corticospinal Tract Has Multiple Origins and Terminations
Corticospinal Axons Arise in Multiple Cortical Areas
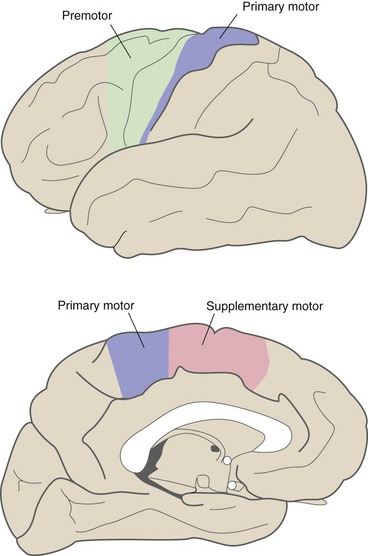
Corticospinal fibers originate from cortical areas near the central sulcus (Fig. 18-5). Many arise in primary motor cortex in the precentral gyrus and in premotor cortex, just anterior to primary motor cortex. Both areas are arranged somatotopically: Neurons projecting to cranial nerve motor neurons are most ventral, those projecting to leg motor neurons are near the top of the central sulcus, and those projecting to arm and hand motor neurons are in between. Particularly large areas are devoted to the hand and mouth. In addition, some corticospinal fibers arise in the supplementary motor area on the medial surface of the hemisphere and others arise in the postcentral gyrus (i.e., somatosensory cortex).
Corticospinal Input Is Essential for Only Some Movements
The outputs of these cortical areas provide another example of the serial-parallel connections characteristic of the motor system (Fig. 18-6). The premotor and supplementary motor areas project to primary motor cortex, as well as projecting to the spinal cord in parallel with corticospinal fibers from primary motor cortex. In addition, all three cortical areas project not only to the spinal cord but also to brainstem sites such as the reticular formation. One consequence is that damage to motor areas of the cortex has effects (described shortly) significantly different from those following damage confined specifically to corticospinal axons, for example in the pyramid. Damage restricted to the corticospinal tract spares movements mediated by things like the reticulospinal tracts and motor programs; most severely affected are skilled, dexterous movements, notably in primates the ability to use fingers individually.
Upper Motor Neuron Damage Causes a Distinctive Syndrome
Upper motor neurons are most commonly damaged in strokes and other cortical lesions that affect corticospinal (and corticobulbar) neurons. The result is spastic hemiparesis, in which the contralateral side of the body is weak, stretch reflexes are increased, and muscle tone is increased. The flexors of the upper extremity and extensors of the lower extremity are particularly affected. Babinski’s sign (dorsiflexion of the big toe and fanning of the others in response to firmly stroking the sole of the foot) is also present on the side contralateral to the lesion, and clonus (rhythmic contractions in response to maintained muscle stretch) may be seen. The increased tone collapses abruptly in response to strong efforts to overcome it (clasp-knife response).
There Are Upper Motor Neurons for Cranial Nerve Motor Nuclei
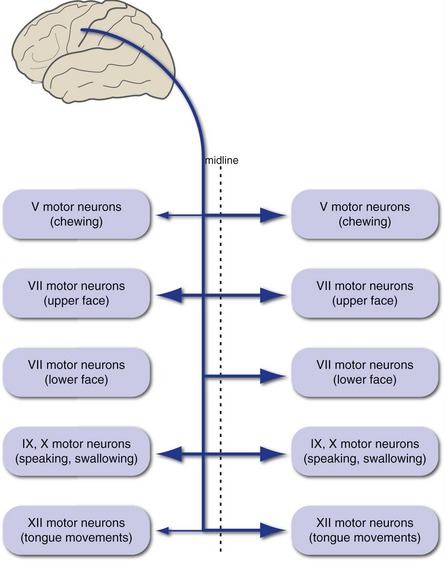
Axons of the upper motor neurons for cranial nerve motor nuclei form the corticobulbar tract (Fig. 18-7). These fibers mostly accompany the corticospinal tract until they reach the level of the nuclei in which they terminate. In contrast to corticospinal fibers, however, they are distributed bilaterally to a great extent. The result is that, with one major exception, corticobulbar damage on one side does not cause lasting contralateral weakness of muscles innervated by cranial nerves. The major exception is the muscles of the lower face, whose motor neurons are innervated predominantly by contralateral cerebral cortex, with the result that corticobulbar damage on one side causes contralateral weakness of only the lower face (see Fig. 12-9). In addition, the trigeminal and hypoglossal nuclei receive more crossed than uncrossed fibers in most individuals, and corticobulbar damage can result in slight and usually transient weakness of the contralateral jaw or tongue.
[/level-membership-for-basic-science-category][not-level-membership-for-basic-science-category]
18 Overview of Motor Systems
The firing rates of our motor neurons, and therefore the states of contraction of our muscles, are determined by multiple influences. Simple reflex arcs like the stretch reflex and more complex motor programs like the basic pattern generator for walking are built into the spinal cord and brainstem. These reflex arcs and motor programs, as well as the motor neurons themselves, are in turn influenced by various descending pathways. Finally, activity in the descending pathways is modulated by other cortical areas, the basal ganglia (see Chapter 19), and the cerebellum (see Chapter 20).
Each Lower Motor Neuron Innervates a Group of Muscle Fibers, Forming a Motor Unit
Each lower motor neuron innervates a fraction of the muscle fibers in one muscle. The combination of a lower motor neuron and all the muscle fibers it innervates is a motor unit (Fig. 18-1). Although there is a range of motor unit sizes in every muscle, their average size varies in a predictable way—those involving finely controlled muscles (e.g., extraocular muscles) contain very few muscle fibers, and those involving less finely controlled muscles may have hundreds of muscle fibers.
Lower Motor Neurons Are Arranged Systematically
The cell bodies of lower motor neurons are arranged systematically in the anterior horn (just as things like body parts and retinal areas are represented systematically in pathways and cortical areas). At any given spinal level, motor neurons for more proximal muscles are located medial to those for more distal muscles, and motor neurons for flexors are located dorsal to those for extensors (Fig. 18-2).
There Are Three Kinds of Muscle Fibers and Three Kinds of Motor Units
We use most muscles for multiple purposes that require different forces of contraction, from relatively weak contractions used in holding positions for long periods of time (e.g., standing) to powerful contractions that cannot be sustained for very long (e.g., sprinting, jumping). Corresponding to this, there are three different kinds of muscle fibers: slow fibers (S) that produce little force but do not fatigue much, fast fibers (FF) that produce a lot of force but fatigue quickly, and intermediate fibers (FR) with intermediate properties. All the muscle fibers innervated by a given motor neuron are of the same type, so there are also three kinds of motor units (Table 18-1).
