Chapter 23
Olfaction and Taste
The olfactory and taste systems sample the rich chemical environment that surrounds us. Information provided by these systems is intimately associated with the enjoyment of foods and beverages. When we refer to the taste of food, what we mean is a complex sensory experience correctly called flavor. Flavor perception results from a combination of the olfactory, taste, and somatosensory cues present in foods and beverages. Olfaction is the sensation of odors that results from the detection of odorous substances aerosolized in the environment. In contrast, taste (gustation) is the sensation evoked by stimulation of taste receptors located in the oropharyngeal cavity. The somatosensory system contributes to the experience of flavor by detecting irritating components in smells like ammonia or the “hot” in spicy food like peppers. In general, this is made possible by the activation of somatosensory endings by strong “aversive” chemical substances. Somatosensory cues include thermal, tactile, and the common chemical sense, and this information is relayed to the brain by branches of the trigeminal nerve that innervate oral and nasal mucosa.
OVERVIEW
For many mammals, smell is the principal means by which information about the environment is received. Macrosmatic animals have a well-developed sense of smell on which they rely to recognize food, to detect predators and prey, and to locate potential mates. In animals such as humans that are less dependent on smell (microsmatic animals), the olfactory system is less well developed. However, humans are still able to distinguish thousands of odors, many at extremely low concentrations. Through connections with cortical and limbic structures, the olfactory system plays a role in the pleasures associated with eating and with the many scents that make up our world.
In contrast to olfaction, the taste system exhibits a limited range of sensations. Traditionally, taste sensations have been divided into sweet, salty, sour, and bitter. In addition to these four basic tastes, a taste sensation termed umami, best exemplified by the taste of monosodium glutamate, may be important for identification of amino acids. Furthermore, recent evidence suggests that taste mechanisms for fats may also exist. Combinations of these different taste qualities account for much of our taste experience. Taste input, which originates from receptors in the oropharyngeal cavity, is important to determine the acceptance or rejection of foods. This information is relayed by neural pathways that underlie various ingestive and digestive functions.
Disorders of olfaction or taste may adversely affect the individual’s quality of life. The intimate association between the chemical senses and ingestion means that chemosensory disorders impair the patient’s ability to enjoy eating. In addition, these disorders can render the patient unable to detect hazards such as gas leaks or spoiled foods.
OLFACTORY RECEPTORS
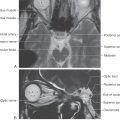
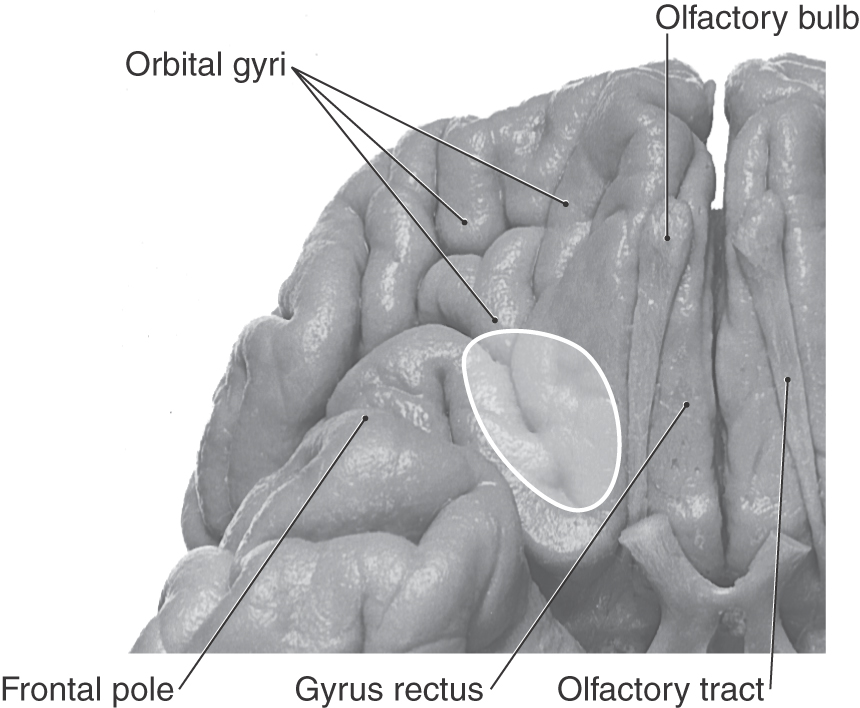
The olfactory bulb lies on the cribriform plate of the ethmoid bone. In this location it is inferior to medial aspects of the frontal lobe (Fig. 23-1), at the rostral end of the olfactory sulcus (see Fig. 23-7), and in the rostral portions of the anterior cranial fossa. Olfactory structures are especially vulnerable to facial trauma, particularly that involving the nasal bones, frontal bone, or concha of the nose.
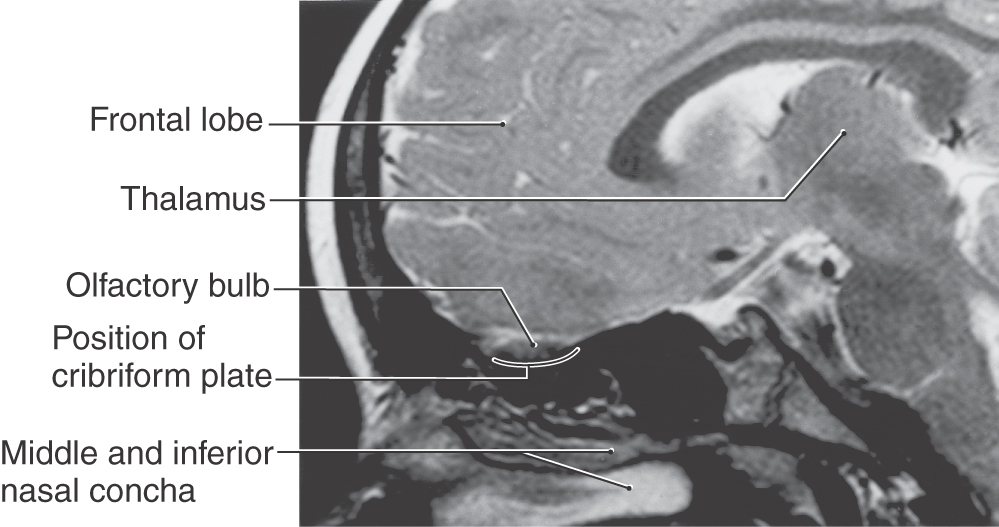
The receptors responsible for transduction of odor molecules are found in the olfactory mucosa. This portion of nasal mucosa is 1 to 2 cm2 in size and is located in the roof of the nasal cavity on the inferior surface of the cribriform plate and along the nasal septum and medial wall of the superior turbinate (Fig. 23-2). The olfactory mucosa is composed of a superficial acellular layer of mucus that covers the olfactory epithelium and underlying lamina propria. The olfactory epithelium is differentiated from the adjacent pinkish respiratory epithelium by its faint yellowish color and greater thickness. In humans the transition between olfactory and respiratory epithelia is gradual.
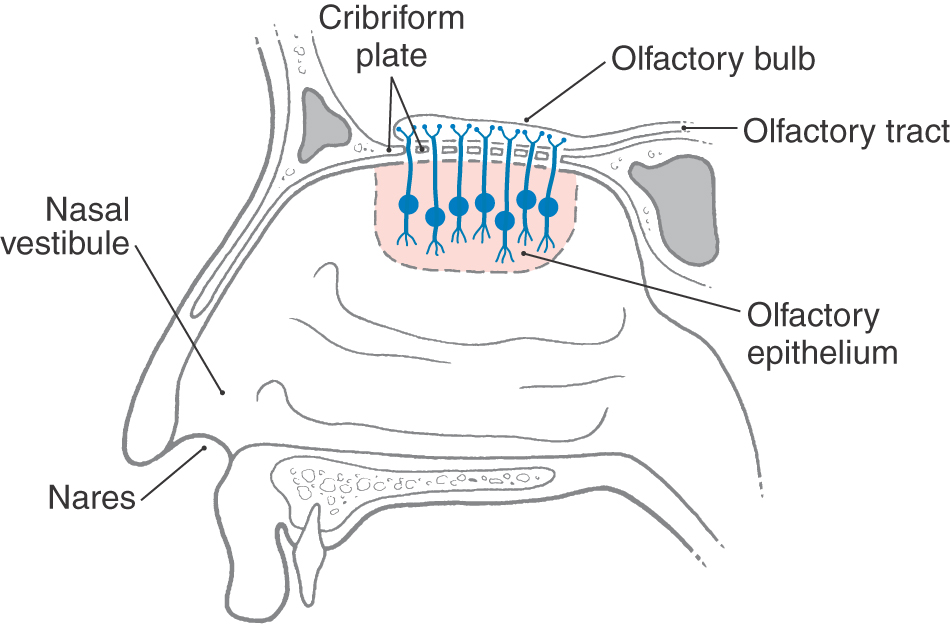
The olfactory epithelium is pseudostratified and contains three main cell types: olfactory receptor neurons, supporting cells (sustentacular cells), and basal cells (Fig. 23-3A, B). The small (5 µm) somata of bipolar olfactory receptor neurons are found in the basal two thirds of the epithelium. Each has a single thin apical dendrite and a basally located unmyelinated axon. The apical dendrite extends to the surface of the epithelium, where it terminates in a knob-like olfactory vesicle from which 10 to 30 nonmotile cilia arise and protrude into the overlying mucus layer (Fig. 23-3C, D). These olfactory cilia contain receptors for odorant molecules.
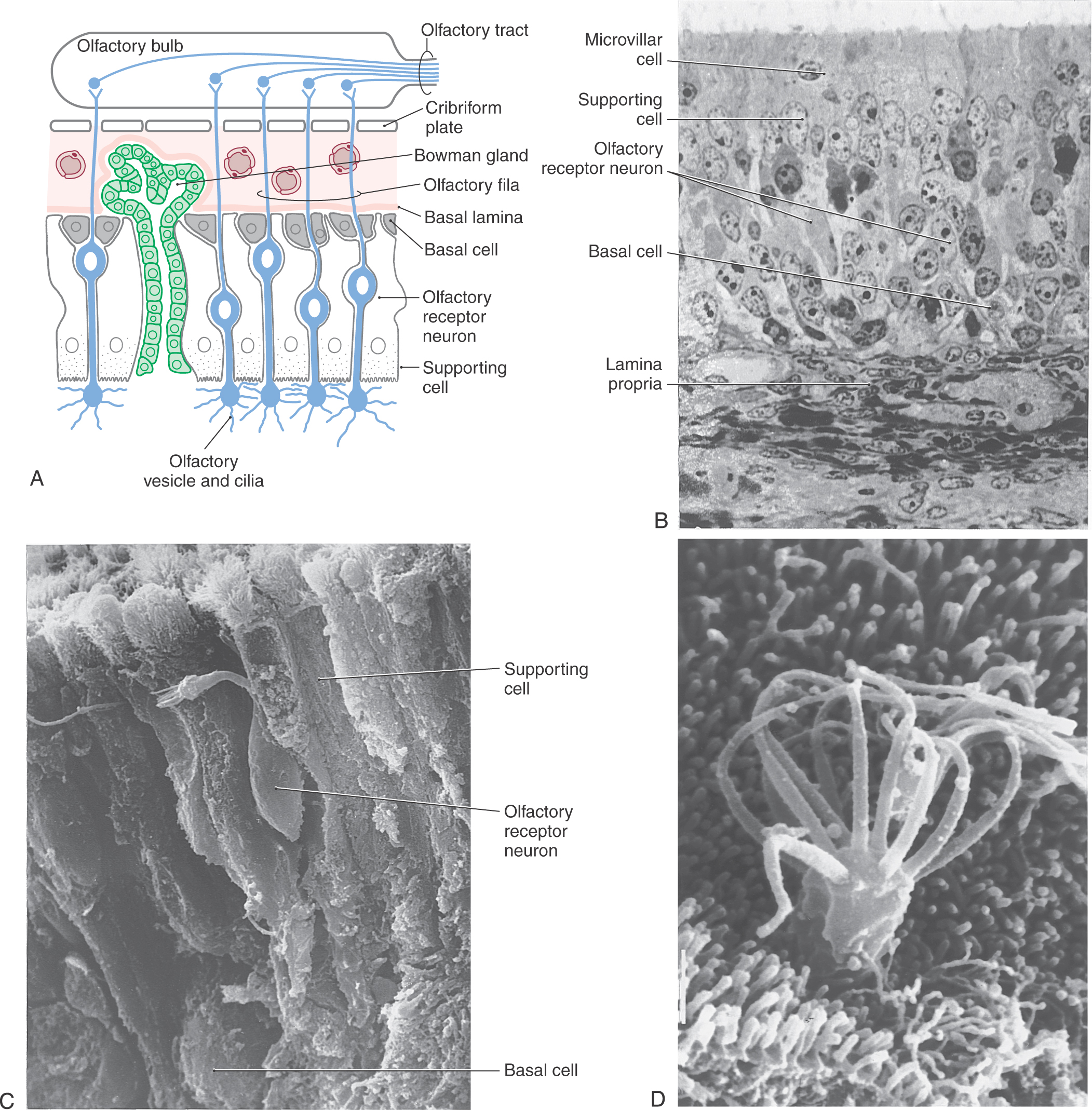
The unmyelinated axon of an olfactory receptor neuron is about 0.2 µm in diameter, making it one of the smallest in the nervous system. These axons pass through the lamina propria and group together into bundles called olfactory fila, which collectively make up the olfactory nerve (cranial nerve I) (Fig. 23-3A). The olfactory fila pass through the cribriform plate to terminate in the olfactory bulb.
Olfactory receptor cells are true neurons because they originate embryologically from the central nervous system. Olfactory receptor cells undergo continuous turnover, with an average life span between 30 and 60 days. They are replaced by receptors arising from undifferentiated basal cells by mitotic division (Fig. 23-3A, C). Thus basal cells are stem cells that give rise to the receptor cells.
The supporting cells are columnar and extend from the lamina propria to the surface of the epithelium, where they end in short microvilli that extend into the overlying mucus (Fig. 23-3A, C, D). Nuclei of the sustentacular cells are found near the surface of the epithelium. These cells provide mechanical support for the olfactory receptor cells (Fig. 23-3B). In addition, they contribute secretions to the overlying mucus that may play a role in the binding or inactivation of odorant molecules.
A fourth and minor cell type, the microvillar cell, is found in the human olfactory epithelium (Fig. 23-3B). These cells have an apical process that projects into the mucus and a basal process that extends to the lamina propria. Although their function is unknown, they may be a second type of receptor neuron.
The lamina propria contains bundles of olfactory axons, blood vessels, fibrous tissue, and numerous Bowman glands (Fig. 23-3A). The serous secretions of the Bowman glands, combined with the secretions of the sustentacular cells, provide the mucus covering of the olfactory mucosa.
OLFACTORY TRANSDUCTION
Olfactory perception begins when volatile odor molecules are inhaled and contact the mucus layer that bathes the olfactory epithelium. This mucus is an aqueous solution of proteins and electrolytes. Odorants, particularly hydrophobic ones such as musk, cross the mucus by interacting with small, water-soluble proteins called odorant-binding proteins. These proteins are ubiquitous in the mucus layer.
After crossing the mucus, odor molecules bind to odorant receptors on the cilia of the olfactory receptor neurons, where transduction occurs (Fig. 23-4). The odorant receptors are membrane proteins belonging to a superfamily of G protein–coupled receptors. Binding of the odorant to one of as many as 1000 different types of odorant receptors leads to activation of a second messenger pathway involving an olfactory-specific G protein, which in turn activates adenyl cyclase to produce cyclic adenosine monophosphate (cAMP). The transient rise in ciliary cAMP opens a cyclic nucleotide–gated cation channel in the ciliary membrane, allowing cations to flow into the cell (Fig. 23-4). The flow of cations into the cell results in a gradual depolarization (generator potential) that travels down the dendrite to the soma of the olfactory receptor neuron. A sufficiently large depolarization initiates an action potential that travels along the axon to the olfactory bulb.
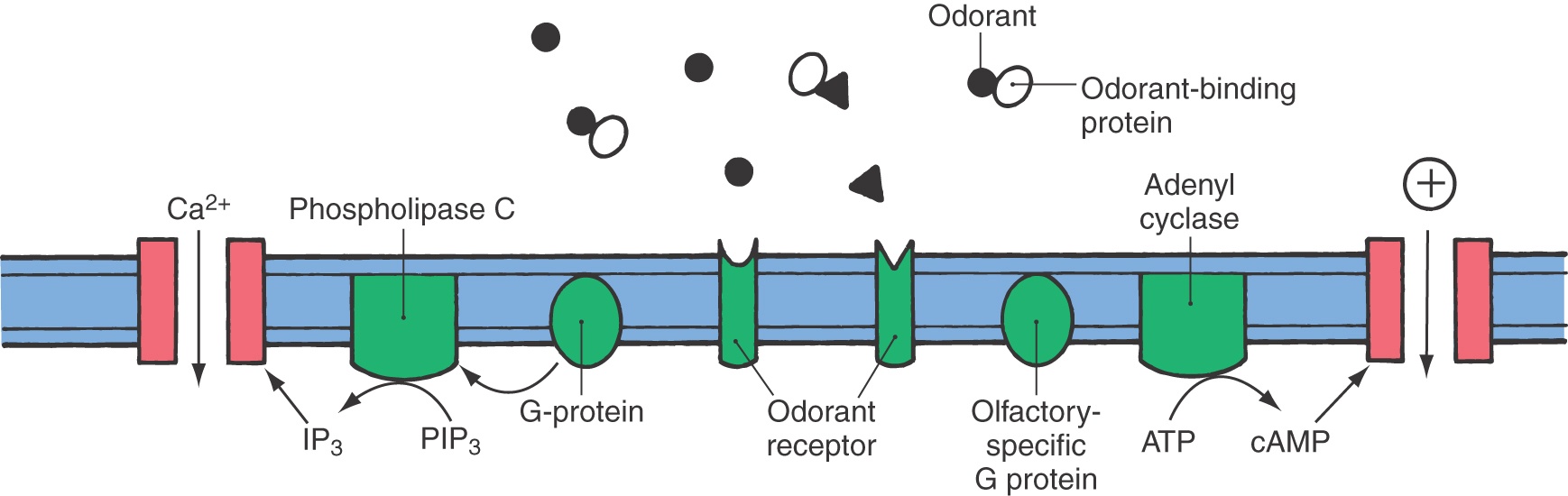
There is also evidence for another intracellular second messenger pathway in olfactory transduction. This pathway, involving inositol 1,4,5-trisphosphate (IP3), is thought to act either separately or with the cAMP pathway. In this pathway, binding of odorant to the receptor activates a G protein that in turn activates phospholipase C to produce IP3. The IP3 opens a channel in the ciliary membrane that permits calcium to enter the cell (Fig. 23-4). Current research further supports a role for cyclic guanosine monophosphate and carbon monoxide in olfactory signal transduction.
Studies suggest that olfactory discrimination begins in the olfactory epithelium and that a “receptor map” is used to encode complex qualities of a given odor. Receptors, which are tuned to specific structural features of a stimulus, appear not only to be selectively expressed within subsets of the olfactory neuron population but also to exhibit spatial organization. In fact, findings indicate that individual olfactory neurons express only one type of odorant receptor and that specific subtypes of odorant receptors preferentially distribute within one of four bilaterally symmetric zones of the olfactory epithelium. Such specificity permits olfactory information to be patterned for additional processing in the olfactory bulb.
CENTRAL OLFACTORY PATHWAYS
Olfactory Bulb
The olfactory bulb, a forebrain structure, is located on the ventral surface of the frontal lobe in the olfactory sulcus and is attached to the rest of the brain by the olfactory tract. The olfactory tract is an inclusive structure that contains fibers of the lateral olfactory tract, cells of the anterior olfactory nucleus, and fibers of the anterior limb of the anterior commissure. The latter part of the olfactory tract is the route through which many centrifugal fibers reach the olfactory bulb (see Fig. 23-7).
The olfactory bulb consists of five well-defined layers of cells and fibers, which give it a laminated appearance. From superficial to deep, these are the olfactory nerve layer, glomerular layer, external plexiform layer, mitral cell layer, and granule cell layer (Fig. 23-5).
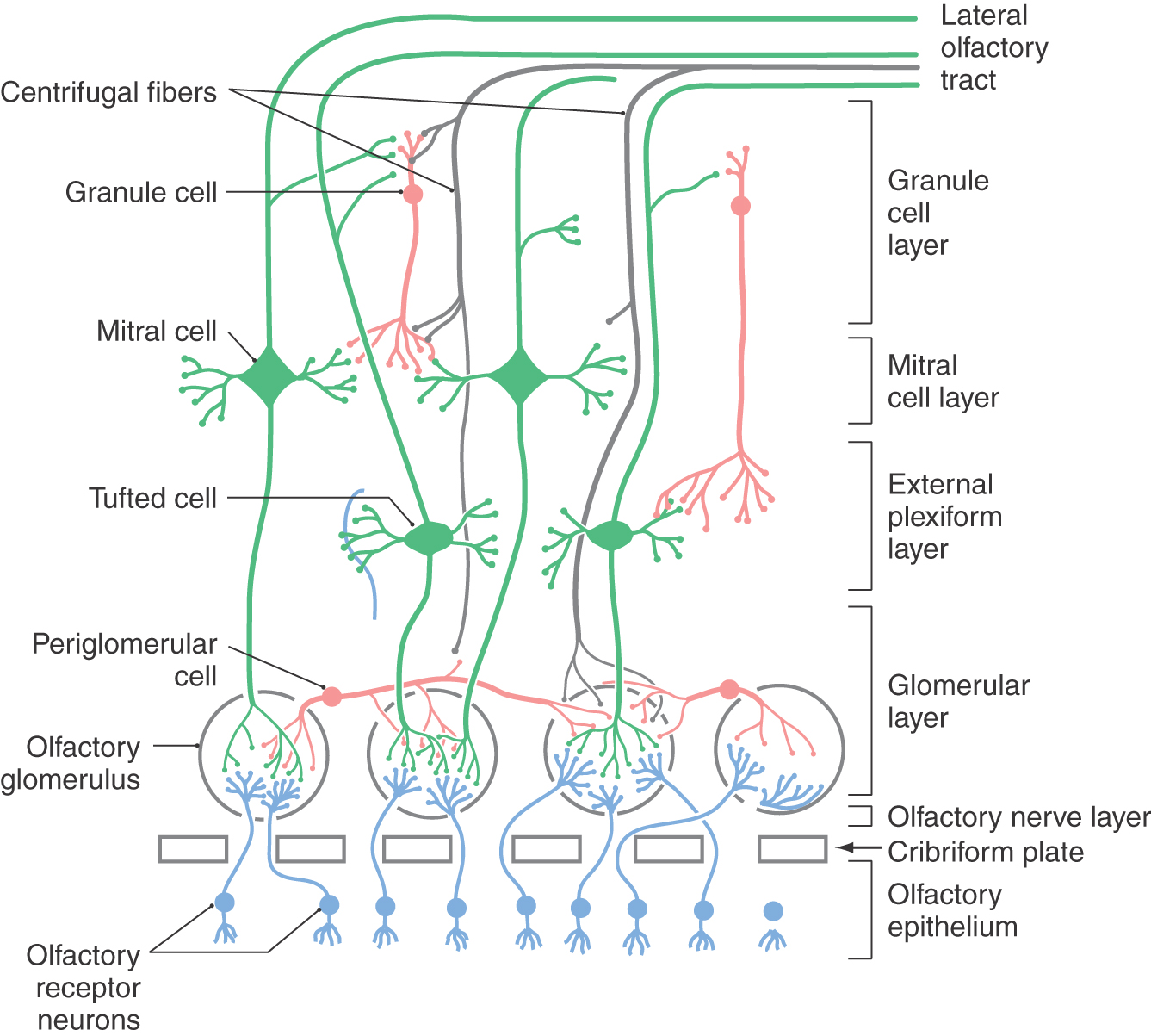
The afferent projections from the olfactory epithelium form the olfactory nerve layer on the surface of the olfactory bulb. These axons terminate exclusively in structures called olfactory glomeruli, which are found in the glomerular layer of the bulb (Figs. 23-5 and 23-6). The axon of each olfactory sensory neuron synapses in only one glomerulus. Interestingly, the terminations of these axons are arranged such that neurons expressing the same receptor subtype target the same few glomeruli. This suggests that each glomerulus receives input from only one type of receptor.
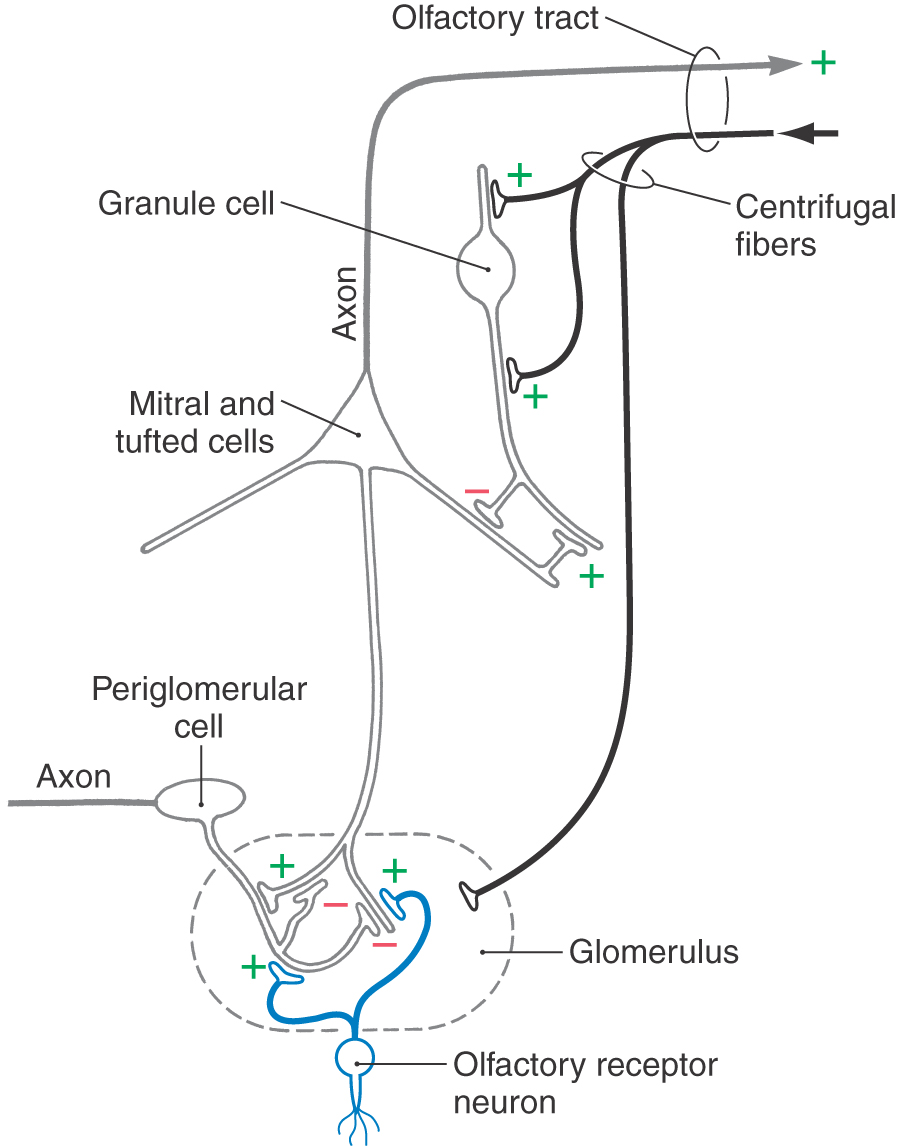
Glomeruli are the most prominent feature of the olfactory bulb. The core of an olfactory glomerulus is made up of the axons of olfactory receptor neurons, which branch and synapse on the bushy endings of the primary dendrites (apical dendrites) of mitral and tufted cells (Fig. 23-5). These two cells are functionally similar and together constitute the efferent neurons of the olfactory bulb. Adjacent to the glomerulus are small interneurons (juxtaglomerular cells), of which periglomerular cells are the principal type. This cell has short bushy dendrites that arborize extensively within a glomerulus and a short axon that distributes within a radius of about five glomeruli.
There is significant neural convergence at the level of the olfactory glomerulus; thousands of olfactory receptor neurons form excitatory (glutaminergic, carnosine) axodendritic synapses on mitral, tufted, and periglomerular cells (Fig. 23-6). The other major synaptic connections within the glomerulus are reciprocal and serial dendrodendritic synapses between mitral or tufted cells and periglomerular cells. It appears that the synapses of mitral and tufted cells onto periglomerular cells are excitatory (glutaminergic), whereas those of periglomerular cells onto mitral and tufted cells are inhibitory (GABAergic; GABA, for γ-aminobutyric acid).
The glomerular layer also receives input from other central nervous system areas via centrifugal afferents that use a wide variety of neurotransmitters and neuromodulators (Figs. 23-5 and 23-6). Noradrenergic centrifugal afferents from the locus ceruleus and serotonergic fibers from the raphe nuclei of the midbrain and rostral pons terminate in the glomeruli. Centrifugal fibers from the ipsilateral anterior olfactory nucleus and the diagonal band terminate in the periglomerular spaces, primarily on periglomerular cells. Excitatory amino acids, such as glutamate, are present in centrifugal fibers that arise in cortical structures.
The external plexiform layer is composed of the somata of tufted cells along with the primary and secondary dendrites (basal dendrites) of tufted and mitral cells and the apical dendrites of granule cells (Fig. 23-5). Within this layer, the apical dendrites of granule cells form reciprocal dendrodendritic GABAergic synapses with the secondary dendrites of tufted and mitral cells. These synapses modulate tufted and mitral cell output through lateral and feedback inhibition. Mitral and tufted cells, in turn, form excitatory (glutaminergic) synapses with granule cell dendrites (Fig. 23-6).
The mitral cell layer is a thin layer containing the large somata of mitral cells. In addition, the axons of tufted cells, granule cell processes, and centrifugal fibers traverse this layer (Fig. 23-5).
Internal to the mitral cell layer, the granular cell layer contains the cell bodies of granule cells, the principal interneuron of the olfactory bulb. This layer also contains primary and collateral axons of mitral and tufted cells and centrifugal afferents from the anterior olfactory nucleus, olfactory cortex, cells of the diagonal band, locus ceruleus, and raphe nucleus (Fig. 23-5). Granule cells lack axons, their only output being via dendrodendritic GABAergic synapses with mitral and tufted cells. In addition, granule cells receive numerous synaptic inputs from both mitral and tufted cell axon collaterals and centrifugal afferent fibers. Granule cells presumably modulate olfactory bulb activity via an inhibitory feedback loop that shuts down the activity of the mitral and tufted neurons.
Olfactory Bulb Projections
Axons of mitral and tufted cells emerge from the caudal portion of the olfactory bulb to form the lateral olfactory tract. Although glutamate is the major neurotransmitter of these efferent fibers, aspartate, dopamine, and substance P also are used. These fibers course caudally to terminate in areas on the ventral surface of the telencephalon, which are broadly defined as the olfactory cortex (Fig. 23-7). The principal areas making up the olfactory cortex are the anterior olfactory nucleus, olfactory tubercle, piriform cortex, anterior cortical amygdaloid nucleus, periamygdaloid cortex, and lateral entorhinal cortex. The olfactory cortex is an example of paleocortex, a phylogenetically older type of cortex that is less complex than the neocortex. Throughout most of its extent, the olfactory cortex has three cell layers, as opposed to the six layers characteristic of neocortex. A unique aspect of the olfactory system is that the olfactory bulb projects directly to the cortex. In other sensory systems, information reaches the cortex after a relay in the thalamus.
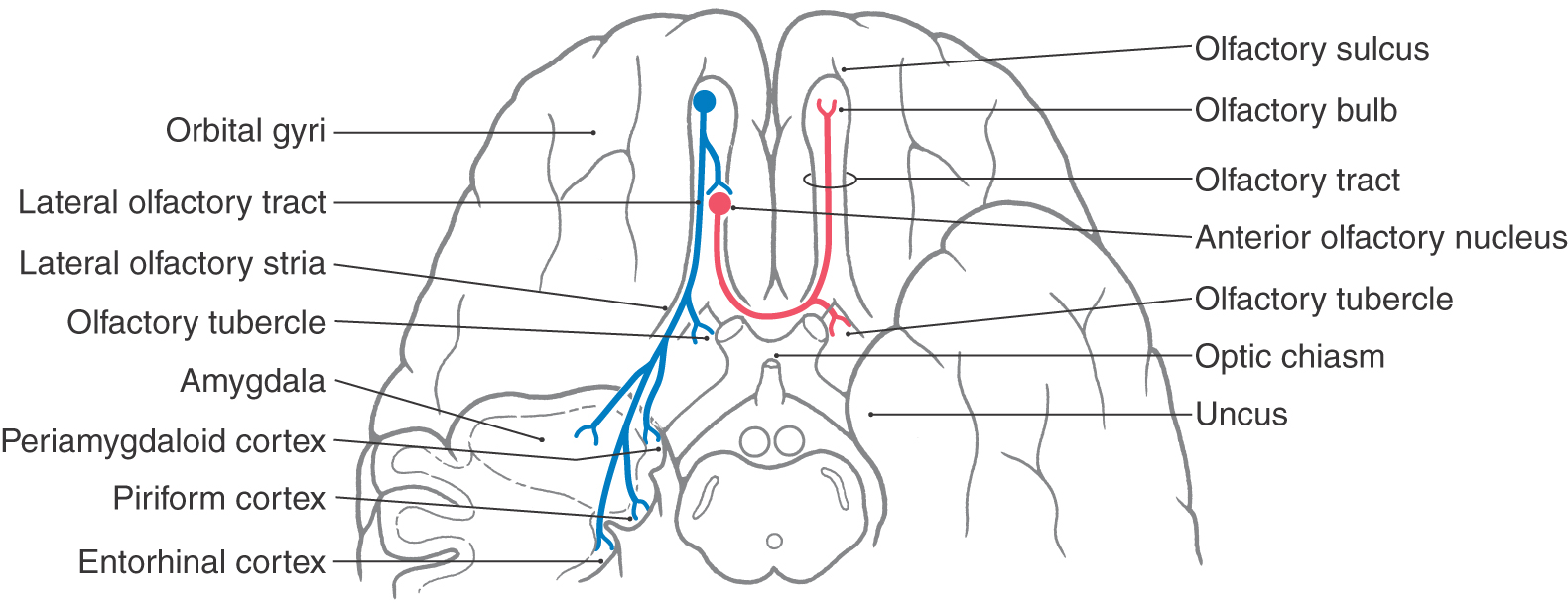
Lateral olfactory tract axons send collaterals to the anterior olfactory nucleus, to other areas of olfactory cortex, and to subcortical limbic structures. The major targets of the anterior olfactory nucleus are the olfactory bulbs bilaterally and the contralateral anterior olfactory nucleus (Figs. 23-7 and 23-8). The large numbers of interbulbar connections, via the anterior olfactory nucleus, suggest that interhemispheric processing of odors plays an important role in olfactory functions.
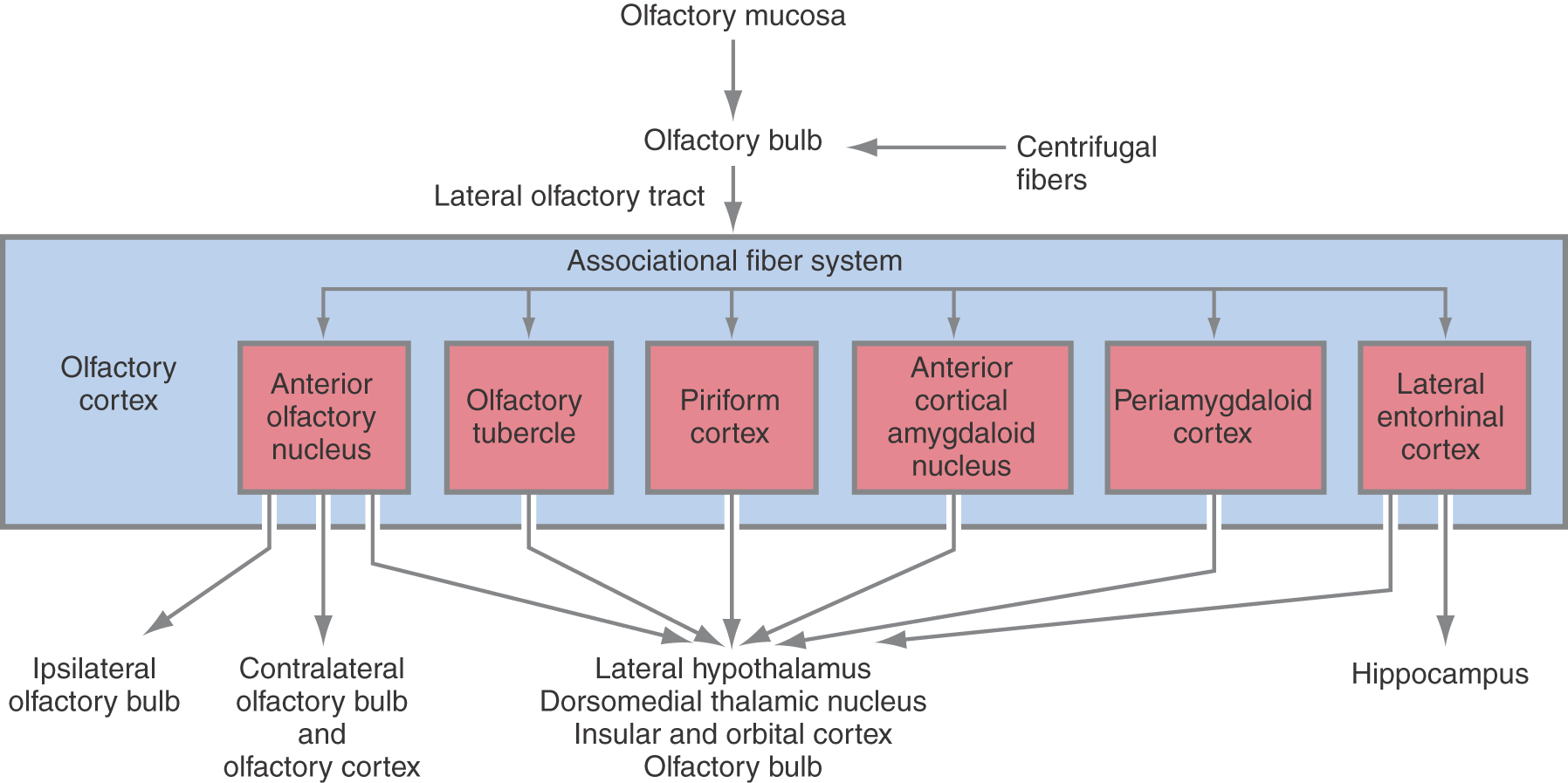
Figure 23-8. Major projections of the olfactory cortex.
Axons of the lateral olfactory tract course caudally in the form of the lateral olfactory stria to terminate in the olfactory tubercle and the piriform cortex (Fig. 23-7). The piriform cortex is a major component of the olfactory cortex. Fibers of the lateral olfactory tract also continue posteriorly to terminate in the anterior cortical amygdaloid nucleus, the periamygdaloid cortex (a part of the piriform cortex overlying the amygdala), and the lateral entorhinal cortex.
There is little evidence of a topographic projection from the olfactory bulb onto the various structures constituting the olfactory cortex. Mitral cells project to all areas of the olfactory cortex, whereas tufted cells terminate primarily in its anterior parts. However, each region of olfactory cortex is regarded as receiving input from all areas of the olfactory bulb.
Olfactory Cortex Projections
Cells of the olfactory cortex have reciprocal connections with other regions of the olfactory cortex (intrinsic or associational connections) and connections with regions outside the olfactory cortex (extrinsic connections) (Fig. 23-8). Most intrinsic connections arise from the anterior olfactory nucleus, piriform cortex, and lateral entorhinal cortex. As a group, these associational fibers distribute to all areas of the olfactory cortex (Fig. 23-8).
Extrinsic connections include extensive projections back to the olfactory bulb. These centrifugal fibers originate from most parts of the olfactory cortex, with the exception of the olfactory tubercle. As in other sensory systems, olfactory information is also relayed to the neocortex. This connection occurs through a direct projection from olfactory cortex to orbitofrontal and ventral agranular insular cortices or via a relay in the thalamus (Fig. 23-9). The latter pathway originates from cells in the olfactory cortex that project to the dorsomedial nucleus of the thalamus (Fig. 23-8). This neocortical representation of olfaction is important for discrimination and identification of odors, and lesions in this area, particularly the orbitofrontal cortex, result in the loss of these capabilities. Another noteworthy point is that the insular cortex and orbitofrontal cortex also receive taste input. The medial orbitofrontal cortex appears to play an especially important role in integrating olfactory, taste, and other food-related cues that produce the experience of flavor (Fig. 23-9).
In addition to neocortical projections, the olfactory cortex also sends fibers directly to the lateral hypothalamus and hippocampus. Those to the lateral hypothalamus arise primarily from the piriform cortex and anterior olfactory nucleus and are probably important for feeding behavior. The projection to the hippocampus arises from the entorhinal cortex and links olfactory input to centers concerned with learning and behavior.
Disorders of the Olfactory System
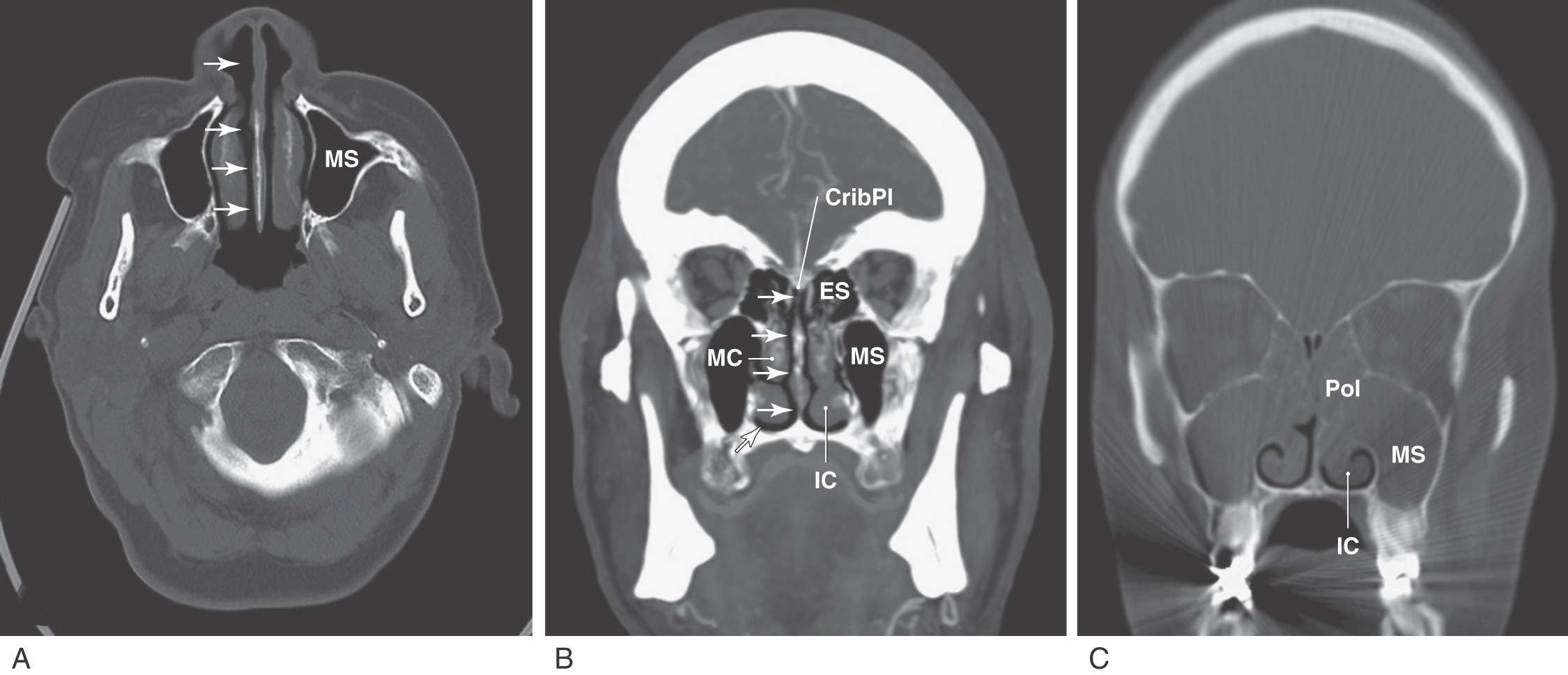
Disorders of smell are normally classified according to the type of loss experienced by the patient. The loss of smell (anosmia) or decreased sensitivity to odorants (hyposmia or olfactory hypesthesia) can be associated with nasal polyps (Fig. 23-10). Nasal polyps are noncancerous growths that occur in the nasal cavity or within the sinuses; a growth in a sinus may extend into the nasal cavity. Nasal polyps may result from inflamed mucous membranes or allergic reactions and can obstruct the nasal cavities or extend into the nares (the openings between the vestibule of the nasal cavity and the external environment) (Fig. 23-2). In these locations, polyps obstruct the flow of air and may cause a number of clinical problems, including a significant compromise of olfactory sense (Fig. 23-10).
More frequently, however, olfactory losses are experienced as a result of upper respiratory tract infections, sinus disease, and trauma. Nasal and paranasal diseases (rhinitis, sinusitis) may block the access of odorants to the olfactory epithelium, but blockage is not the sole cause because even when blockage is absent, olfactory dysfunction may be present. Frequently, treatment with systemic antiinflammatory drugs results in rapid attenuation of symptoms, suggesting that edema of the olfactory epithelium may be partly responsible for the olfactory dysfunction. In addition, the viruses associated with upper respiratory tract infections may permanently damage the olfactory epithelium. Although the process is not completely understood, it is thought that viral infection of an olfactory receptor cell may lead to cell death.
Head trauma can produce olfactory deficits by damaging central olfactory pathways or olfactory receptor axons as they pass through the cribriform plate. Boxers, for instance, may sustain damage to their olfactory senses as a result of blows to the head that produce shearing movements of the olfactory bulb relative to the cribriform plate. Transection of the thin axons comprising the olfactory fila may result in a partial or total loss of smell that may persist as a temporary or permanent condition. Fractures along the anterior base of the skull are also commonly observed after head trauma. Dural lacerations may cause drainage of cerebrospinal fluid into the paranasal sinuses and through the nose (rhinorrhea). Involvement of the anterior venous sinuses can further lead to the pooling of blood in the periorbital tissues and produce a characteristic “raccoon” or “panda bear” appearance. The notable lack of subconjunctival hemorrhage can be used to rule out the prospect of direct ocular trauma. In cases of anterior skull base fracture, there is always the threat of olfactory nerve damage as well as the possible complication of meningitis. To reduce the chance of bacterial infection, patients may receive treatment with antibiotics or undergo surgical measures to repair a severe meningeal breach.
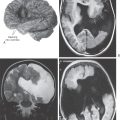
Neuroblastomas (malignant) and meningiomas (typically benign) may also compromise olfactory function. Neuroblastoma, a type of primitive neuroectodermal tumor deriving from primitive progenitor cells, is rare, occurs in children, and is most frequently localized to the frontoparietal lobe. The olfactory neuroblastoma (esthesioneuroblastoma) is believed to originate from olfactory neuroepithelial cells in the nasal cavity and to contribute to nasal obstruction and epistaxis (nosebleed) in affected individuals. Olfactory groove meningiomas, however, arise from arachnoid cells along the cribriform plate (see Fig. 7-9). Like medial sphenoid wing and other parasellar meningiomas, these neoplasms may elicit multiple sensory and motor disturbances. In addition to deficits in smell, visual abnormalities may be detected as the tumor compresses the optic nerve and causes blindness, optic atrophy, and contralateral papilledema (Foster Kennedy syndrome). Extension of the mass into the cavernous sinus may further produce extraocular palsies and facial numbness.
Olfactory losses are also the result of excessive smoking and prolonged use of cocaine. However, reports also indicate instances in which healthy individuals with otherwise normal olfactory acuity are unable to perceive the odor of a particular compound or class of compounds. This phenomenon, termed specific anosmia, is a condition that may be inherited as an autosomal recessive trait and is likely to be explained by the absence of a specific odorant receptor. Conversely, increased olfactory acuity (hyperosmia or olfactory hyperesthesia) has been described in conjunction with migraine and hysteria. Occurrences of hyperosmia have also been found in certain psychotic states and with certain types of substance abuse.
The chief complaint of most patients with chemosensory disturbances is the loss or alteration of taste. However, clinical studies reveal that in all but a small number of patients, the dysfunction actually resides in the olfactory system. The reason for this discrepancy is that most people confuse taste with flavor. In the evaluation of a patient with a potential olfactory lesion, it is important to note whether deficits exist in the individual’s ability to perceive or the ability to recognize a smell. The detection of an odorous substance indicates the integrity of the peripheral nerve and its pathway. Identification of the odor reveals intact cortical function. If the patient is aware of a smell but cannot recall the name of the scent (olfactory agnosia), the disorder is likely to reside at higher levels of the sensory system. A second relevant point pertains to the fact that olfactory losses may be manifested unilaterally or bilaterally. Unilateral deficiencies are typically observed subsequent to nasal cavity disease or after tumor-associated compression injury of one olfactory bulb or tract. Bilateral anosmia is usually sustained in response to head injury or the common cold. However, the absence of smell from either nostril has been noted in advanced cases of olfactory groove meningioma.
Age-related declines in olfactory function are common even in healthy persons. Typically, the loss occurs gradually, and the patient often fails to notice these changes. These gradual changes can affect the palatability of foods in the elderly. Olfactory dysfunction is also encountered in neurodegenerative diseases, such as Alzheimer and Parkinson diseases, or in Huntington chorea. These neurodegenerative diseases involve central olfactory pathways and produce marked reduction in the affected person’s olfactory capabilities. Most notably, these olfactory deficits appear very early in the course of the disease and may be among its first manifestations.
Disorders of olfaction are also associated with epilepsy and various depressive and psychiatric disorders, such as schizophrenia and Korsakoff psychosis. The patients frequently experience parosmia (dysosmia), a distortion in a smell experience or the perception of a smell when no odor is present (olfactory hallucination or phantosmia). Such episodes, resulting presumably from abnormal sequences of neuronal activity, can be elicited by an irritative lesion of the anterior medial temporal lobe, hippocampus, amygdala, or medial dorsal thalamic nucleus. In the case of epilepsy, focal (partial) seizure activity in the uncal region of the temporal lobe has been found to correlate with the formation of repugnant or disagreeable olfactory auras (cacosmia). This type of attack, known as an uncinate fit, may occur in people who are genetically predisposed to seizure-like discharge or in individuals who demonstrate tumor growth, a vascular disorder, head trauma, infection, drug use, or substance withdrawal.
TASTE RECEPTORS
The taste experiences of sweet, salty, sour, bitter, and umami result from an interaction between gustatory stimuli and receptor cells located in sensory organs called taste buds. Although they are found throughout the oropharyngeal cavity, taste buds are most obvious on the tongue, where they are ovoid structures with a constriction at their apical end.
Each taste bud contains 40 to 100 cells. It is now widely accepted that there are functionally distinct populations of cells within the taste bud. Type I cells mainly perform glial functions. Type II or receptor cells possess G protein–coupled receptors for bitter, sweet, and umami compounds. Type III or presynaptic cells express synapse-related proteins and contain conventional synapses. These cells do not have G protein–coupled taste receptors but do express candidate acid transducer channels that appear to play a role in the detection of sour stimuli. Type IV cells, or basal cells, are progenitor cells.
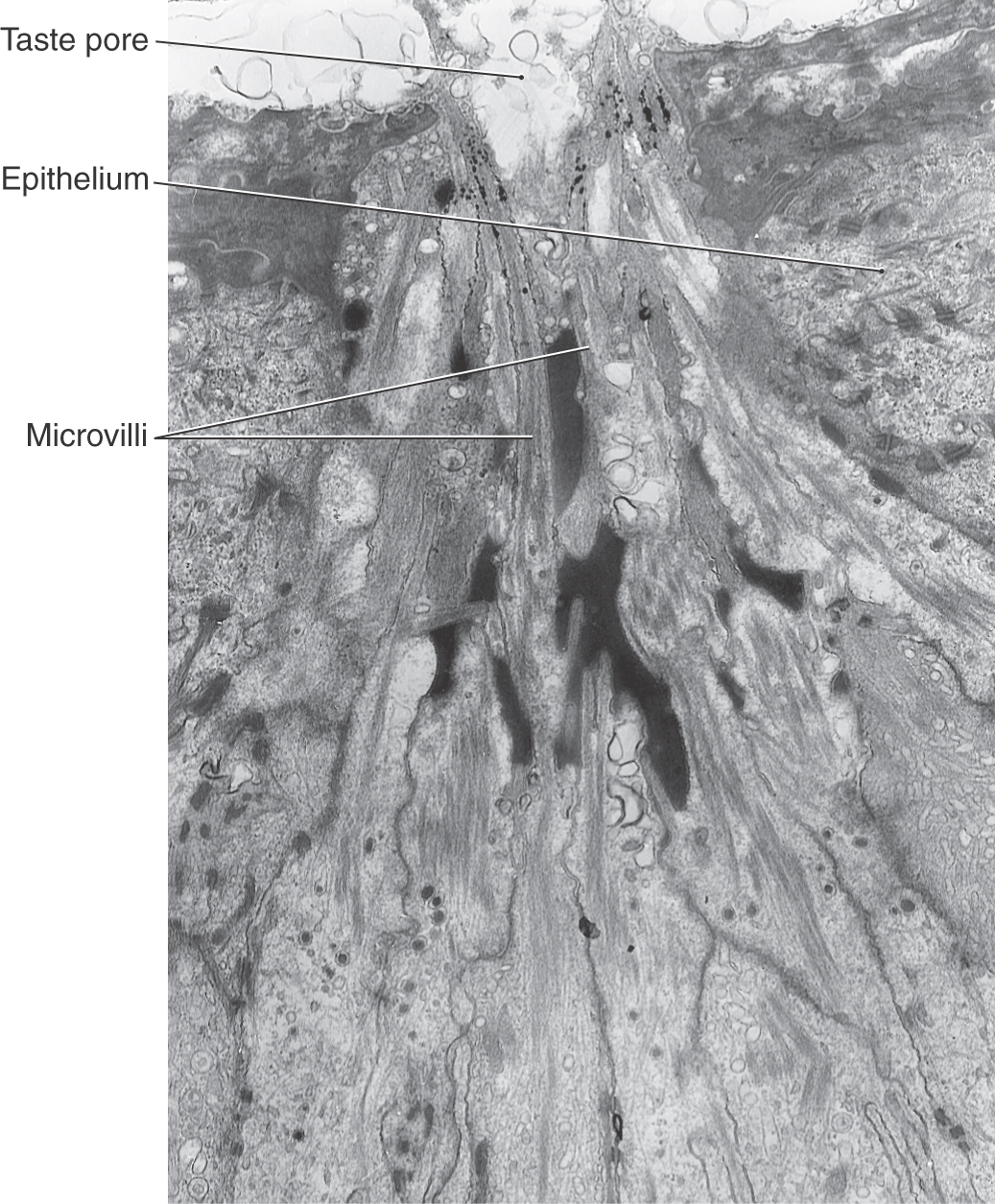
Taste cells extend from a basal lamina to the surface of the epithelium. The apical ends of these cells are covered with microvilli of variable lengths that extend into a taste pore. The pore forms a pocket to permit contact between the microvilli of the taste cell and the external milieu (Figs. 23-11 and 23-12). Numerous junctional complexes located between the apices of taste cells restrict access of stimuli to the microvilli, where taste transduction occurs. The taste pore is filled with a protein-rich substance through which substances must pass to reach the taste cell microvilli. Taste cells undergo a continuous process of turnover, having a life span of 10 to 14 days. New taste cells are thought to arise from polygonal basal cells located in basolateral areas of the taste bud. These cells are not involved in taste transduction.
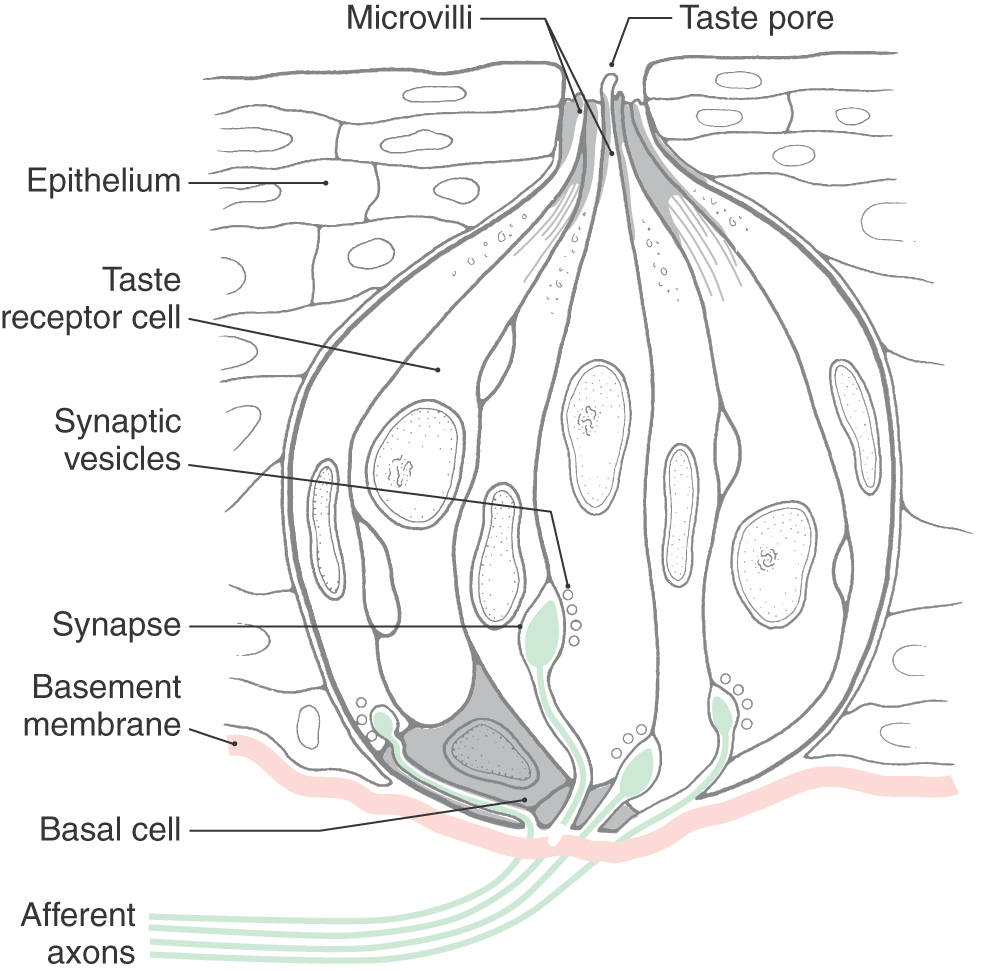
In response to taste stimulation, receptor cells secrete ATP via gap junction hemichannels. This taste neurotransmitter excites adjacent type III cells, which have been shown to form morphologically identifiable synapses with postsynaptic structures and to contain proteins involved in vesicular exocytosis. When activated, type III cells release serotonin and norepinephrine. Taste afferent fibers form the postsynaptic element of a chemical synapse near the base of the taste cell.
Afferent fibers penetrate the basement membrane and then branch within the base of the taste bud (Fig. 23-11). Each taste bud is typically innervated by more than one afferent fiber, and an individual fiber may innervate multiple taste buds.
DISTRIBUTION OF TASTE RECEPTORS
Lingual Taste Buds
Taste buds are found, in variable numbers, on the human tongue, palate, pharynx, and larynx. On the tongue, taste buds are located exclusively in specialized structures called papillae, of which there are three types (Fig. 23-13A). Taste buds on the anterior two thirds of the tongue reside in mushroom-shaped fungiform papillae (Fig. 23-13B). These structures are scattered among the more numerous nongustatory filiform papillae distributed over the surface of the tongue. The size, shape, and number of fungiform papillae vary widely, and usually two to four taste buds are found in the dorsal epithelium of each. The vallate (circumvallate) papillae are located on the dorsal surface of the tongue at the junction of the oral and pharyngeal cavities (Fig. 23-13A). There are 8 to 12 vallate papillae, each of which is composed of a central papilla surrounded by a cleft containing taste buds in its epithelium (Fig. 23-13C). A single foliate papilla on each side of the tongue appears as a series of clefts along the lateral margin of the tongue (Fig. 23-13A). Each is composed of two to nine clefts, with five being the most common number. Taste buds in foliate papillae are also located in the epithelium that lines the clefts (Fig. 23-13D).
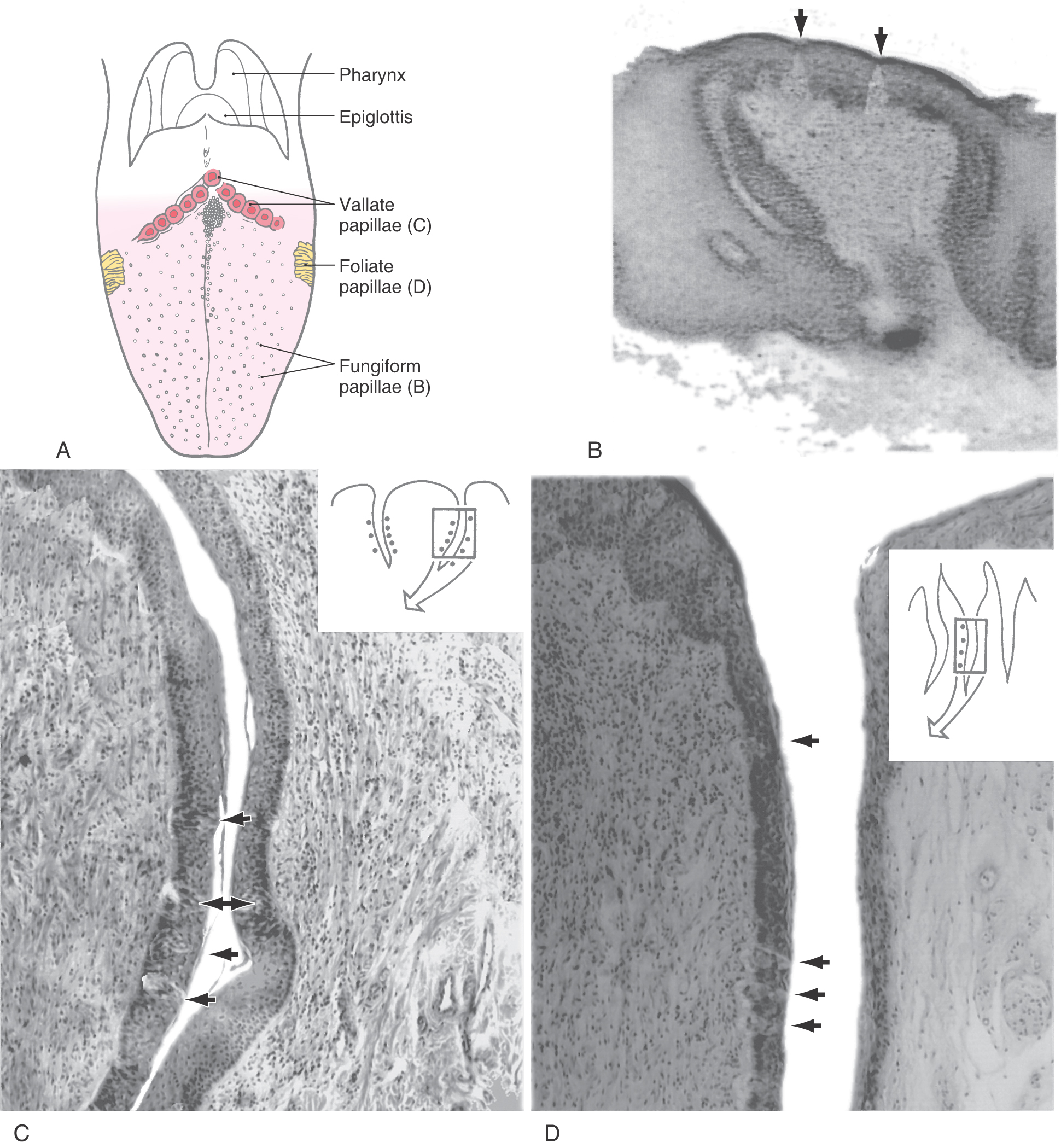
Associated with both the vallate and foliate papillae are the von Ebner lingual salivary glands. These glands drain into the base of the clefts and influence their microenvironment. Taste stimulation of vallate and foliate papillae influences the secretions of the von Ebner glands via circuits located in the brainstem.
It was once believed that different regions of the tongue were specialized for the detection of particular taste qualities. It is now known that all taste qualities are detected in all regions of the tongue, although sensitivity to the different taste qualities and taste transduction mechanisms may vary by tongue region.
Extralingual Taste Buds
Additional taste buds are located on the human soft palate, oral and laryngeal pharynx, larynx, and upper esophagus. Extralingual taste buds are not located in papillae but rather are situated in the epithelium. Palatal taste buds are located at the juncture of the hard and soft palates and on the soft palate. Laryngeal taste buds are found on the laryngeal surface of the epiglottis and adjacent aryepiglottic folds. The number of extralingual taste buds is substantial, and they may contribute to the taste experience. Stimulation of some extralingual taste buds, particularly those near the larynx, elicits brainstem-mediated reflexes that prevent accidental aspiration of ingested materials.
TASTE TRANSDUCTION
As research has begun to clarify the mechanisms underlying taste transduction, it has become clear that individual taste qualities or even individual taste compounds use several transduction mechanisms. In general, taste transduction is initiated when soluble chemicals diffuse through the contents of the taste pore and interact with receptors located on the exposed apical microvilli of the taste cells. Several different receptor types have recently been cloned, including a novel variant of the metabotropic glutamate receptor that functions as an umami taste receptor and G protein–coupled receptors that function as bitter receptors. The interaction of the chemical stimulus with the taste cell receptor results in either depolarization or hyperpolarization of the taste cell microvilli. It is now generally accepted that depolarizing potentials of sufficient magnitude result in action potential generation within taste cells, which in turn produce an increase in intracellular calcium, either by the release of calcium from internal stores or by the activation of voltage-gated calcium channels located in the basolateral membrane of taste cells (Fig. 23-14). This calcium release results in a release of chemical transmitters at the afferent synapse, which in turn leads to an action potential in the afferent fiber.
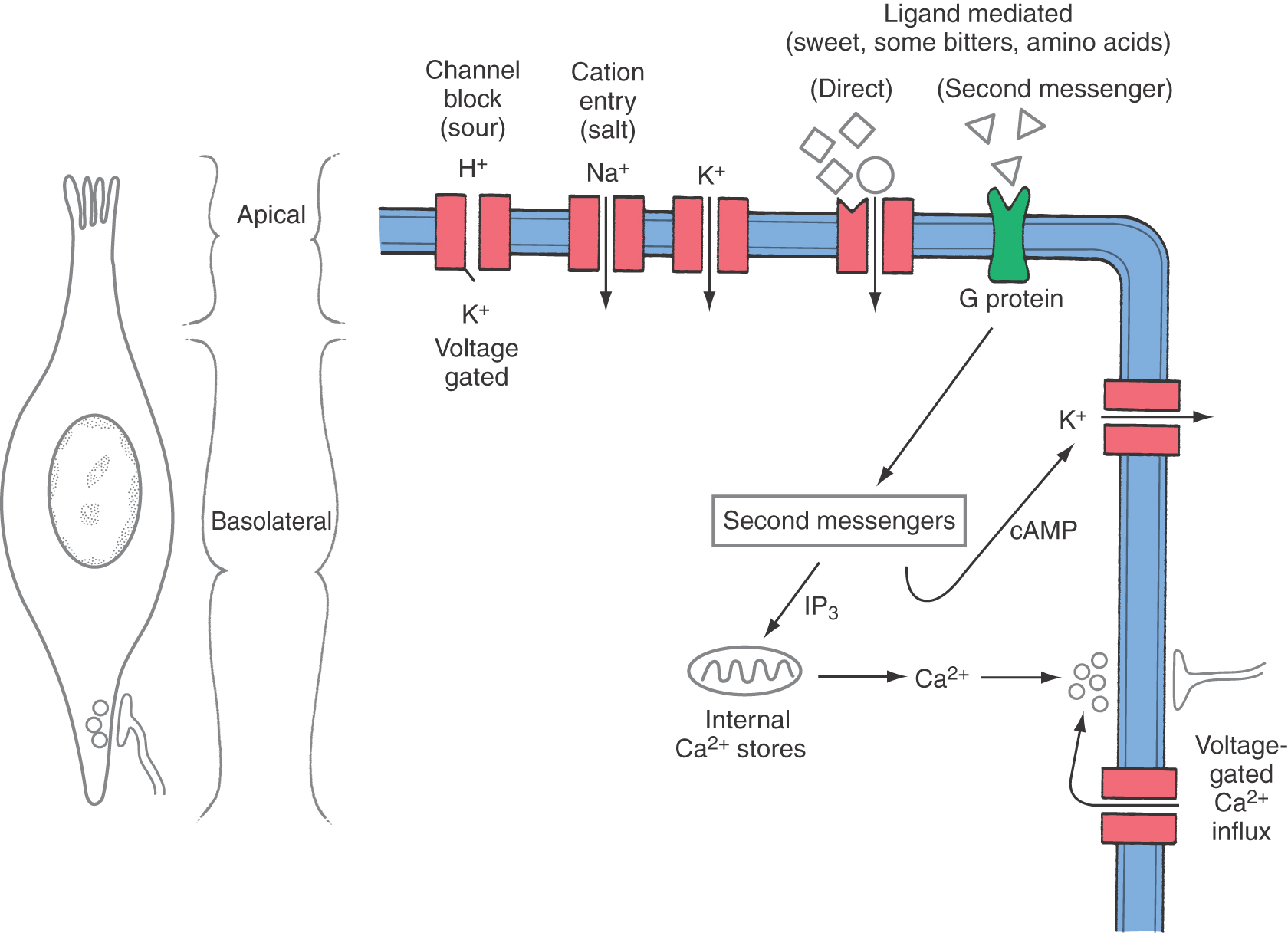
Transduction of stimuli leading to salty and perhaps some sour and bitter tastes appears to be the result of a direct interaction of these tastants with specific ion channels located in the apical membrane of taste cells. One mechanism for the transduction of sodium salts such as sodium chloride involves movement of sodium into the taste cell through apically located amiloride-sensitive cation channels. Similar mechanisms have been proposed for potassium salts (Fig. 23-14). One pathway put forward for transduction of some sour and bitter stimuli is blockage of apical voltage-sensitive potassium channels. At the resting potential, there is a small outward potassium current through the apical membranes of taste cells. Protons provided by sour stimuli such as hydrochloric acid are thought to block this outward current, causing the cell to depolarize. Interestingly, more recent evidence points to the importance of the membrane-permeant, uncharged acetic acid moiety (CH3COOH) as the active stimulus for acid taste. Such findings emphasize the role of intracellular acidification in sour transduction.
Sweet and some bitter-tasting compounds are transduced by receptors that activate intracellular G protein–mediated second messenger pathways (Fig. 23-14). Binding of sweet-tasting compounds such as sucrose to apically located receptors stimulates an adenyl cyclase–cAMP second messenger pathway that closes basolateral potassium channels, leading to depolarization of the taste receptor cell. Additional mechanisms for sweet transduction have been proposed, and there is good evidence for multiple sweet receptor types. A different second messenger pathway (IP3) that releases calcium from intracellular stores has been shown in conjunction with some bitter compounds but may also play a role in the perception of sweet. Alkaloids, glucosides, and some amino acids are known to convey a bitter taste.
Evidence exists for several possible transduction pathways for amino acids. One pathway involves binding of amino acids by receptors that are directly coupled to cation channels having properties similar to those of the nicotinic acetylcholine receptor. Another receptor activates a G protein–dependent increase in the second messengers cAMP and IP3. For umami transduction, it is hypothesized that a glutamate receptor activates a G protein that stimulates phosphodiesterase, causing a reduction in intracellular cAMP and subsequent changes in receptor cell activity.
PERIPHERAL TASTE PATHWAYS
The afferent fibers of first-order taste neurons (special visceral afferent [SVA/VA]) innervating oropharyngeal taste buds travel in the facial (VII), glossopharyngeal (IX), and vagus (X) nerves (Fig. 23-15). The chorda tympani branch of the facial nerve innervates taste buds in the fungiform papillae on the anterior two thirds of the tongue and in the most anterior clefts of the foliate papillae. The greater superficial petrosal nerve, also a branch of the facial nerve, innervates taste buds on the soft palate. The cell bodies of facial nerve fibers subserving taste are located in the geniculate ganglion, and their central processes enter the brainstem at the pontomedullary junction in the intermediate nerve, which is actually a part of the facial nerve. These primary afferent taste fibers enter the solitary tract, travel caudally, and terminate on cells of the surrounding solitary nucleus (Fig. 23-15).
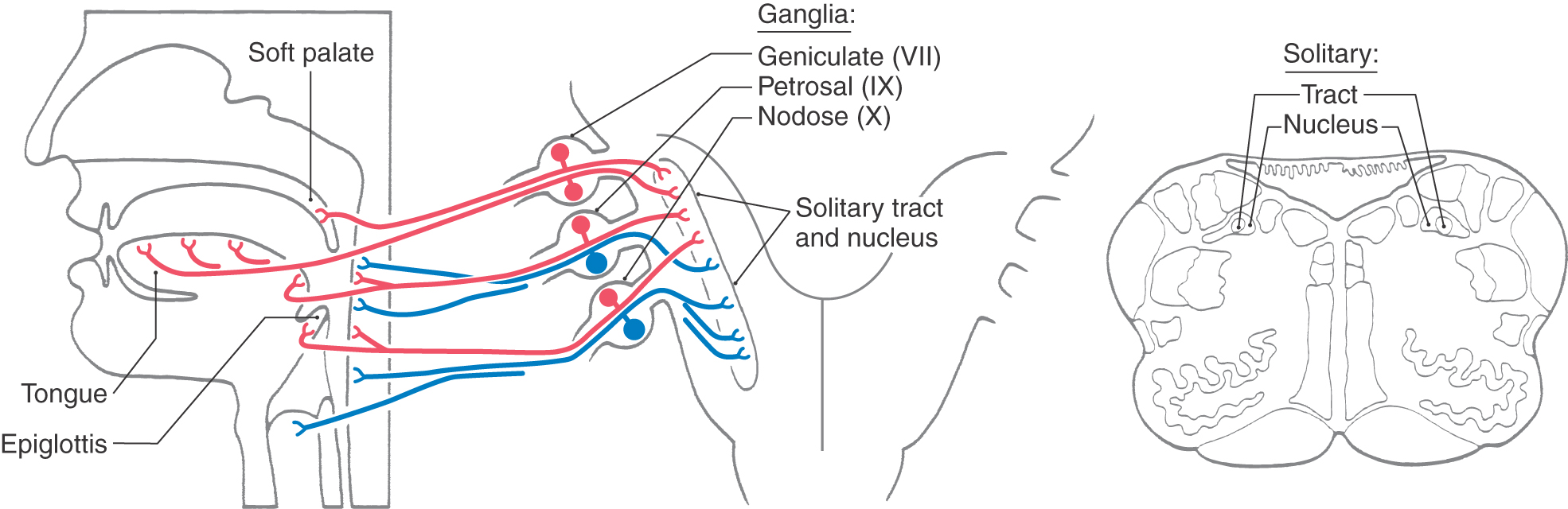
 Figure 23-15.
Figure 23-15. Taste buds located in the vallate papillae and posterior clefts of the foliate papillae are innervated by the lingual-tonsillar branch of the glossopharyngeal nerve (cranial nerve IX). Those located on the epiglottis and esophagus are innervated by the superior laryngeal nerve, a branch of the vagus nerve (cranial nerve X) (Fig. 23-15). Taste fibers in cranial nerves IX and X have their cell bodies of origin in the inferior ganglia (petrosal and nodose, respectively) of these cranial nerves (Fig. 23-15). The central processes of these fibers, like those of the facial nerve, enter the medulla, descend in the solitary tract, and terminate on neurons in the adjacent solitary nucleus (Fig. 23-15).
CENTRAL TASTE PATHWAYS
The solitary nucleus is the principal visceral afferent nucleus of the brainstem. On the basis of functional characteristics, it is divided into a rostral (gustatory) nucleus and a caudal (visceral or cardiorespiratory) nucleus. Taste fibers traveling in cranial nerves VII, IX, and X terminate primarily in the rostral portions of the solitary nucleus because this region contains most of the second-order neurons in the taste pathway (Fig. 23-15). Although there is considerable overlap in the distribution of the terminals of the primary taste cranial afferents from these three nerves in the gustatory nucleus, information derived from different areas of the tongue is spatially segregated. General visceral afferent fibers of the vagus and those that travel in the glossopharyngeal nerve terminate in the caudal part of the solitary nucleus (Fig. 23-15). These visceral fibers are involved in the central control of respiration, cardiac function, and certain aspects of swallowing.
Axons arising from second-order taste neurons in the gustatory nucleus ascend in association with the ipsilateral central tegmental tract and terminate in the parvicellular division of the ventral posteromedial nucleus of the thalamus (VPMpc), medial to the head representation (Fig. 23-16). Axons from these neurons in the VPMpc travel through the ipsilateral posterior limb of the internal capsule to terminate in the inner portion of the frontal operculum and anterior insular cortex and in the rostral extension of Brodmann area 3b on the lateral convexity of the postcentral gyrus (Fig. 23-16). This pathway (solitary nucleus → VPMpc → cortex) is responsible for the discriminative aspects of taste and, in contrast to other sensory pathways, is exclusively ipsilateral.
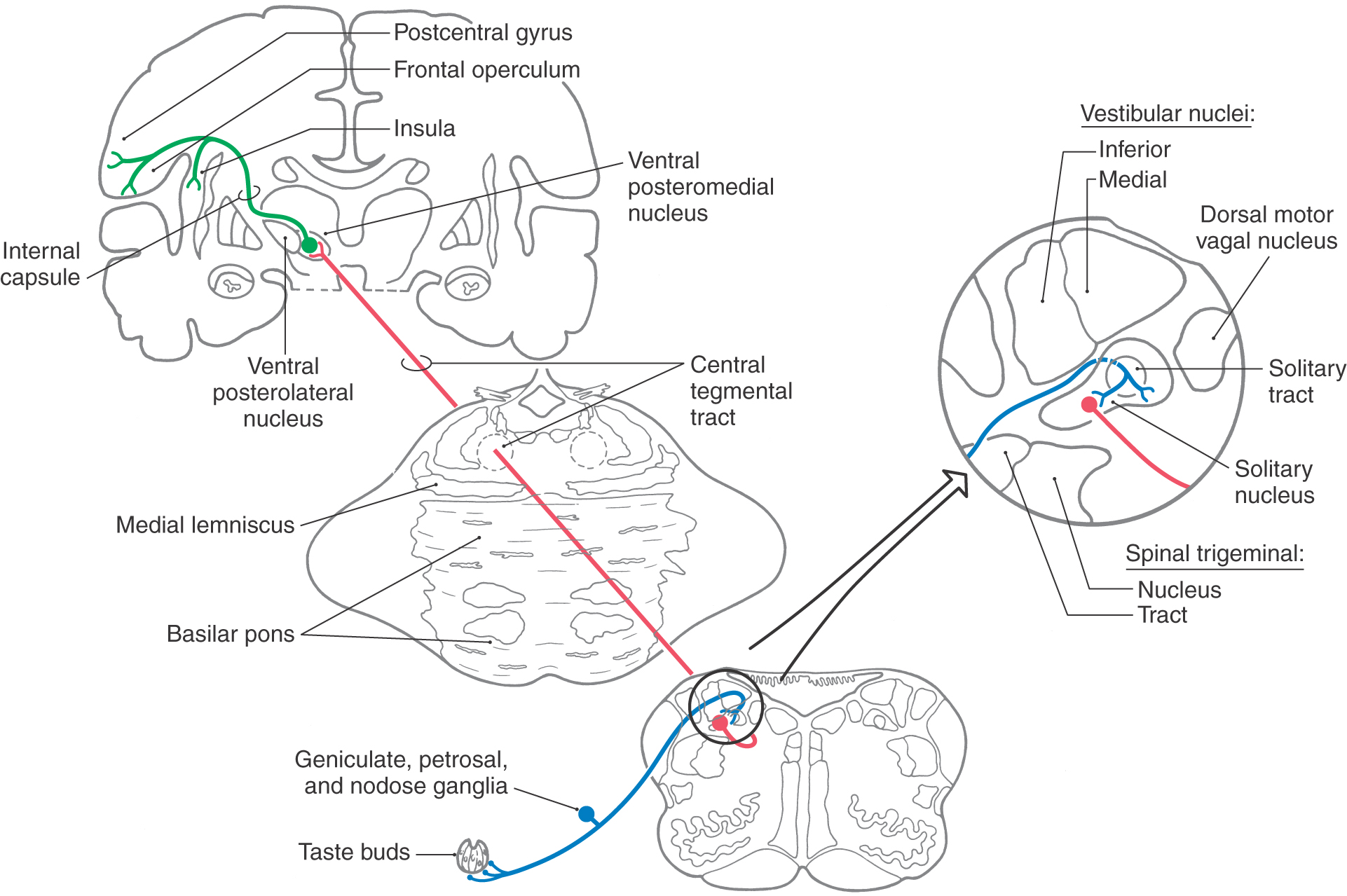
Physiologic studies in primates indicate that there is an additional region of the cortex that processes taste information. The lateral posterior orbitofrontal cortex receives inputs from primary taste cortex and acts as a site of integration for taste, olfactory, and visual cues associated with the ingestion of foods. Recent data suggest that cells in this area are involved in the appreciation of flavor, food reward, and the control of feeding. Taste-responsive cells have also been found in the primate amygdala and hypothalamus. These cells do not respond exclusively to taste, and their connecting pathways and role in taste-mediated behavior are not fully understood. Taste information is also relayed from cells in the solitary nucleus into medullary reflex connections that influence salivary secretion, mimetic responses, and swallowing.
Disorders of the Gustatory System
The chorda tympani branch leaves the facial nerve just distal to the geniculate ganglion. Consequently, lesions of the root of the seventh cranial nerve or tumors in the internal auditory meatus, such as a vestibular schwannoma, sometimes called an acoustic neuroma (see Fig. 22-10), will result in loss of taste perception from the anterior two thirds of the tongue on the ipsilateral side. Accompanying this deficit are paralysis of the ipsilateral facial muscles, hyperacusis (paralysis of the stapedius muscle), and impaired secretion of the nasal and lacrimal glands and of submandibular and sublingual salivary glands. Damage just distal to the geniculate ganglion may or may not result in taste loss, depending on the origin of chorda tympani branches, but an ipsilateral facial paralysis will be seen (see Fig. 14-14).
The complete loss of taste (ageusia) is rarely encountered, in part because of the large numbers of nerves that relay taste information to the central nervous system. Rarely does a patient have bilateral injury to all the nerves innervating the oropharyngeal region. More frequently a patient has hypogeusia, decreased taste sensitivity, or parageusia (dysgeusia), distortions in the perception of a taste. One of the most frequent causes of taste disturbances is drug use. A wide array of medications can affect taste function, the most frequent complaint being dysgeusia. Like olfactory disorders, taste disorders are associated with head trauma, viral infections, and various psychiatric disorders. Notable impairments of taste have been documented in Bell palsy (see Chapter 25). It has been suggested that herpes simplex virus may be a causative factor of this disease and that administration of prednisone may reduce potential damage from nerve swelling in the facial canal. Taste changes have also been documented in the elderly as numbers of taste buds decrease and taste transduction mechanisms become less effective. These changes are often accompanied by alterations in eating habits. For example, a person may use more sugar in his or her coffee to compensate for reduced acuity of sweet stimuli. Taste disturbances are similarly known to occur in cancer patients who receive radiation therapy and chemotherapy and in patients with diabetes, in whom losses are progressive.
Another consideration is saliva, an important medium for the relay of chemical information to taste receptors. Diseases that affect the production of saliva may have an impact on taste quality. Individuals with cystic fibrosis, for instance, have been described as experiencing increases in taste sensitivity that may be related to the hyperviscosity of their saliva. Other factors contributing to altered taste sensation include oropharyngeal tumors, which can compromise the function of the chorda tympani or lingual nerves, and foci of seizure activity in central taste processing areas, which can trigger unpleasant taste sensations (cacogeusia). Although experienced less frequently than olfactory auras, gustatory hallucinations have been elicited by electrical stimulation of the frontal and parietal opercula as well as the hippocampus and amygdala.
Sources and Additional Reading
Anholt RRH. Molecular neurobiology of olfaction. Crit Rev Neurobiol. 1993;7:1–22.
Kinnamon SC. Taste transduction: A diversity of mechanisms. Trends Neurosci. 1988;10:491–496.
Kinnamon SC. A plethora of taste receptors. Neuron. 2000;25:507–510.
Lewcock JW, Reed RR. ORs rule the roost in the olfactory system. Science. 2003;302:2078–2079.
Speielman AI. Chemosensory function and dysfunction. Crit Rev Oral Biol Med. 1998;9:267–291.
Victor M, Ropper AH. Adams and Victor’s Principles of Neurology. 7th ed New York: McGraw-Hill; 2001.