[level-membership-for-basic-science-category]
CHAPTER 68 Liver
The liver is the largest of the abdominal viscera, occupying a substantial portion of the upper abdominal cavity. It occupies most of the right hypochondrium and epigastrium, and frequently extends into the left hypochondrium as far as the left lateral line (Fig. 68.1). As the body grows from infancy to adulthood the liver rapidly increases in size. This period of growth reaches a plateau around 18 years and is followed by a gradual decrease in the liver weight from middle age. The ratio of liver to body weight decreases with growth from infancy to adulthood. The liver weighs approximately 5% of the body weight in infancy and it decreases to approximately 2% in adulthood. The size of the liver also varies according to sex, age and body size. It has an overall wedge shape, which is in part determined by the form of the upper abdominal cavity into which it grows. The narrow end of the wedge lies towards the left hypochondrium, and the anterior edge points anteriorly and inferiorly. The superior and right lateral aspects are shaped by the anterolateral abdominal and chest wall as well as the diaphragm. The inferior aspect is shaped by the adjacent viscera. The capsule is no longer thought to play an important part in maintaining the integrity of the shape of the liver.
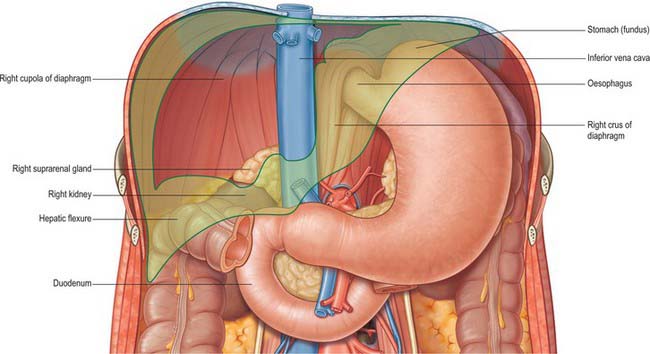
Fig. 68.1 The ‘bed’ of the liver. The outline of the liver is shaded green. The central bare area is unshaded.
SURFACES OF THE LIVER
The liver is usually described as having superior, anterior, right, posterior and inferior surfaces, and has a distinct inferior border (Figs 68.2, 68.3). However, the superior, anterior and right surfaces are continuous and no definable borders separate them. It would be more appropriate to group them as the diaphragmatic surface, which is mostly separated from the inferior, or visceral surface, by a narrow inferior border. At the infrasternal angle the inferior border is related to the anterior abdominal wall and is accessible to examination by percussion, but is not usually palpable. In the midline, the inferior border of the liver is near the transpyloric plane, about a hand’s breadth below the xiphisternal joint. In women and children the border often projects a little below the right costal margin.
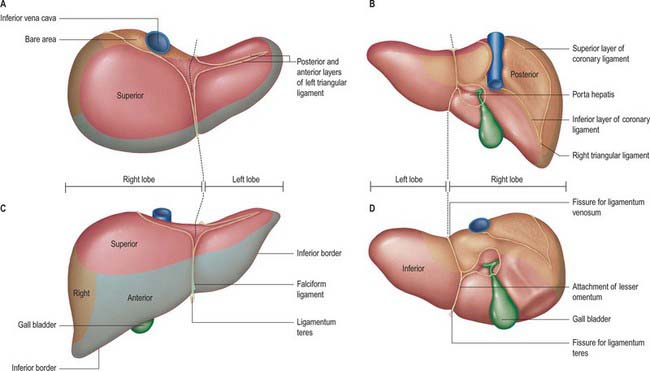
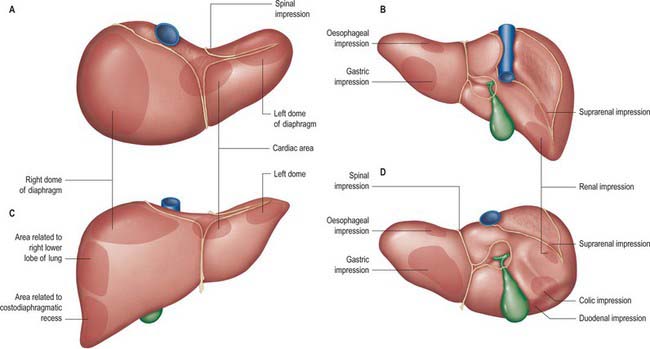
Fig. 68.3 Relations of the liver. A, superior view; B, posterior view; C, anterior view; D, inferior view.
The posterior surface is convex, wide on the right, but narrow on the left. A deep median concavity corresponds to the forward convexity of the vertebral column close to the attachment of the ligamentum venosum. Much of the posterior surface is attached to the diaphragm by loose connective tissue, forming the triangular ‘bare area’. The inferior vena cava lies in a groove or tunnel in the medial end of the ‘bare area’. To the left of the caval groove the posterior surface of the liver is formed by the caudate lobe, and covered by a layer of peritoneum continuous with that of the inferior layer of the coronary ligament and the layers of the lesser omentum. The caudate lobe is related to the diaphragmatic crura above the aortic opening and the right inferior phrenic artery, and separated by these structures from the descending thoracic aorta.
The fissure for the ligamentum venosum separates the posterior aspect of the caudate lobe from the main part of the left lobe. The fissure cuts deeply in front of the caudate lobe and contains the two layers of the lesser omentum. The posterior surface over the left lobe bears a shallow impression near the upper end of the fissure for the ligamentum venosum that is caused by the abdominal part of the oesophagus. The posterior surface of the left lobe to the left of this impression is related to part of the fundus of the stomach. Together these posterior relations make up what is sometimes referred to as the ‘bed’ of the liver (Fig. 68.1).
FUNCTIONAL ANATOMICAL DIVISIONS
Current understanding of the functional anatomy of the liver is based on Couinaud’s division of the liver into eight (subsequently nine) functional segments, based upon the distribution of portal venous branches and the location of the hepatic veins in the parenchyma (Couinaud 1957). Further understanding of the intrahepatic biliary anatomy, especially of the right ductal system, was enhanced by contributions from Hjortsjo (1948) and Healey & Schroy (1953) using the biliary system as the main guide for division of the liver (Fig. 68.4).
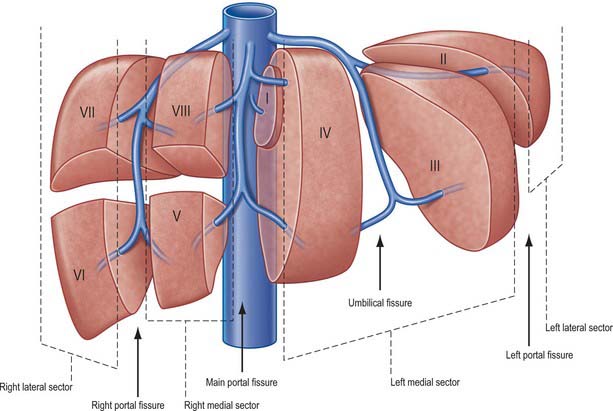
Sectors and segments of the liver
Sectors
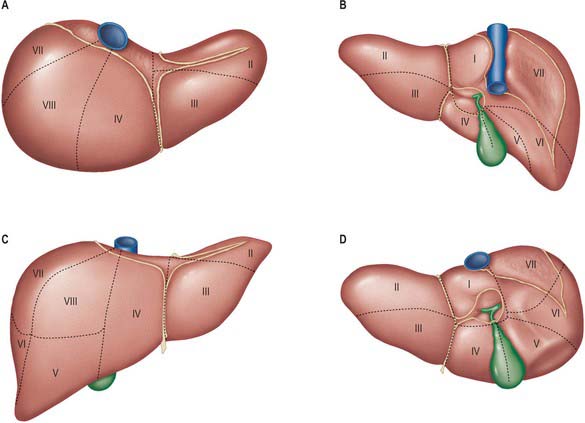
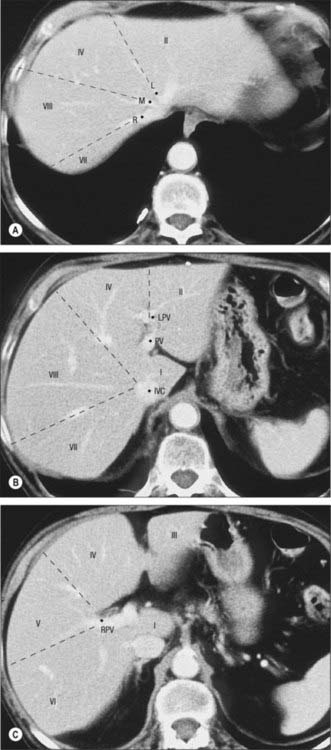
The sectors of the liver are made up of between one and three segments: right lateral sector = segments VI and VII; right medial sector = segments V and VIII; left medial sector = segments III and IV (and part of I); left lateral sector = segment II (Fig. 68.5). Segments are numbered in an ante-clockwise spiral centered on the portal vein with the liver viewed from beneath, starting with segment I up to segment VI, and then back clockwise for the most cranial two segments VII and VIII (Fig. 68.6).
Peritoneal attachments and ligaments of the liver
Falciform ligament
The liver is attached in front to the anterior abdominal wall by the falciform ligament. The two layers of this ligament descend from the posterior surface of the anterior abdominal wall and diaphragm and turn onto the anterior and superior surfaces of the liver. On the dome of the superior surface, the right leaf runs laterally and is continuous with the upper layer of the coronary ligament. The left layer of the falciform ligament turns medially and is continuous with the anterior layer of the left triangular ligament. The ligamentum teres, which represents the obliterated left umbilical vein, runs in the lower free border of the falciform ligament and continues into a fissure on the inferior surface of the liver. In fetal life the left umbilical vein opens into the left portal vein: it is supposed to be obliterated in adult life, but frequently remains partially patent. This lumen may open up in conditions such as portal hypertension to form a collateral channel. The ligamentum teres has importance in abdominal surgery for several reasons. It is quite often divided in upper abdominal surgery to optimize access to the upper abdominal viscera or as the first step in the mobilization of the liver. The ligament is vascularized by numerous arterial branches, mainly from the segment IV artery, and these form an anastomotic connection with the branches of the internal thoracic artery: it is therefore important to ligate or coagulate the ligament during its division. The ligamentum teres is a landmark and guide to the segment III hepatic duct used in hepatocojejunostomy formation, and to the left portal vein lying in the umbilical fissure during creation of a mesentericoportal shunt.
Porta hepatis, hepatoduodenal ligament and hilar plate
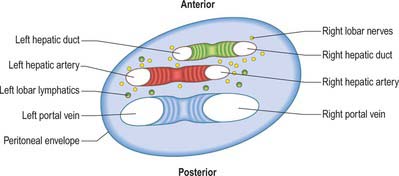
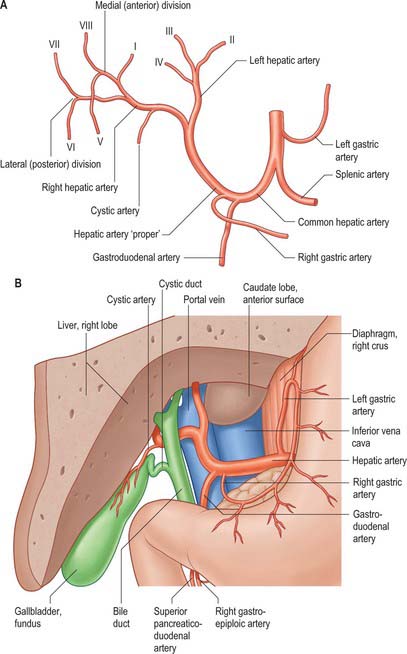
The porta hepatis is a deep fissure on the inferior surface of the liver. It is situated between the quadrate lobe in front and the caudate process behind, and contains the portal vein, hepatic artery and hepatic nervous plexuses as they ascend into the parenchyma of the liver, and the right and left bile ducts and some lymph vessels as they emerge from the liver. The hepatic ducts lie anterior to the portal vein and its branches, and the hepatic artery with its branches lies between the two (Fig. 68.7). All these structures are enveloped in the perivascular fibrous capsule, the hepatobiliary capsule of Glisson, a sheath of loose connective tissue which surrounds the vessels as they course through the portal canals in the liver and is also continuous with the fibrous hepatic capsule. The dense aggregation of vessels, supporting connective tissue, and liver parenchyma just above the porta hepatis is often referred to as the ‘hilar plate’ of the liver (Fig. 68.8). Understanding of the concept of hilar plate is important in surgical approaches to the hilar structures. Division or lowering of the hilar plate are essential for surgical access to the left hepatic duct. The hepatic artery, bile duct and portal vein extend from the porta hepatis towards the duodenum in the free edge of the hepatoduodenal ligament, which forms the anterior boundary of the epiploic foramen. Rapid control of the vessels entering the porta hepatis can be obtained by dividing the lesser omentum to the left of these structures and passing a tape around them from left to right through the epiploic foramen (a ‘Pringle’ manoeuvre).
VASCULAR SUPPLY AND LYMPHATIC DRAINAGE
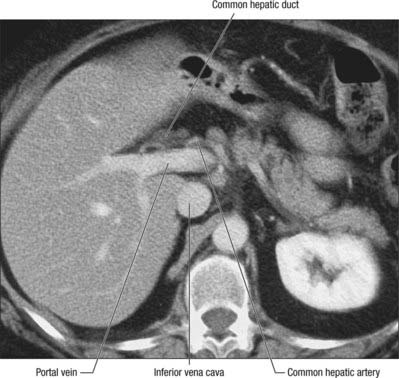
The vessels connected with the liver are the portal vein, hepatic artery and hepatic veins. The portal vein and hepatic artery ascend in the lesser omentum to the porta hepatis, where each bifurcates. The hepatic bile duct and lymphatic vessels descend from the porta hepatis in the same omentum (Figs 68.7 and 68.8). The hepatic veins leave the liver via its posterior surface and run directly into the inferior vena cava.
Hepatic artery
In adults the hepatic artery is intermediate in size between the left gastric and splenic arteries. In fetal and early postnatal life it is the largest branch of the coeliac axis. The hepatic artery gives off right gastric, gastroduodenal and cystic branches as well as direct branches to the bile duct from the right hepatic and sometimes the supraduodenal artery (Fig. 68.9A). After its origin from the coeliac axis, the hepatic artery passes anteriorly and laterally below the epiploic foramen to the upper aspect of the first part of the duodenum. It may be subdivided into the common hepatic artery, from the coeliac trunk to the origin of the gastroduodenal artery, and the hepatic artery ‘proper’, from that point to its bifurcation. It passes anterior to the portal vein and ascends anterior to the epiploic foramen between the layers of the lesser omentum. Within the free border of the lesser omentum the hepatic artery is medial to the common bile duct and anterior to the portal vein. At the porta hepatis it divides into right and left branches before these run into the parenchyma of the liver. The right hepatic artery usually crosses posterior (occasionally anterior) to the common hepatic duct (Fig. 68.9B). This close proximity often means that the right hepatic artery is involved in bile duct cancer earlier than the left hepatic artery. Occasionally the right hepatic artery crosses in front of the common bile duct and may be injured in surgery of the common bile duct. It almost always divides into an anterior branch supplying segments V and VIII, and a posterior branch supplying segments VI and VII. The anterior division often supplies a branch to segment I and the gallbladder. The segmental arteries are macroscopically end-arteries although some collateral circulation occurs between segments via fine terminal branches.
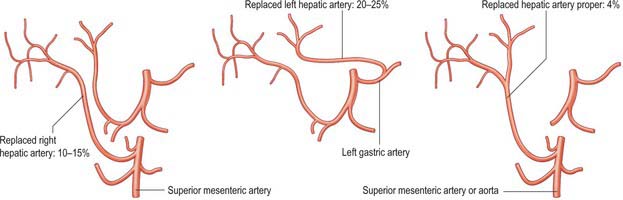
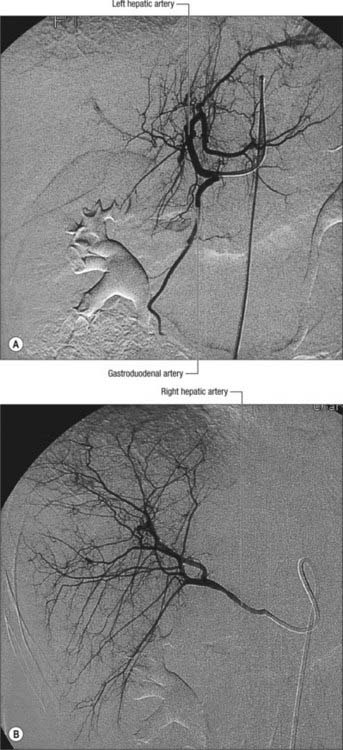
A small number of normal variants are important to demonstrate angiographically because they may influence surgical and interventional radiological procedures. A vessel that supplies a lobe in addition to its normal vessel is defined as an accessory artery. A replaced hepatic artery is a vessel that does not originate from an orthodox position and provides the sole supply to that lobe. Rarely a replaced common hepatic artery arises from the superior mesenteric artery (Fig. 68.10) and is identified at surgery by a relatively superficial portal vein (reflecting the absence of a common hepatic artery that would normally cross in front of the vein). More commonly a replaced right hepatic artery or an accessory right hepatic artery arises from the superior mesenteric artery (Fig. 68.11). In this case they run behind the portal vein and bile duct in the lesser omentum and can be identified at surgery by pulsation behind the portal vein. The accessory right hepatic artery may be injured during resections of the pancreatic head because the artery lies in close proximity to the portal vein. Occasionally, a replaced left hepatic artery or an accessory branch arises from the left gastric artery: these vessels provides a source of collateral arterial circulation in cases of occlusion of the vessels in the porta hepatis but may also be injured during mobilization of the stomach as it lies in the upper portion of the lesser omentum. Rarely, accessory left or right hepatic arteries may arise from the gastroduodenal artery or aorta. The presence of replaced arteries can be lifesaving in patients with bile duct cancer: because they are further away from the bile duct they tend to be spared from the cancer, making excision of the tumour feasible. Knowledge of these variations is also important in planning whole and split liver transplantation.
Veins
The liver has two venous systems. The portal system conveys venous blood from the majority of the gastrointestinal tract and its associated organs to the liver (see Ch. 60). The hepatic venous system drains blood from the liver parenchyma into the inferior vena cava.
Portal vein
The portal vein begins at the level of the second lumbar vertebra and is formed from the convergence of the superior mesenteric and splenic veins (Fig. 68.12; see also Fig. 71.4 and Fig. 60.8. It is approximately 8 cm long and lies anterior to the inferior vena cava and posterior to the neck of the pancreas. It lies obliquely to the right and ascends behind the first part of the duodenum, the common bile duct and gastroduodenal artery. At this point it is directly anterior to the inferior vena cava. It enters the right border of the lesser omentum, ascends anterior to the epiploic foramen to reach the right end of the porta hepatis and then divides into right and left main branches which accompany the corresponding branches of the hepatic artery into the liver. In the lesser omentum the portal vein lies posterior to both the common bile duct and hepatic artery. It is surrounded by the hepatic nerve plexus and accompanied by many lymph vessels and some lymph nodes.
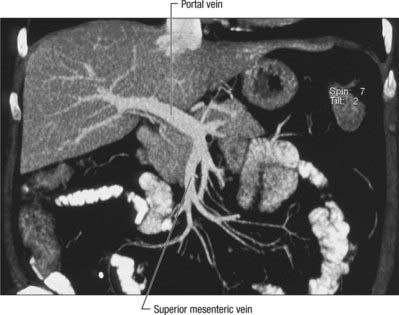
The main extrahepatic tributaries of the portal vein are the coronary or the left gastric vein, which ends in the left margin of the portal vein, and the posterior superior pancreatoduodenal vein nearer to the head of the pancreas. The portal vein divides into right and left branches at the hilum (Fig. 68.13). The left portal vein has a longer extraparenchymal course (4–5 cm) and tends to lie slightly more horizontally than the right portal vein, but is often of smaller calibre. It has horizontal and vertical portions. The horizontal portion runs along the base of segment VI and often gives branches to segment I and sometimes to segment VI in this part of its course. The branch to segment II continues laterally but the main left portal vein takes a more anterior and vertical course in the umbilical fissure (the vein of the umbilical fissure) where it gives branches to segments III and IV and receives the obliterated left umbilical vein (ligamentum teres). The majority of the supply to segment IV comes from the left portal vein, and only occasionally from the right via proximal branches of the main vein or branches from veins to segments V or VIII. The right portal vein is only 2–3 cm in length and usually divides into a right medial (anterior) sectoral division supplying segments V and VIII, and a right lateral (posterior) sectoral division supplying segments VI and VII. The medial division may give a branch to segment I.
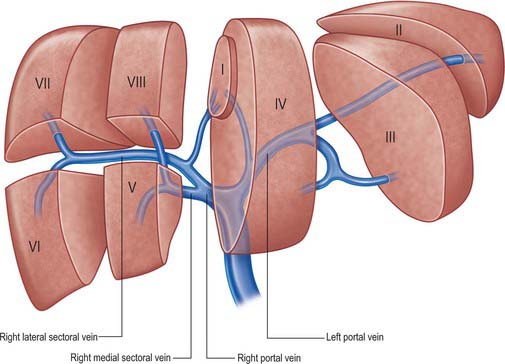
Variations usually involve the right portal vein: absence of a right portal vein with the resulting portal trifurcation in the form of left portal vein, right medial and right lateral portal veins, is present in 10–15% of livers. Occasionally the right medial vein arises from the left portal vein, a variant which is important to remember during left sided liver resection. The portal trifurcation has implications for split liver and live donor liver transplantation, where its presence might be considered as a relative contra-indication. On rare occasions, the portal bifurcation is absent, in which case the main portal vein enters the liver giving off the right segmental branches and then turns left to supply the left lobe of the liver (a contraindication to major liver surgery). Occasionally one or more of the segmental branches of the right lobe (especially segment IV) arises proximally.
Porto-systemic shunts
Increased pressure within the portal venous system may result in dilatation of the portal venous tributaries: a reversal of flow may occur where these veins form anastomoses with veins which drain into the systemic venous circulation. The common sites where porto-systemic shunts may occur, and the associated clinical implications, are listed in Table 68.1.
Table 68.1 Common sites of occurrence of porto-systemic shunts, and associated clinical implications.
| Portal veins | Systemic veins | Clinical presentations |
|---|---|---|
| Left gastric and lower oesophageal veins | Lower branches of oesophageal veins that drain into azygos and accessory hemiazygos veins | Oesophageal or gastric varices |
| Superior rectal veins | Middle and inferior rectal veins that drain into internal iliac and pudendal veins | Rectal varices |
| Persistent tributaries of left branch of portal vein, running in ligamentum teres | Peri-umbilical branches of superior and inferior epigastric veins | ‘Caput medusae’ |
| Intraparenchymal branches of right branch of portal vein, lying in liver tissue exposed in ‘bare area’ | Retroperitoneal veins that drain into lumbar, azygos and hemiazygos veins | Retroperitoneal dilated veins are at risk during surgery or interventional procedures |
| Omental and colonic veins in the region of the hepatic and splenic flexure | Retroperitoneal veins in the region of the hepatic and splenic flexure | |
| Patent ductus venosus connected to the left branch of the portal vein | Inferior vena cava | Is rare |
Hepatic veins
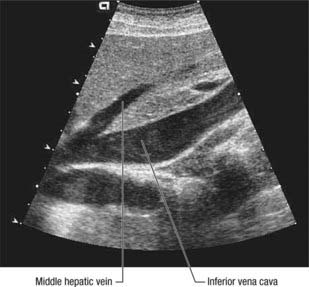
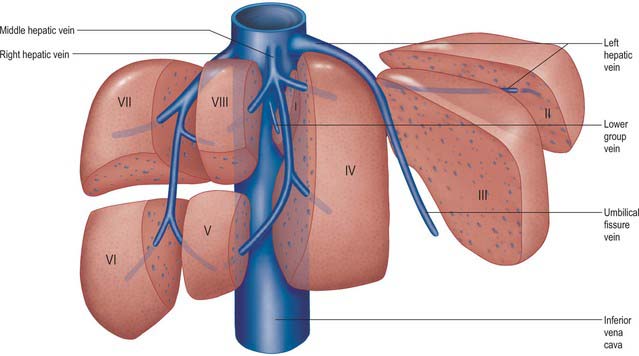
The liver is drained by three major hepatic veins into the suprahepatic part of the inferior vena cava and a multitude of minor hepatic veins that drain into the intrahepatic inferior vena cava. The three major veins are located between the four major sectors of the liver (Figs 68.6A, 68.14 and 68.15).
INNERVATION
Hepatic plexus
Nerves from the hepatic plexus run with the branches of the common hepatic artery to supply, or contribute to the supply of, foregut derivatives. Branches run inferiorly from the plexus with the right gastric artery to contribute to the supply of the pylorus, with the gastroduodenal artery and its branches to reach the pylorus and the first part of the duodenum, and with the right gastroepiploic artery to provide a small contribution to the supply the right side of the stomach and the greater curvature. The superior pancreaticoduodenal extension supplies the descending part of the duodenum, the pancreatic head and the intrapancreatic part of the common bile duct.
MICROSTRUCTURE
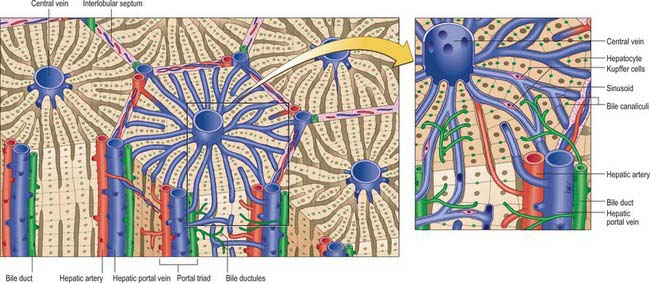
The liver is essentially an epithelial-mesenchymal outgrowth of the caudal part of the foregut, with which it retains its connection via the biliary tree. The surface of the liver facing the peritoneal cavity is covered by a typical serosa, the visceral peritoneum. Beneath this, and enclosing the whole structure, is a thin (50–100 μm) layer of connective tissue from which extensions pass into the liver as connective tissue septa and trabeculae. Branches of the hepatic artery and hepatic portal vein, together with bile ductules and ducts, run within these connective tissue trabeculae which are termed portal tracts (portal canals). The combination of the two types of vessel and a bile duct is termed a portal triad (Fig. 68.16); these structures are usually accompanied by one or more lymphatic vessels.
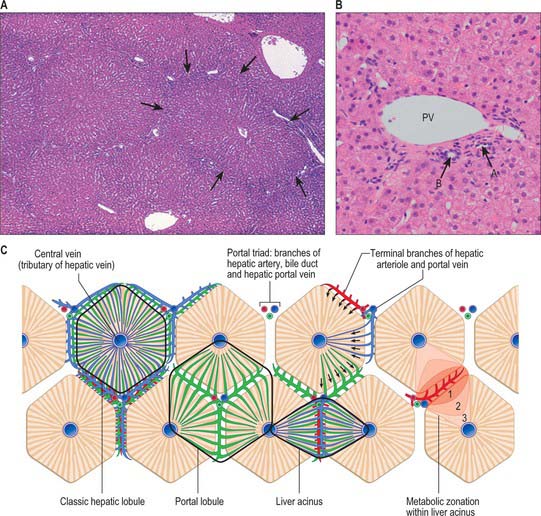
Lobulation of the liver
The structural unit of the liver is the lobule: a roughly hexagonal arrangement of plates of hepatocytes, separated by intervening sinusoids which radiate outward from a central vein, with portal triads at the vertices of each hexagon (Fig. 68.16). The central vein is a tributary of the hepatic vein that drains the tissue. In some species, the classic lobular units are delimited microscopically by distinct connective tissue septa. However, the lobular organization of the human liver is not immediately evident in histological sections: the lobules do not have distinct boundaries, and connective tissue is sparse. The plates do not pass straight to the periphery of a lobule like the spokes of a wheel but run irregularly as they anastomose and branch.
Cells of the liver
Cells of the liver include hepatocytes, hepatic stellate cells (also known as perisinusoidal lipocytes, or Ito cells), sinusoidal endothelial cells, macrophages (Kupffer cells), the cells of the biliary tree (cuboidal to columnar epithelium) and connective tissue cells of the capsule and portal tracts (Figs 68.16B and 68.17).
About 80% of the liver volume and 60% of its cell number are formed by hepatocytes (parenchymal cells) (Fig. 68.17). They are polyhedral, with 5–12 sides and are from 20 to 30 μm across. Their nuclei are round, euchromatic and often tetraploid, polyploid or multiple, with two or more in each cell. Their cytoplasm typically contains a considerable amount of rough and smooth endoplasmic reticulum, many mitochondria, lysosomes and well-developed Golgi apparatus, which are all features that indicate a high metabolic activity. Glycogen granules and lipid vacuoles are usually prominent. Numerous large peroxisomes and vacuoles containing enzymes, e.g. urease in distinctive crystalline forms, indicate the complex metabolism of these cells. Their role in iron metabolism is reflected by the presence of storage vacuoles containing crystals of ferritin and haemosiderin.
Couinaud C. Le foié: etudes anatomique et chirurgicules. Paris: Masson, 1957.
The original description of hepatic segmentation by Couinaud..
Healey JE, Schroy PC. Anatomy of the biliary ducts within the human liver; analysis of the prevailing pattern of branchings and the major variations of the biliary ducts. Arch Surg. 1953;66:599-616.
Mitchell AWM, Dick R. Liver, gall-bladder, pancreas and spleen. In: Butler P, Mitchell AWM, Ellis H. Applied Radiological Anatomy. Cambridge: Cambridge University Press; 1999:239-258.
[/level-membership-for-basic-science-category][not-level-membership-for-basic-science-category]
CHAPTER 68 Liver
The liver is the largest of the abdominal viscera, occupying a substantial portion of the upper abdominal cavity. It occupies most of the right hypochondrium and epigastrium, and frequently extends into the left hypochondrium as far as the left lateral line (Fig. 68.1). As the body grows from infancy to adulthood the liver rapidly increases in size. This period of growth reaches a plateau around 18 years and is followed by a gradual decrease in the liver weight from middle age. The ratio of liver to body weight decreases with growth from infancy to adulthood. The liver weighs approximately 5% of the body weight in infancy and it decreases to approximately 2% in adulthood. The size of the liver also varies according to sex, age and body size. It has an overall wedge shape, which is in part determined by the form of the upper abdominal cavity into which it grows. The narrow end of the wedge lies towards the left hypochondrium, and the anterior edge points anteriorly and inferiorly. The superior and right lateral aspects are shaped by the anterolateral abdominal and chest wall as well as the diaphragm. The inferior aspect is shaped by the adjacent viscera. The capsule is no longer thought to play an important part in maintaining the integrity of the shape of the liver.
Fig. 68.1 The ‘bed’ of the liver. The outline of the liver is shaded green. The central bare area is unshaded.
SURFACES OF THE LIVER
The liver is usually described as having superior, anterior, right, posterior and inferior surfaces, and has a distinct inferior border (Figs 68.2, 68.3). However, the superior, anterior and right surfaces are continuous and no definable borders separate them. It would be more appropriate to group them as the diaphragmatic surface, which is mostly separated from the inferior, or visceral surface, by a narrow inferior border. At the infrasternal angle the inferior border is related to the anterior abdominal wall and is accessible to examination by percussion, but is not usually palpable. In the midline, the inferior border of the liver is near the transpyloric plane, about a hand’s breadth below the xiphisternal joint. In women and children the border often projects a little below the right costal margin.
Fig. 68.3 Relations of the liver. A, superior view; B, posterior view; C, anterior view; D, inferior view.
The posterior surface is convex, wide on the right, but narrow on the left. A deep median concavity corresponds to the forward convexity of the vertebral column close to the attachment of the ligamentum venosum. Much of the posterior surface is attached to the diaphragm by loose connective tissue, forming the triangular ‘bare area’. The inferior vena cava lies in a groove or tunnel in the medial end of the ‘bare area’. To the left of the caval groove the posterior surface of the liver is formed by the caudate lobe, and covered by a layer of peritoneum continuous with that of the inferior layer of the coronary ligament and the layers of the lesser omentum. The caudate lobe is related to the diaphragmatic crura above the aortic opening and the right inferior phrenic artery, and separated by these structures from the descending thoracic aorta.
The fissure for the ligamentum venosum separates the posterior aspect of the caudate lobe from the main part of the left lobe. The fissure cuts deeply in front of the caudate lobe and contains the two layers of the lesser omentum. The posterior surface over the left lobe bears a shallow impression near the upper end of the fissure for the ligamentum venosum that is caused by the abdominal part of the oesophagus. The posterior surface of the left lobe to the left of this impression is related to part of the fundus of the stomach. Together these posterior relations make up what is sometimes referred to as the ‘bed’ of the liver (Fig. 68.1).
FUNCTIONAL ANATOMICAL DIVISIONS
Current understanding of the functional anatomy of the liver is based on Couinaud’s division of the liver into eight (subsequently nine) functional segments, based upon the distribution of portal venous branches and the location of the hepatic veins in the parenchyma (Couinaud 1957). Further understanding of the intrahepatic biliary anatomy, especially of the right ductal system, was enhanced by contributions from Hjortsjo (1948) and Healey & Schroy (1953) using the biliary system as the main guide for division of the liver (Fig. 68.4).
Sectors and segments of the liver
Sectors
The sectors of the liver are made up of between one and three segments: right lateral sector = segments VI and VII; right medial sector = segments V and VIII; left medial sector = segments III and IV (and part of I); left lateral sector = segment II (Fig. 68.5). Segments are numbered in an ante-clockwise spiral centered on the portal vein with the liver viewed from beneath, starting with segment I up to segment VI, and then back clockwise for the most cranial two segments VII and VIII (Fig. 68.6).
