11 Innervation of skin
Sensory Units
A given stem fiber forms the same kind of nerve ending at all of its terminals. In physiologic recordings, the stem fiber and its family of endings constitute a sensory unit. Together with its parent unipolar nerve cell, the sensory unit is analogous to the motor unit described in Chapter 10.
Nerve Endings
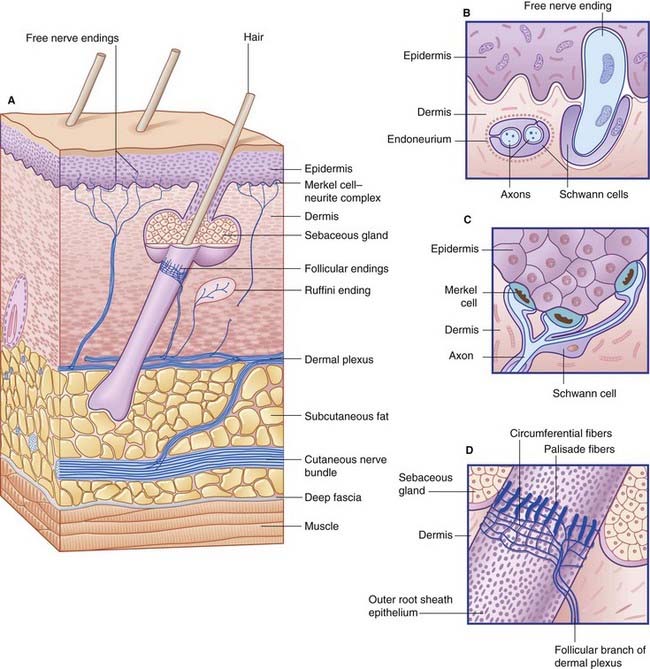
Free nerve endings (Figure 11.1A, B)
Functions
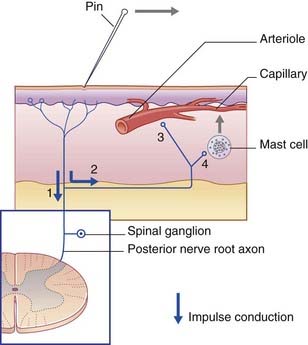
C-fiber units are responsible for the axon reflex (Clinical Panel 11.1).
Clinical Panel 11.1 Neurogenic inflammation: the axon reflex
When sensitive skin is stroked with a sharp object, a red line appears in seconds, owing to capillary dilatation in direct response to the injury. A few minutes later, a red flare spreads into the surrounding skin, owing to arteriolar dilatation, followed by a white wheal, owing to exudation of plasma from the capillaries. These phenomena constitute the triple response. The wheal and flare responses are produced by axon reflexes in the local sensory cutaneous nerves. The sequence of events follows the numbers in Figure CP 11.1.1.
Encapsulated nerve endings
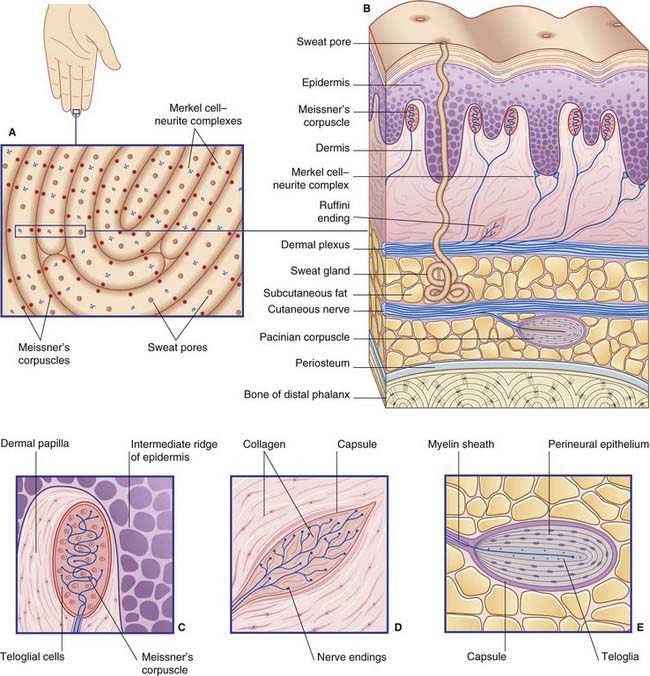
The digital receptors are classified as follows by sensory physiologists.
Cutaneous sensory effects of peripheral neuropathies are described in Chapter 12. Clinical Panel 11.2 gives a short account of leprosy.
Lauria G, Lombardi R. Skin biopsy: a new tool for diagnosing peripheral neuropathy. BMJ. 2007:1159-1162.
Meftah E-M, Chapman CE. Tactile perception of roughness. Exp Brain Res. 2009;3:235-244.
Schepers RJ, Ringkamp M. Thermoreceptors and thermoceciptive afferents. Neurosci Behav Rev. 2009;33:205-212.