CHAPTER 81 Hip
The hip joint is a synovial joint of ball-and-socket (multiaxial spheroidal, cotyloid) type (Figs 81.1–81.3).
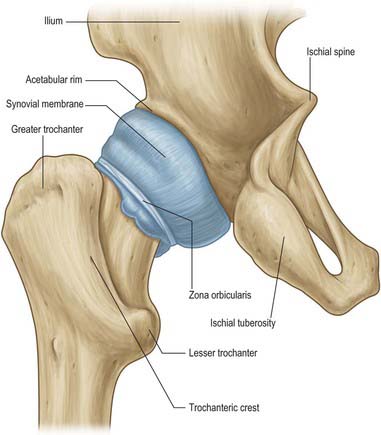
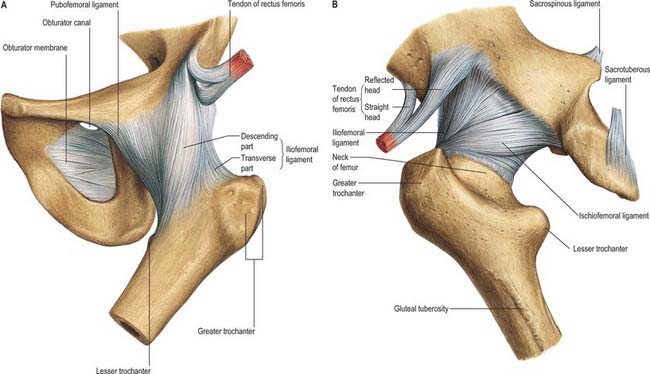
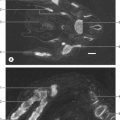
Fig. 81.3 The ligaments of the hip joint: A, anterior aspect; B, posterior aspect.
(From Sobotta 2006.)
ARTICULAR SURFACES
The femoral head articulates with the cup-shaped (cotyloid) acetabulum, its centre lying a little below the middle third of the inguinal ligament. (The profile of the anterior margin of the joint is parallel to the middle third of the inguinal ligament.) The articular surfaces are reciprocally curved but neither coextensive nor completely congruent (Fig. 81.2; see Figs 80.4, 80.15). The close-packed position of the hip joint is one of full extension, with slight abduction and medial rotation. As in the shoulder joint, the surfaces are considered ovoid or spheroidal rather than spherical, but this view is controversial. Evidence appears to suggest that the articular surface on the femoral head is a spheroidal, or slightly ovoid, surface in youth but that it tends to become almost spherical with advancing age. The femoral head is covered by articular cartilage, except over the rough pit where the ligamentum teres is attached. In front the cartilage extends laterally over a small area on the adjoining neck. Articular cartilage is, generally, thicker centrally than at the periphery. Cartilage thickness is maximal anterosuperiorly in the acetabulum and anterolaterally on the femoral head, the two areas that correspond to the principal load-bearing areas within the joint The acetabular articular surface, the lunate surface, is an incomplete ring, broadest anterosuperiorly where the pressure of body weight falls in the erect posture, and narrowest in its pubic region. It is deficient inferiorly opposite the acetabular notch. The lunate surface is covered by articular cartilage, which is thickest where the surface is broadest. The acetabular fossa, the central non-articular area in the floor of the acetabulum, is devoid of cartilage but contains fibroelastic fat largely covered by synovial membrane. The acetabular labrum, a fibrocartilaginous rim attached to the acetabular margin, serves to deepen the acetabulum and bridges the acetabular notch by attaching to the peripheral edge of the transverse acetabular ligament. The labrum is triangular in section; it is attached by its base to the acetabular margin and its acute free edge projects beyond the acetabular margin. The diameter of the acetabular cavity is constricted by the labral rim which embraces the femoral head, maintaining joint stability both as a static restraint and by providing proprioceptive information.
FIBROUS CAPSULE
The capsule is strong and dense. It is attached above to the acetabular margin 5–6 mm medial to the labral attachment, in front to the outer labral aspect and, near the acetabular notch, to the transverse acetabular ligament and the adjacent rim of the obturator foramen. From its acetabular attachment it extends laterally to surround the femoral head and neck, and is attached anteriorly to the intertrochanteric line, superiorly to the base of the femoral neck, posteriorly 1 cm superomedial to the intertrochanteric crest, and inferiorly to the femoral neck near the lesser trochanter. Anteriorly many fibres ascend along the neck as longitudinal retinacula, containing blood vessels for both the femoral head and neck. The capsule is thicker anterosuperiorly, where maximal stress occurs, particularly in standing. Posteroinferiorly it is relatively thin and loosely attached. It has two sets of fibres, circular and longitudinal. The circular fibres (zona orbicularis) are internal and form a collar round the femoral neck; although partly blended with the pubofemoral and ischiofemoral ligaments, these fibres are not directly attached to bone. Externally, longitudinal fibres are most numerous in the anterosuperior region, where they are reinforced by the iliofemoral ligament. The capsule is also strengthened inferiorly by the pubofemoral ligament, and posteriorly by the ischiofemoral ligament. Externally it is rough, covered by muscles and tendons and separated anteriorly from psoas major and iliacus by a bursa (see below). The capsular attachment to the femur lies well distal to the growth plate of the femoral head both anteriorly and posteriorly. Thus the upper femoral epiphysis is entirely intracapsular. The capsular attachment intersects the growth plate of the greater trochanter on the upper surface of the base of the neck (see Fig. 80.18).
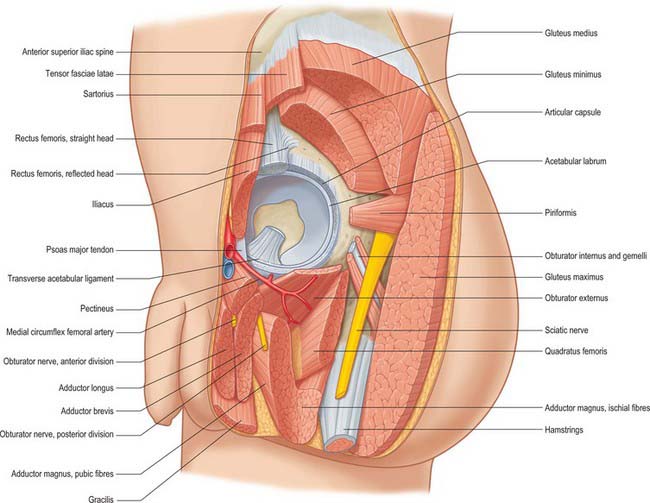
The joint capsule is surrounded by muscles (Fig. 81.4). Anteriorly, lateral fibres of pectineus separate it from the femoral vein. Lateral to this the tendon of psoas major, with iliacus lateral to it, descends across it, partly separated from the capsule by a bursa. The femoral artery is anterior to the tendon of psoas major, and the femoral nerve lies deep in a groove between the tendon and iliacus. More laterally the straight head of rectus femoris crosses the joint with a deep layer of the fascial iliotibial tract, which blends with the capsule under the lateral border of the muscle. Superiorly, the reflected head of rectus femoris contacts the capsule medially, while gluteus minimus covers it laterally, being closely adherent. Inferiorly, medial fibres of pectineus adjoin the capsule and, more posteriorly, obturator externus spirals obliquely to its posterior aspect. Posteroinferiorly, the capsule is covered by the tendon of obturator externus, separating it from quadratus femoris and accompanied by an ascending branch of the medial circumflex femoral artery. Above this the tendon of obturator internus and the gemelli contact the joint capsule, separating it from the sciatic nerve. The nerve to quadratus femoris is deep to the obturator internus tendon, and descends medially on the capsule. Above this, the posterior surface of the joint is crossed by piriformis.
LIGAMENTS
The ligaments of the hip joint (Fig. 81.3) are the iliofemoral, pubofemoral, ischiofemoral and transverse acetabular ligaments and the ligamentum teres. As the hip moves so the capsular ligaments, as capsular thickenings, wind and unwind, tightening around the hip, affecting stability, excursion and joint capacity. Joint capacity is maximal when the hip is held in a partially flexed and abducted position: a patient with an effusion in the hip joint is therefore most comfortable when the joint is held in a position of flexion and abduction. See Fuss & Bacher (1991) for details.
The ischiofemoral ligament thickens the back of the capsule and consists of three distinct parts. The central part, the superior ischiofemoral ligament, spirals superolaterally from the ischium, where it is attached posteroinferior to the acetabulum, behind the femoral neck to attach to the greater trochanter deep to the iliofemoral ligament. Some fibres blend with the zona orbicularis. Lateral and medial inferior ischiofemoral ligaments embrace the posterior circumference of the femoral neck.
Transverse acetabular ligament
The transverse acetabular ligament (see Fig. 80.4) is continuous peripherally with the labrum but does not, itself, possess any cartilage cells. Its strong, flat fibres cross the acetabular notch forming a foramen through which vessels and nerves enter the joint.
Ligamentum teres (ligament of the head of the femur)
The ligamentum teres (Fig. 81.5) is a triangular and somewhat flattened band. Its apex is attached anterosuperiorly in the fovea on the femoral head and its base is attached principally to both edges of the acetabular notch, between which it blends with the transverse ligament. It also receives weaker contributions from the margins of the acetabular fossa. The ligamentum teres is ensheathed by synovial membrane, and varies in strength. Occasionally the synovial sheath exists by itself in the absence of the ligament; rarely both ligament and sheath are absent. The ligament appears to become tense when the hip joint is semi-flexed and adducted, and appears to relax when the joint is abducted.
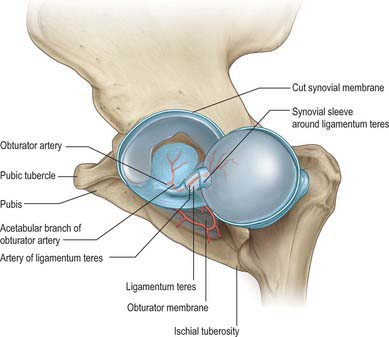
VASCULAR SUPPLY AND LYMPHATIC DRAINAGE
Articular arteries are branches from the obturator, medial circumflex femoral, and superior and inferior gluteal arteries, with corresponding venous drainage. They form the cruciate and trochanteric anastomoses (Ch. 80). Lymphatics from the anterior aspect of the hip joint drain to the deep inguinal nodes, while those from the medial and posterior aspects run with the obturator and gluteal arteries respectively to reach the internal iliac nodes.
INNERVATION
Pain in the hip and groin may be due to any of a number of possible causes. True hip joint pain is usually felt deep in the crease of the groin. Pain over the lateral aspect of the hip usually suggests a local problem such as trochanteric bursitis. Pain in the buttock is only occasionally from the hip joint: most frequently it is referred from the lumbosacral spine or may be caused by vascular claudication pain. Pain in the groin may be due to inguinal or femoral hernias (see Ch. 61). In the younger age group groin pain afflicts those engaged in sport, especially running ball sports. Here the pain is sometimes due to a hernia but more commonly related to adductor tendinitis secondary to acute or chronic overload of the adductor longus tendon. Another common problem is attritional chronic overload of the anterior abdominal wall at its attachments to the pubis and inguinal ligament, or of external oblique at the external inguinal ring. Rarely problems may occur at the symphysis pubis.
BIOMECHANICS OF THE HIP JOINT
FEMUR
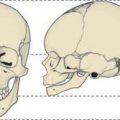
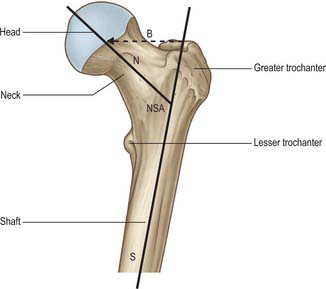
The femur is essentially a tubular structure with distortions that consist of bows and twists. The most notable is the anterior bow in its mid-portion, where the radius of curvature is relatively constant along the length of the femoral shaft. In the coronal plane, the femoral neck is inclined obliquely to the shaft at an angle of 135° (range 120–145°; Fig. 81.6). Although the neck/shaft angle and neck length are variable, the centre of the neck in the coronal plane is at the level of the apex of the greater trochanter. In the axial plane the femoral neck is anteverted, i.e. rotated anteriorly relative to the posterior surfaces of the femoral condyles: in the adult, this angle is 10–15o (Fig. 81.7). The degree of anteversion affects many aspects of lower limb biomechanics, including the abductor moment arm (the perpendicular distance from the centre of rotation of the femur to the line of action of the resultant abductor muscle force); patellar tracking (the motion of the patella relative to the femur or femoral groove on knee flexion and extension); foot orientation.
MOVEMENTS
With the foot stationary on the ground, medial and lateral femoral rotation occur around a vertical axis through the centre of the femoral head and medial condyle. This axial medial rotation is the inevitable conjunct rotation that accompanies terminal extension of the knee and is often referred to as ‘screw-home’. The reverse occurs in initial flexion at the knee joint. The ‘screw-home’ phenomenon allows achievement of a ‘close packed position’ for the tibiofemoral joint, so aiding stability in the extended knee. The relationship of this axis to the whole femur (which has an angulated neck and oblique shaft) means that the medial condyle moves very little on the medial tibial condyle during medial rotation, while the lateral condyle moves forward on its tibial surface, and the greater trochanter moves simultaneously in a forward arc: converse movements occur during lateral rotation. With the foot either free or in loose contact with the ground, medial and lateral adjunct rotation of the whole lower limb occur around variable axes, all of which pass through the femoral head and any part of the foot. Conversely, with one foot stationary and the other free, the whole trunk may rotate on one femoral head, as in cross-kicking. Abduction and adduction occur about an anteroposterior axis through the femoral head, but since the latter is not strictly spherical, the axis is not a fixed one. Some kinesiologists refer to a mechanical axis coincident with the topographical long axis of the femoral neck, impinging on the approximate centre of the articular surface of the head, making extension and flexion of the thigh relatively pure ‘spins’ at the hip joint and most effective in tightening or relaxing the capsular ligaments. All other movements are regarded as pure or impure swings. While this permits analysis closer to actual articular function than a cardinal triaxial system related arbitrarily to the ‘anatomical’ position, the mechanical axis is itself regarded (incorrectly) by some as dynamic because forms of apparent dynamic ‘spin’ may occur in many positions. This view stems from an imprecise definition of the varieties of axes employed in arthrokinematics. The mechanical axis is not dynamic relative to the femur. It is stationary during pure spins, and moves relative to its coarticular surface in chordal or arcuate paths during pure or impure swings respectively.
Simple flexion is possible to 110–120° from the vertical; extension beyond the vertical is limited (perhaps 10–20°). Both movements are augmented by adjustments of the spinal column and pelvis, flexion of the knee and concomitant medial or lateral hip rotation. For example, knee flexion (lessening tension in the posterior femoral muscles) increases hip flexion to 120°; the thigh can be drawn passively to the trunk, though with some spinal flexion. Extension in walking and running is increased by forward inclination of the body, pelvic tilting and rotation and lateral rotation of the hip (Joseph 1975). Abduction and adduction can be similarly increased.
FORCES ACTING ON THE HIP
The range of motion of the hip far exceeds that required for normal walking. The muscles of the joint are the major stabilizing factors: the ligaments of the hip joint provide little rotational stability when walking. During normal gait the hip is subjected to wide variations in load bearing, ranging from one third of body weight in the double-leg support phase to four times body weight in the single-leg support phase. The walking cycle is characterized by two load peaks (see p. 1460) at heel strike (see Fig. 81.9) and toe off, corresponding to maximal acceleration and deceleration respectively in single-leg stance (see Fig. 84.25). Between these two peaks the body translates smoothly with little vertical acceleration.
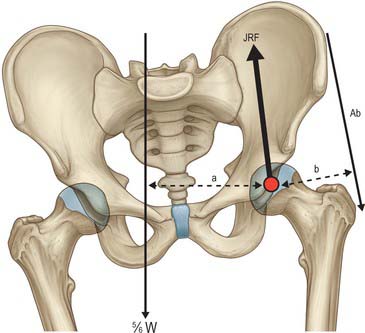
In single-leg stance, the effective centre of gravity moves away from the supporting leg as the unsupported leg contributes to body weight. Body weight will exert a turning moment around the centre of the supporting femoral head: in order to maintain equilibrium, an equal but opposite turning moment is provided by the abductor muscles (primarily gluteii medius and minimus, tensor fasciae latae, piriformis and obturator internus). Since the lever arm of the abductors is much shorter than that of body weight acting through the centre of gravity, an abduction force greater than body weight is required to maintain equilibrium (Fig. 81.8). The vectors of these two forces produce a resultant compressive load on the femur that is oriented approximately 16o obliquely, laterally and distally. The joint reaction vector is exactly equal and opposite to this vector and is approximately four times body weight.
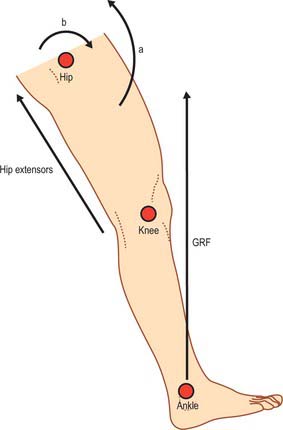
Simplified kinetics of heel strike
Contact with the ground produces a reaction force equal to body weight multiplied by acceleration (see p. 1443). This ground reaction force acting at the foot produces a flexion moment about the hip which is approximately 3.3 times body weight, and is counterbalanced by the hip extensors (gluteus maximus and hamstrings) (Fig. 81.9).