CHAPTER 1 Gastrointestinal Hormones and Neurotransmitters
CELLULAR COMMUNICATION
Chemical transmitters of the gut are produced by discrete cells of the GI mucosa and can be classified as endocrine, paracrine, synaptic (“neurocrine”), or autocrine (Fig. 1-1). Specialized signaling cells that secrete transmitters into the blood are known as endocrine cells, and the transmitters they produce are known as hormones. Hormones bind to specific receptors on the surface of target cells at remote sites and regulate metabolic processes.1
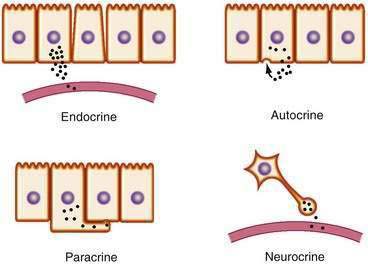
In contrast with endocrine cells that act on distant target tissues, other signaling cells of the GI tract may produce transmitters that act on neighboring cells. This process is known as paracrine signaling and is typical of cells that produce somatostatin.2 Paracrine transmitters are secreted locally and cannot diffuse far. They bind to receptors on nearby cells to exert their biological actions. These actions are limited because they are taken up rapidly by their target cells, destroyed by extracellular enzymes, and adhere to extracellular matrix, all of which limit their ability to act at distant sites. Because paracrine signals act locally, their onset of action is generally rapid and can be terminated abruptly. By comparison, endocrine signaling takes much longer, and termination of signaling requires clearance of hormone from the circulation.
Endocrine transmitters of the GI tract consist predominantly of peptides (e.g., gastrin, secretin). Paracrine transmitters can be peptides, such as somatostatin, or nonpeptides, such as histamine, that act locally on neighboring cells. Neurotransmitters can be peptides, such as vasoactive intestinal polypeptide (VIP) and tachykinins, or small molecules, such as acetylcholine and norepinephrine, that are secreted, or nitric oxide (NO), which simply diffuses across the synaptic cleft. The major transmitters and hormones of the GI tract are listed in Table 1-1.
Table 1-1 Hormones and Transmitters of the Gastrointestinal Tract
Peptides That May Function as Hormones, Neuropeptides, or Paracrine Agents
Some cells release messengers locally and possess cell surface receptors for the same messengers, thus enabling those cells to respond to their own secreted products. This mode of transmission, known as autocrine signaling, has been demonstrated for several growth factors and has been implicated in the growth of certain cancers, including colorectal cancer (see Chapter 3).3
NEURAL REGULATION OF THE GI TRACT
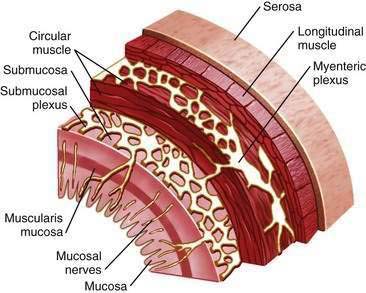
The enteric nervous system plays an integral role in the regulation of gut mucosal and motor function.4 It is organized into two major plexuses (Fig. 1-2). The myenteric plexus lies between the external longitudinal and internal circular muscle layers. The submucosal plexus lies between the circular muscle layer and the mucosa. Although the enteric nervous system receives input from the central and autonomic nervous systems, it can function independently. Nerves of the myenteric plexus project fibers primarily to the smooth muscle of the gut, with only a few axons extending to the submucosal plexus. Most of the fibers of the submucosal plexus project into the mucosa and the submucosal and myenteric plexuses. Various peptide and nonpeptide neurotransmitters are found in the enteric nervous system. Studies using immunohistochemical staining have localized neurotransmitters to specific neurons in the GI tract. γ-Aminobutyric acid is found primarily in the myenteric plexus and is involved in regulating smooth muscle contraction. Serotonin is found within the plexus and functions as an interneuron transmitter. Adrenergic neurons originate in ganglia of the autonomic nervous system and synapse with enteric neurons. Peptides such as neuropeptide Y (NPY) are often secreted from the same adrenergic neurons and generally exert inhibitory effects, such as vasoconstriction.5 Other adrenergic neurons containing somatostatin project to the submucosal plexus, where they inhibit intestinal secretion. Coexistence of peptides and neurotransmitters in the same neurons is not unusual; in fact, the interplay among transmitters is critical for coordinated neural regulation.6 For example, the peptides VIP and peptide histidine isoleucine (PHI) are commonly found together, as are the tachykinins substance P and substance K, where they have complementary effects.
The ability of hormones to act on nerves locally within the submucosa of the intestine and affect more distant sites on nerves such as the vagus expands the potential organs that may be regulated by gut hormones.7 Chemical and mechanical stimuli cause the release of hormones from endocrine cells of the intestinal mucosa. These interactions initiate a wide variety of secretomotor responses, many of which are mediated by enteric neurons. Secretomotor circuits consist of intrinsic primary afferent neurons with nerve endings in the mucosa and extension through the myenteric and submucosal plexi. This circuitry allows nerves to stimulate mucosal cells to secrete fluid and electrolytes and at the same time stimulate muscle contraction. The same motor neurons also have axons that supply arterioles and can initiate vasodilator reflexes.
Extrinsic primary afferent neurons can be of the vagus, with somal bodies in the nodose ganglia and axons that reach the gut through the vagus nerve, or of the spinal nerves of the thoracic and lumbar regions, whose cell bodies lie in the dorsal root ganglia. Information conducted by extrinsic primary afferent neurons includes pain, heat, and sensations of fullness or emptiness. These neurons are also targets for hormones. For example, the satiety effect of CCK in the bloodstream is mediated through the vagus nerve.8 Specific CCK receptors have been identified on the vagus, and blockade of these receptors abolishes the satiation induced by peripheral CCK.
Endocrine, paracrine, and neural transmitters existing within the lamina propria modulate effects on the gut immune system.7 Lymphocytes, macrophages, mast cells, neutrophils, and eosinophils are potential targets for endocrine and neural transmitters and participate in the inflammatory cascade. Moreover, inflammatory mediators can act directly on enteric nerves. Serotonin released from endocrine cells is involved in intestinal anaphylaxis and stimulates vagal afferent fibers that possess the 5-hydroxytryptamine 3 (5-HT3) receptor.
PEPTIDE HORMONES OF THE GI TRACT
SYNTHESIS, POST-TRANSLATIONAL MODIFICATION, AND SECRETION
All GI peptides are synthesized via gene transcription of DNA into messenger RNA (mRNA) and subsequent translation of mRNA into precursor proteins known as preprohormones. Peptides that are to be secreted contain a signal sequence that directs the newly translated protein to the endoplasmic reticulum, where the signal sequence is cleaved and the prepropeptide product is prepared for structural modifications.9 These precursors undergo intracellular processing and are transported to the Golgi apparatus and packaged in secretory granules. Further modifications in peptide structure may occur within the Golgi apparatus (e.g., sulfation) that is important for the bioactivity of many peptide hormones, such as CCK. Secretory granules may be targeted for immediate release or stored in close proximity to the plasma membrane for release following appropriate cell stimulation. When GI endocrine cells are stimulated, mature hormone is secreted into the paracellular space and is taken up into the bloodstream. For many hormones, such as gastrin and CCK, multiple molecular forms exist in blood and tissues. Although there is only a single gene for these peptides, the different molecular forms result from differences in pretranslational or post-translational processing (Fig. 1-3). A common mechanism of pretranslational processing includes alternative splicing of mRNA, which generates unique peptides from the same gene. Post-translational changes include cleavage of precursor molecules.
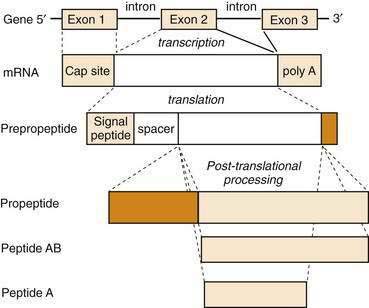
Enzymatic cleavage of the signal peptide produces a prohormone. Other post-translational features that result in mature GI peptides include peptide cleavage to smaller forms (e.g., somatostatin), amidation of the carboxyl terminus (e.g., gastrin), and sulfation of tyrosine residues (e.g., CCK). These processing steps are usually critical for biological activity of the hormone. For example, sulfated CCK is 100-fold more potent than its unsulfated form. The vast biochemical complexity of gastroenteropancreatic hormones is evident in the different tissues that secrete these peptides. As GI peptides are secreted from endocrine as well as nervous tissue, the distinct tissue involved often determines the processing steps for production of the peptide. Many hormone genes are capable of manufacturing alternatively spliced mRNAs or proteins that undergo different post-translational processing and ultimately produce hormones of different sizes. These modifications are important for receptor binding, signal transduction, and consequent cellular responses.10
It has become possible to express human genes in other species. By introducing specific hormone-producing genes into pigs or sheep, human hormones have been produced for medicinal use.11 With the rapid sequencing of the human genome, it is likely that novel methods of gene expression will expand the therapeutic use of human proteins. Moreover, drugs are being developed that inhibit the transcription of DNA into mRNA or that block the gene elements responsible for turning on specific hormone production (e.g., antisense oligonucleotides).12 This technology is based on the principle that nucleotide sequences bind to critical DNA regions and prevent transcription into mRNA. Similarly, oligonucleotides can be made to interact with mRNA and alter (or inhibit) translation of a protein product. These principles may be applicable to the treatment of the growing list of diseases that result from aberrant protein processing.13,14
GASTRIN
As discussed in more detail in Chapter 49, gastrin is the major hormone that stimulates gastric acid secretion. Subsequently, gastrin was found to have growth-promoting effects on the gastric mucosa and possibly some cancers.15 Human gastrin is the product of a single gene located on chromosome 17. The active hormone is generated from a precursor peptide called preprogastrin. Human preprogastrin contains 101 amino acids (AAs), including a signal peptide (21 AAs), spacer sequence (37 AAs), gastrin component (34 AAs), and a 9-AA extension at the carboxyl terminus. The enzymatic processing of preprogastrin produces all the known physiologically active forms of gastrin.
Preprogastrin is processed into progastrin and gastrin peptide fragments of various sizes by sequential enzymatic cleavage. The two major forms of gastrin are G34 and G17, although smaller forms exist. The common feature of all gastrins is an amidated tetrapeptide (Try-Met-Asp-Phe-NH2) carboxyl terminus, which imparts full biological activity. Modification by sulfation at tyrosine residues produces alternative gastrin forms of equal biological potency. A nonamidated form of gastrin known as glycine-extended gastrin is produced by colonic mucosa. Glycine-extended gastrin has been shown in animal models to stimulate proliferation of normal colonic mucosa and enhance the development of colorectal cancer. It is not known whether local production of this form of gastrin contributes to human colon carcinogenesis, and the receptor for glycine-extended gastrin has not been identified.16
Most gastrin is produced in endocrine cells of the gastric antrum.17 Much smaller amounts of gastrin are produced in other regions of the GI tract, including the proximal stomach, duodenum, jejunum, ileum, and pancreas. Gastrin has also been found outside the GI tract, including in the brain, adrenal gland, respiratory tract, and reproductive organs, although its biological role in these sites is unknown.
The receptors for gastrin and CCK are related and constitute the so-called gastrin-CCK receptor family. The CCK-1 and CCK-2 (previously known as CCK-A and -B) receptor complementary DNAs were cloned from the pancreas and brain, respectively, after which it was recognized that the CCK-2 receptor is identical to the gastrin receptor of the stomach.18
Hypergastrinemia occurs in pathologic states associated with decreased acid production, such as atrophic gastritis. Serum gastrin levels can also become elevated in patients on prolonged acid-suppressive medications, such as histamine receptor antagonists and proton pump inhibitors. Hypergastrinemia in these conditions is caused by stimulation of gastrin production by the alkaline pH environment. Another important but far less common cause of hypergastrinemia is a gastrin-producing tumor, also known as Zollinger-Ellison syndrome (see Chapter 32).
The gastrin analog, pentagastrin, has been used clinically to stimulate histamine and gastric acid secretion in diagnostic tests of acid secretory capacity (see Chapter 49).
CHOLECYSTOKININ
CCK is a peptide transmitter produced by I cells of the small intestine and is secreted into the blood following ingestion of a meal. Circulating CCK binds to specific CCK-1 receptors on the gallbladder, pancreas, smooth muscle of the stomach, and peripheral nerves to stimulate gallbladder contraction and pancreatic secretion, regulate gastric emptying and bowel motility, and induce satiety.19 These effects serve to coordinate the ingestion, digestion, and absorption of dietary nutrients. Ingested fat and protein are the major food components that stimulate CCK release.
CCK was originally identified as a 33–amino acid peptide. However, since its discovery larger and smaller forms of CCK have been isolated from blood, intestine, and brain. All forms of CCK are produced from a single gene by post-translational processing of a preprohormone. Forms of CCK ranging in size from CCK-58 to CCK-8 have similar biological activities.20
CCK is the major hormonal regulator of gallbladder contraction. It also plays an important role in regulating meal-stimulated pancreatic secretion (see Chapter 56) In many species, this latter effect is mediated directly through receptors on pancreatic acinar cells but in humans, in whom pancreatic CCK-1 receptors are less abundant, CCK appears to stimulate pancreatic secretion indirectly through enteropancreatic neurons that possess CCK-1 receptors. In some species, CCK has trophic effects on the pancreas, although its potential role in human pancreatic neoplasia is speculative. CCK also has been shown to delay gastric emptying.21 This action may be important in coordinating the delivery of food from the stomach to the intestine. CCK has been proposed as a major mediator of satiety and food intake, an effect that is particularly noticeable when food is in the stomach or intestine. CCK inhibits gastric acid secretion by binding to CCK-1 receptors on somatostatin (D) cells in the antrum and oxyntic mucosa. Somatostatin acts locally to inhibit gastrin release from adjacent G cells and directly inhibits acid secretion from parietal cells.22
Clinically, CCK has been used together with secretin to stimulate pancreatic secretion for pancreatic function testing. It is also used radiographically or scintigraphically to evaluate gallbladder contractility. There are no known diseases of CCK excess. Low CCK levels have been reported in individuals with celiac disease who have reduced intestinal mucosal surface area and in those with bulimia nervosa.23,24 Elevated levels of CCK have been reported in some patients with chronic pancreatitis (see Chapter 59), presumably because of reduced pancreatic enzyme secretion and interruption of negative feedback regulation of CCK release.25
SECRETIN
The first hormone, secretin, was discovered when it was observed that intestinal extracts, when injected intravenously into dogs, caused pancreatic secretion.26 Secretin is released by acid in the duodenum and stimulates pancreatic fluid and bicarbonate secretion, leading to neutralization of acidic chyme in the intestine (see Chapter 56). Secretin also inhibits gastric acid secretion (see Chapter 49) and intestinal motility.
Human secretin is a 27–amino acid peptide and, similar to many other GI peptides, is amidated at the carboxyl terminus. It is the founding member of the secretin-glucagon-VIP family of structurally related GI hormones. Secretin is selectively expressed in specialized enteroendocrine cells of the small intestine called S cells.27
One of the major physiological actions of secretin is stimulation of pancreatic fluid and bicarbonate secretion (see Chapter 56). Pancreatic bicarbonate, on reaching the duodenum, neutralizes gastric acid and raises the duodenal pH, thereby “turning off” secretin release (negative feedback). It has been suggested that acid-stimulated secretin release is regulated by an endogenous intestinal secretin-releasing factor.28 This peptide stimulates secretin release from S cells until the flow of pancreatic proteases is sufficient to degrade the releasing factor and terminate secretin release.
Although the primary action of secretin is to produce pancreatic fluid and bicarbonate secretion, it is also an enterogastrone, a substance that is released when fat is present in the GI lumen and that inhibits gastric acid secretion. In physiologic concentrations, secretin inhibits gastrin release, gastric acid secretion, and gastric motility.29 The most common clinical application of secretin is in the diagnosis of gastrin-secreting tumors,30 as discussed in Chapter 32.
VASOACTIVE INTESTINAL POLYPEPTIDE
VIP is a neuromodulator that has broad significance in intestinal physiology. VIP is a potent vasodilator that increases blood flow in the GI tract and causes smooth muscle relaxation and epithelial cell secretion.31,32 As a chemical messenger, VIP is released from nerve terminals and acts locally on cells bearing VIP receptors. VIP belongs to a family of GI peptides, including secretin and glucagon, that are structurally related. The VIP receptor is a G protein–coupled receptor that stimulates intracellular cAMP generation.
Like other GI peptides, VIP is synthesized as a precursor molecule that is cleaved to an active peptide of 28 amino acids. VIP is expressed primarily in neurons of the peripheral-enteric and central nervous systems and is released along with other peptides, including primarily PHI and/or PHM (see Table 1-1).33
VIP is an important neurotransmitter throughout the central and peripheral nervous systems.34 Because of its wide distribution, VIP has effects on many organ systems; most notably, in the GI tract, VIP stimulates fluid and electrolyte secretion from intestinal epithelium and bile duct cholangiocytes.35,36
VIP, along with NO, is a primary component of nonadrenergic, noncholinergic nerve transmission in the gut.37 GI smooth muscle exhibits a basal tone, or sustained tension, caused by rhythmic depolarizations of the smooth muscle membrane potential. VIP serves as an inhibitory transmitter of this rhythmic activity, causing membrane hyperpolarization and subsequent relaxation of GI smooth muscle. Accordingly, VIP is an important neuromodulator of sphincters of the GI tract, including the lower esophageal sphincter and sphincter of Oddi. In certain pathologic conditions, such as achalasia and Hirschsprung’s disease, the lack of VIP innervation is believed to play a major role in defective esophageal relaxation and bowel dysmotility, respectively.38,39
Unlike GI endocrine cells that line the mucosa of the gut, VIP is produced and released from neurons and it is likely that most measurable VIP in serum is of neuronal origin. Normally, serum VIP levels are low and do not appreciably change with a meal. However, in pancreatic cholera, also known as Verner-Morrison syndrome and manifested by watery diarrhea, hypokalemia, and achlorhydria,40 VIP levels can be extraordinarily high.35 VIP-secreting tumors usually produce a voluminous diarrhea41 (see Chapter 32).
GLUCAGON
Glucagon is synthesized and released from pancreatic alpha cells and from intestinal L cells of the ileum and colon. Pancreatic glucagon is a 29–amino acid peptide that regulates glucose homeostasis via gluconeogenesis, glycogenolysis, and lipolysis and is counterregulatory to insulin. The gene for glucagon encodes not only preproglucagon but also glucagon-like peptides (GLPs). This precursor peptide consists of a signal peptide, a glucagon-related polypeptide, glucagon, and GLP-1 and GLP-2. Tissue-specific peptide processing occurs through prohormone convertases that produce glucagon in the pancreas and GLP-1 and GLP-2 in the intestine (Fig. 1-4).42
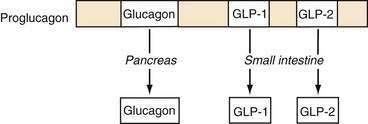
Glucagon and GLP-1 regulate glucose homeostasis.43 Glucagon is released from the endocrine pancreas in response to a meal and binds to G protein–coupled receptors on skeletal muscle and the liver to exert its glucoregulatory effects. GLP-1 stimulates insulin secretion and augments the insulin-releasing effects of glucose on the pancreatic beta cell (see later, “Enteroinsular Axis”). GLP-1 analogs have been developed for the treatment of type II diabetes mellitus. A long-acting human GLP-1 analog improves beta cell function and can lower body weight in patients with type II diabetes.44,45 GLP-2 is an intestinal growth factor and may have therapeutic implications in the maintenance of the GI mucosal mass and the reversal of villus atrophy.
GLUCOSE-DEPENDENT INSULINOTROPIC POLYPEPTIDE
GIP was discovered based on its ability to inhibit gastric acid secretion (enterogastrone effect) and was originally termed gastric inhibitory polypeptide. It was subsequently shown that the effects on gastric acid secretion occur only at very high concentrations that are above the physiologic range. However, GIP has potent effects on insulin release that (like GLP-1) potentiates glucose-stimulated insulin secretion.46 Based on this action, GIP was redefined as glucose-dependent insulinotropic polypeptide.
GIP receptors are also expressed on adipocytes through which GIP augments triglyceride storage, which may contribute to fat accumulation. Based on the insulinotropic properties of GIP, coupled with its effects on adipocytes, it has been proposed that GIP may play a role in obesity and development of insulin resistance associated with type II diabetes mellitus.47 Consistent with this proposal was the experimental finding that mice lacking the GIP receptor do not gain weight when placed on a high-fat diet.48 It remains to be seen whether GIP antagonists can be used to treat obesity. In rare circumstances, receptors for GIP may be aberrantly expressed in the adrenal cortex, resulting in food-dependent Cushing’s syndrome.49,50
PANCREATIC POLYPEPTIDE FAMILY
Originally isolated during the preparation of insulin, pancreatic polypeptide (PP) is the founding member of the PP family.51 The PP family of peptides includes NPY and peptide tyrosine tyrosine (PYY), which were discovered because of the presence of a C-terminal tyrosine amide.52,53 PP is stored and secreted from specialized pancreatic endocrine cells (PP cells),54 whereas NPY is a principal neurotransmitter found in the central and peripheral nervous systems.55 PYY has been localized to enteroendocrine cells throughout the GI tract but is found in greatest concentrations in the ileum and colon.56
The PP-PYY-NPY family of peptides functions as endocrine, paracrine, and neurocrine transmitters in the regulation of a number of actions that result from binding to one of five receptor subtypes.57 PP inhibits pancreatic exocrine secretion, gallbladder contraction, and gut motility.58 PYY inhibits vagally stimulated gastric acid secretion and other motor and secretory functions.59 An abbreviated form of PYY lacking the first two amino acids of the normally produced 36 amino acid peptide, PYY3-36, has been shown to reduce food intake when administered to humans, indicating that intestinally released peptide may play a role in regulating meal size.60 NPY is one of the most abundant peptides in the central nervous system and, in contrast to PYY3-36, is a potent stimulant of food intake.61 Peripherally, NPY affects vascular and GI smooth muscle function.62
SUBSTANCE P AND THE TACHYKININS
Substance P belongs to the tachykinin family of peptides, which includes neurokinin A and neurokinin B. The tachykinins are found throughout the peripheral and central nervous systems, and are important mediators of neuropathic inflammation.63 Tachykinins, as a group, are encoded by two genes that produce preprotachykinin A and preprotachykinin B. Common to both is a well-conserved C-terminal pentapeptide. Transcriptional and translational processing produce substance P, neurokinin A, and/or neurokinin B, which are regulated in large part by alternative splicing. These peptides function primarily as neuropeptides. Substance P is a neurotransmitter of primary sensory afferent neurons and binds to specific receptors in lamina I of the spinal cord.64 Three receptors for this family of peptides have been identified—NK-1, NK-2, and NK-3.65 Substance P is the primary ligand for the NK-1 receptor, neurokinin A for the NK-2 receptor, and neurokinin B for the NK-3 receptor. However, all these peptides can bind and signal through all three receptor subtypes.
Substance P has been implicated as a primary mediator of neurogenic inflammation. In the intestine, Clostridium difficile–initiated experimental colitis results from toxin-induced release of substance P and consequent activation of the NK-1 receptor.66 These inflammatory sequelae can be blocked by substance P receptor antagonists. Substance P receptors are more abundant in the intestine of patients with ulcerative colitis and Crohn’s disease.67
SOMATOSTATIN
Somatostatin is a 14–amino acid cyclic peptide that was initially identified as an inhibitor of growth hormone secretion. Since its discovery, it has been found in almost every organ in the body and throughout the GI tract. In the gut, somatostatin is produced by D cells in the gastric and intestinal mucosa and islets of the pancreas, as well as enteric neurons.68 Somatostatin has a number of pharmacologic effects that are mostly inhibitory.
In the stomach, somatostatin plays an important role in regulating gastric acid secretion.69 In the antrum, D cells are open to the lumen, where they are directly exposed to acid. A low gastric pH stimulates D cells that lie in close proximity to gastrin-producing cells to secrete somatostatin and inhibit gastrin release (see Chapter 49). Reduced gastrin secretion decreases the stimulus for acid production and the pH of the stomach contents rises. Thus, some of the inhibitory effects of gastric acid on gastrin release (see earlier, “Gastrin”) are mediated by somatostatin.
Somatostatin release is also influenced by mechanical stimulation, dietary components of a meal, including protein, fat, and glucose, and other hormones and neurotransmitters.70 Muscarinic stimulation appears to be the most important neural stimulus to somatostatin secretion.
At least five somatostatin receptors have been identified that account for divergent pharmacologic properties.71 For example, receptor subtypes 2 and 3 couple to inhibitory G proteins but receptor subtype 1 does not. In addition, only somatostatin receptor subtype 3 inhibits adenylate cyclase. The inhibitory effects of somatostatin are mediated by a decrease in cAMP, Ca2+ channel inhibition, or K+ channel opening.
In the gut, somatostatin has broad inhibitory actions. In addition to effects on gastric acid, somatostatin reduces pepsinogen secretion. Somatostatin profoundly inhibits pancreatic enzyme, fluid, and bicarbonate secretion and reduces bile flow.72 The effects of somatostatin on gut motility are largely inhibitory, with the exception that it stimulates the migrating motor complex, possibly through effects on motilin. Somatostatin also reduces intestinal transport of nutrients and fluid, reduces splanchnic blood flow, and has inhibitory effects on tissue growth and proliferation.73,74
Because of its varied physiologic effects, somatostatin has several clinically important pharmacologic uses. Many endocrine cells possess somatostatin receptors and are sensitive to inhibitory regulation. Therefore, somatostatin and more recently developed somatostatin analogs are used to treat conditions of hormone excess produced by endocrine tumors, such as acromegaly, carcinoid tumors, and islet cell tumors (including gastrinomas).75 Its ability to reduce splanchnic blood flow and portal venous pressure has led to somatostatin analogs being useful in treating esophageal variceal bleeding (see Chapter 90).76 The inhibitory effects on secretion have been exploited by using somatostatin analogs to treat some forms of diarrhea and reduce fluid output from pancreatic fistulas. Many endocrine tumors express abundant somatostatin receptors, making it possible to use radiolabeled somatostatin analogs, such as octreotide, to localize even small tumors throughout the body.
MOTILIN
Motilin is a 22–amino acid peptide produced by endocrine cells of the duodenal epithelium.77 Motilin is secreted into the blood in a periodic and recurrent pattern that is synchronized with the migrating motor complex (MMC) under fasting conditions. Elevations in blood motilin levels regulate the phase III contractions that initiate in the antroduodenal region and progress toward the distal gut. Motilin secretion is not stimulated by eating.
Motilin binds to specific receptors on smooth muscle cells of the esophagus, stomach, and small and large intestines through which it exerts propulsive activity.78 Agonists to the motilin receptor such as erythromycin have pronounced effects on GI motility, which occasionally produces undesired side effects of abdominal cramping and diarrhea.79 However, motilin agonists may be useful to treat conditions of impaired gastric and intestinal motility and are being investigated for the treatment of constipation-predominant irritable bowel syndrome.80
LEPTIN
Leptin is a 167–amino acid protein that is secreted primarily from adipocytes. Blood leptin levels reflect total body fat stores.81 Its primary action appears to be to reduce food intake. Leptin is a member of the cytokine family of signaling molecules. Five different forms of leptin receptors have been reported.82 A short form of the receptor appears to transport leptin from the blood across the blood-brain barrier, where it has access to the hypothalamus. A long form of the leptin receptor is located in hypothalamic nuclei, where leptin binds and activates the Janis kinase signal transduction and translation system (JAK STAT).83 Small amounts of leptin are produced by the chief cells of the stomach and by the placenta, and are present in breast milk.
Peripheral administration of leptin reduces food intake. However, this effect is reduced as animals become obese. Interestingly, when injected into the central nervous system, obese animals respond normally to leptin and reduce food intake, suggesting that leptin “resistance” in obesity occurs at the level of the leptin receptor that transports leptin across the blood-brain barrier.84 Leptin’s ability to reduce food intake occurs within the brain by decreasing NPY (a potent stimulant of food intake) and by increasing α–melanocyte-stimulating hormone (α−MSH), an inhibitor of food intake.85 Peripherally, leptin acts synergistically with cholecystokinin to reduce meal size.86 In obese rats lacking the leptin receptor, the synergistic effects of leptin plus CCK to reduce meal size are lost, but could be restored with genetic reconstitution of the leptin receptor in the brain.87 One might expect loss of leptin-CCK synergy on meal size in those rare cases of human obesity caused by leptin receptor defects or even with leptin resistance.
Blood levels of leptin increase as obesity develops and leptin appears to reflect total fat content.88 At the cellular level, large adipocytes produce more leptin than small adipocytes. Because of its effects on food intake, it was initially thought that exogenous leptin could be used therapeutically to treat obesity. However, only a very modest effect on weight loss has been demonstrated in clinical trials. Leptin deficiency has been reported as a cause of obesity in a few families, but this condition is extremely rare.89,90 Mutation of the leptin receptor has been described as a cause of obesity in at least one family.91
GHRELIN
Ghrelin is a 28–amino acid peptide produced by the stomach and is the natural ligand for the growth hormone secretagogue (GHS) receptor.92 When administered centrally or peripherally ghrelin stimulates growth hormone secretion, increases food intake, and produces weight gain.93,94 Circulating ghrelin levels increase during periods of fasting or under conditions associated with negative energy balance, such as starvation or anorexia. In contrast, ghrelin levels are low after eating and in obesity. Ghrelin appears to play a central role in the neurohormonal regulation of food intake and energy homeostasis.
The gastric fundus is the most abundant source of ghrelin, although lower amounts of ghrelin are found in the intestine, pancreas, pituitary, kidney, and placenta. Ghrelin is produced by distinctive endocrine cells known as P/D1 cells95,96 that are of two types, open and closed. The open type is exposed to the lumen of the stomach, where it comes into contact with gastric contents, whereas the closed type lies in close proximity to the capillary network of the lamina propria.97 Both cell types secrete hormone into the bloodstream. Based on its structure, ghrelin is a member of the motilin family of peptides and, like motilin, ghrelin stimulates gastric contraction and enhances stomach emptying.
The observations that circulating ghrelin levels increase sharply before a meal and fall abruptly after a meal suggest that it serves as a signal for initiation of feeding. The effects of food on plasma ghrelin levels can be reproduced by ingestion of glucose and appear to be unrelated to the physical effects of a meal on gastric distention. Circulating ghrelin levels are low in states of positive energy balance such as obesity and are inversely correlated with body mass index.98,99 Conversely, ghrelin levels are high in fasting, cachexia, and anorexia. Importantly, weight loss increases circulating ghrelin levels.100
Ghrelin released from the stomach acts on the vagus nerve to exert its effects on feeding. However, it is also active when delivered to the central nervous system and, in this location, ghrelin activates NPY and agouti-related protein-producing neurons in the arcuate nucleus of the hypothalamus, which is involved in the regulation of feeding.94,101
Gastric bypass patients do not demonstrate the premeal increase in plasma ghrelin that is seen in normal individuals.102 This lack of ghrelin release may be one of the mechanisms contributing to the overall effectiveness of gastric bypass surgery for inducing weight loss.
Prader-Willi syndrome is a congenital obesity syndrome characterized by severe hyperphagia, growth hormone deficiency, and hypogonadism. Although obesity is ordinarily associated with low ghrelin levels, patients with Prader-Willi syndrome have high circulating ghrelin levels that do not decline after a meal.103,104 The levels of ghrelin in this syndrome are similar to those that can stimulate appetite and increase food intake in individuals receiving infusions of exogenous ghrelin, suggesting that abnormal ghrelin secretion may be responsible for the hyperphagia in Prader-Willi syndrome.105
OTHER CHEMICAL MESSENGERS OF THE GASTROINTESTINAL TRACT
ACETYLCHOLINE
Acetylcholine is synthesized in cholinergic neurons and is the principal regulator of GI motility and pancreatic secretion. Acetylcholine is stored in nerve terminals and released by nerve depolarization. Released acetylcholine binds to postsynaptic muscarinic and/or nicotinic receptors. Nicotinic acetylcholine receptors belong to a family of ligand-gated ion channels and are homopentamers or heteropentamers composed of α, β, γ, δ, and ε subunits.106 The α subunit is believed to be the mediator of postsynaptic membrane depolarization following acetylcholine receptor binding. Muscarinic receptors belong to the heptahelical GPCR family. There are five known muscarinic cholinergic receptors (M1 to M5). Muscarinic receptors can be further classified based on receptor signal transduction, with M1, M3, and M5 stimulating adenylate cyclase and M2 and M4 inhibiting this enzyme. Acetylcholine is degraded by the enzyme acetylcholinesterase, and the products may be recycled through high-affinity transporters on the nerve terminal.
DOPAMINE
Dopamine is an important mediator of GI secretion, absorption, and motility and is the predominant catecholamine neurotransmitter of the central and peripheral nervous systems. In the central nervous system, dopamine regulates food intake, emotions, and endocrine responses and, peripherally, it controls hormone secretion, vascular tone, and GI motility. Characterization of dopamine in the GI tract has been challenging for several reasons. First, dopamine can produce inhibitory and excitatory effects on GI motility.107 Generally, the excitatory response, which is mediated by presynaptic receptors, occurs at a lower agonist concentration than the inhibitory effect, which is mediated by postsynaptic receptors. Second, localization of dopamine receptors has been hampered by identification of dopamine receptors in locations that appear to be species specific.108 Third, studies of dopamine in GI tract motility have often used pharmacologic amounts of this agonist. Therefore, the interpretation of results has been confounded by the ability of dopamine to activate adrenergic receptors at high doses.
Classically, dopamine was thought to act via two distinct receptor subtypes, type 1 and type 2. Molecular cloning has now demonstrated five dopamine receptor subtypes, each with a unique molecular structure and gene locus.108 Dopamine receptors are integral membrane GPCRs, and each receptor subtype has a specific pharmacologic profile when exposed to agonists and antagonists. After release from the nerve terminal, dopamine is cleared from the synaptic cleft by a specific dopamine transporter.
SEROTONIN
Serotonin has long been known to play a role in GI neurotransmission.109 The GI tract contains more than 95% of the total body serotonin, and serotonin is important in various processes, including epithelial secretion, bowel motility, nausea and emesis.110 Serotonin is synthesized from tryptophan, an essential amino acid, and is converted to its active form in nerve terminals. Secreted serotonin is inactivated in the synaptic cleft by reuptake via a serotonin-specific transporter. Most plasma serotonin is derived from the gut, where it is found in mucosal enterochromaffin cells and the enteric nervous system. Serotonin mediates its effects by binding to a specific receptor. There are seven different serotonin receptor subtypes found on enteric neurons, enterochromaffin cells, and GI smooth muscle (5-HT1 to 5-HT7).
The actions of serotonin are complex (Fig. 1-5).111 It can cause smooth muscle contraction through stimulation of cholinergic nerves or relaxation by stimulating inhibitory NO-containing neurons.110 Serotonin released from mucosal cells stimulates sensory neurons, initiating a peristaltic reflex and secretion (via 5-HT4 receptors) and modulates sensation through activation of 5-HT3 receptors.109 The myenteric plexus contains serotoninergic interneurons that project to the submucosal plexus and ganglia extrinsic to the bowel wall. Extrinsic neurons activated by serotonin participate in bowel sensation and may be responsible for abdominal pain, nausea, and symptoms associated with irritable bowel syndrome. Intrinsic neurons activated by serotonin are primary components of the peristaltic and secretory reflexes responsible for normal GI function. Serotonin may also activate vagal afferent pathways and, in the central nervous system, modulates appetite, mood, and sexual function. Because of these diverse effects, it is not surprising that selective serotonin reuptake inhibitor drugs (SSRIs), commonly used to treat depression and anxiety, have prominent GI side effects when compared with placebo treatment.
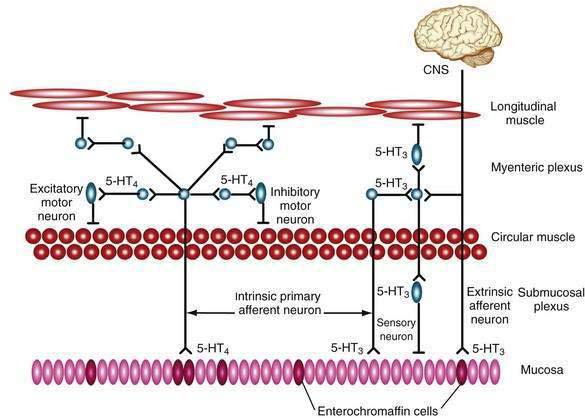
Serotonin and its receptor have been implicated in the pathogenesis of motility disorders of the GI tract.112 Characterization of specific serotonin receptor subtypes has led to the development of selective agonists and antagonists for the treatment of irritable bowel syndrome and chronic constipation and diarrhea. For example, 5-HT3 receptor antagonists, which reduce intestinal secretion, are used to treat diarrhea-predominant irritable bowel syndrome. 5-HT4 receptor agonists elicit prokinetic effects and are used to treat constipation-predominant irritable bowel syndrome and other motility disorders.113,114
Serotonin can also be enzymatically converted to melatonin by serotonin N-acetyltransferase.115 Other than the pineal gland, the GI tract is the major source of the body’s melatonin. Melatonin is produced in enterochromaffin cells and released into the blood after ingestion of a meal. A number of actions on the GI tract have been described for melatonin, including reducing gastric acid and pepsin secretion, inducing smooth muscle relaxation, and preventing epithelial injury through an antioxidant effect.116 It has been proposed that melatonin released after a meal may contribute to postprandial somnolence.117
HISTAMINE
In the GI tract, histamine is best known for its central role in regulating gastric acid secretion (see Chapter 49) and intestinal motility. Histamine is produced by enterochromaffin-like cells of the stomach and intestine as well as enteric nerves. Histamine is synthesized from l-histidine by histidine decarboxylase and activates three GPCR subtypes. H1 receptors are found on smooth muscle and vascular endothelial cells and are linked to phospholipase C (PLC) activation. As such, the H1 receptor mediates many of the allergic responses induced by histamine. H2 receptors are present on gastric parietal cells, smooth muscle, and cardiac myocytes. H2 receptor binding stimulates Gs (G proteins that stimulate adenylate cyclase) and activates adenylate cyclase. H3 receptors are present in the central nervous system and GI tract enterochromaffin cells. These receptors signal through Gi and inhibit adenylate cyclase.118 Histamine can also interact with the N-methyl-d-aspartate (NMDA) receptor and enhance activity of NMDA-bearing neurons independently of the three known histamine receptor subtypes.
NITRIC OXIDE
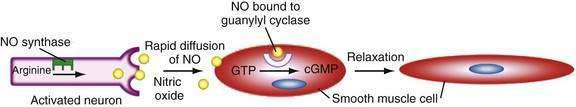
NO is a unique chemical messenger produced from l-arginine by the enzyme nitric oxide synthase (NOS).119 Three types of NOS are known. Types I and III are also known as endothelial NOS and neuronal NOS, respectively, and are constitutively active. Small changes in NOS activity can occur through elevations in intracellular calcium. The inducible form of NOS (type II) is apparent only when cells become activated by specific inflammatory cytokines. This form of NOS is capable of producing large amounts of NO and is calcium-independent. NOS is often colocalized with VIP and PACAP in neurons of the enteric nervous system.120
NO, being an unstable gas, has a relatively short half-life. Unlike most neurotransmitters and hormones, NO does not act via a membrane-bound receptor. Instead, NO readily diffuses into adjacent cells to activate guanylate cyclase directly (Fig. 1-6). NO activity is terminated by its oxidation to nitrate and nitrite. Many enteric nerves use NO to signal neighboring cells and induce epithelial secretion, vasodilation, or muscle relaxation. NO is also produced by macrophages and neutrophils to help kill invading organisms.121
ADENOSINE
Adenosine is an endogenous nucleoside that acts through any of four GPCR subtypes.122 Adenosine causes relaxation of intestinal smooth muscle and stimulates intestinal secretion. Adenosine can also cause peripheral vasodilation and activation of nociceptors that participate in neural pain pathways.
CYTOKINES
Cytokines are a group of polypeptides produced by various immunomodulatory cells and are involved in cell proliferation, immunity, and inflammation. Cytokines are induced by specific stimuli, such as toxins produced by pathogens, and often elicit a complex response involving other cellular mediators to eradicate the foreign substance. Cytokines may be categorized as interleukins (ILs), tumor necrosis factors (TNFs), lymphotoxins, interferons, colony-stimulating factors (CSFs), and others.123 Interleukins can be further subtyped into at least 35 separate substances, IL-1 to IL-35. There are two TNFs, TNF-α and TNF-β, which are also known as lymphotoxin-α. Interferons are produced during viral or bacterial infection and come in two varieties, interferon-α (also known as leukocyte-derived interferon or interferon-β) and interferon-γ. Interferon-α is produced by T lymphocytes and is used clinically for the treatment of viral hepatitis (see Chapters 78 and 79). The major CSFs are granulocyte mononuclear phagocyte CSF, mononuclear phagocyte CSF, and granulocyte CSF. These agents are used for chemotherapy-induced neutropenia and marrow support after bone marrow transplantation. Chemokines initiate and propagate inflammation and are of two groups, CXC (α chemokines) and CC (β chemokines). Other cytokines, such as transforming growth factor-β (TGF)-β and platelet-derived growth factor (PDGF), have proliferative effects.
SIGNAL TRANSDUCTION
G PROTEIN–COUPLED RECEPTORS
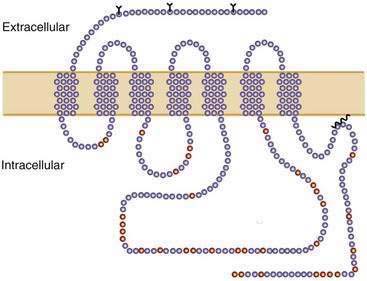
GPCRs are seven membrane-spanning domain proteins associated with a heterotrimeric G protein (Fig. 1-7). The membrane regions consist of α-helical domains with a conserved structural motif.124 GPCRs contain an extracellular amino terminus and an intracellular carboxyl terminus (see Fig. 1-7). When stimulated by the appropriate chemical messenger, the GPCR undergoes a conformational change and couples to a specific G protein. The first crystal structure of a GPCR, for rhodopsin, was elucidated in 2000.125 The three-dimensional structure of the rhodopsin receptor reveals a highly organized heptahelical transmembrane component with a portion of the C-terminus perpendicular to the seventh and final membrane-spanning domains of the protein.
G PROTEINS
G proteins are molecular intermediaries that initiate the intracellular communication process on ligand binding to its GPCR (Fig. 1-8).126 G proteins are composed of three subunits—α, β, and γ—and are classified according to their α subunit. They activate various effector systems, including adenylate cyclase, guanylate cyclase, phospholipases, and specific ion channels.127 G proteins that stimulate adenylate cyclase are classified as Gs; those that inhibit adenylate cyclase are called Gi.128
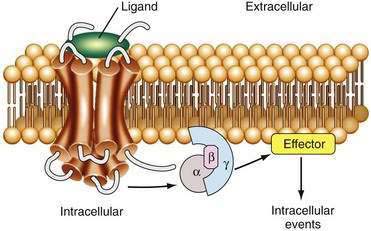
When an agonist binds to a Gs-coupled receptor, a conformational change occurs, allowing the receptor to associate with the Gαs subunit. Under basal (unstimulated) conditions, Gαs is bound to guanosine diphosphate (GDP); however, with agonist binding, GDP is released and replaced with guanosine triphosphate (GTP). The Gs-GTP complex then activates adenylate cyclase, resulting in the generation of cAMP from adenosine triphosphate (ATP) within the cell cytoplasm. cAMP phosphorylates effector proteins that ultimately lead to responses such as secretion, cell movement, and growth. Receptor activation also initiates the dissociation of the α subunit from the βγ subunits. However, the βγ subunits remain tightly associated and themselves participate in a vast array of cellular signals. For example, not only can βγ subunits activate GPCR kinases, adenylate cyclase, and ion channels, they induce receptor desensitization and stimulate Ras-mediated mitogen-activated protein (MAP) kinase.129,130
Certain GPCRs activate an inhibitory G protein (Gαi) that inhibits cAMP accumulation and antagonizes the effects of Gs-coupled events. In this manner, GPCRs can maintain fine control of the cellular cAMP concentration and subsequent intracellular signaling. Members of this GPCR family also activate phospholipases and phosphodiesterases, and are often involved with ion channel regulation. Other GPCRs couple with Gq and G12 (see Table 1-2). The Gq family of G protein subunits regulates the production of inositol 1,4,5-trisphosphate (IP3) and diacylglycerol (DAG).131 Following α subunit dissociation from βγ, when the α subunit reverts to the GDP-bound form, it reassociates with βγ. With reestablishment of the αβγ heterotrimer, along with other mechanisms of desensitization, receptor signaling via the separate subunits ceases.
Table 1-2 Classification of G Protein α Subunits and Their Signaling Pathways
| CLASS | SIGNALING |
|---|---|
| Gαs | Adenylate cyclase, calcium channels |
| Gαi and Gαo | Adenylate cyclase, cyclic guanosine monophosphate, phosphodiesterase, c-Src, STAT 3 |
| Gαq | Phospholipase C-β |
| Gα12 and Gα13 | Sodium-hydrogen exchange |
Effector Systems
Following receptor occupation, G protein subunits cause activation of enzymes or other proteins, ultimately resulting in intracellular signaling events (Table 1-2). Enzymes such as adenylate cyclase or phospholipase C generate specific second messengers such as cAMP or IP3 and DAG. Some G proteins couple directly to specific ion channels, such as potassium or calcium channels, and initiate changes in ion permeability. The effector systems are not well understood for some receptors, such as those involved with cell growth and differentiation.
Receptor Desensitization
Phosphorylation of the receptor is one of the initial events involved in turning off the signal after agonist binding and occurs through binding of arrestin-like molecules, which uncouple the receptor from the G protein.132 This uncoupling and subsequent receptor internalization (sequestration) continue the process of signal termination and eventually lead to the reestablishment of cell responsiveness.
RECEPTORS NOT COUPLED TO G PROTEINS
Enzyme-Coupled Receptors
Receptor Tyrosine Kinases
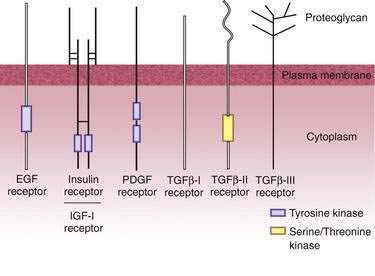
Unlike GPCRs, where ligand-receptor interaction causes activation of a G protein intermediary, some ligand receptors possess intrinsic protein tyrosine kinase activity. These membrane-spanning cell surface receptors catalyze the transfer of phosphate from ATP to target proteins. Such receptors are structurally unique in that they contain glycosylated extracellular binding domains, a single transmembrane domain, and a cytoplasmic domain. The cytoplasmic domain contains a protein tyrosine kinase region and substrate region for agonist-activated receptor phosphorylation. With activation, these receptors may phosphorylate themselves or be phosphorylated by other protein kinases.133 In general, receptor tyrosine kinases exist in the cell membrane as monomers. However, with ligand binding, these receptors dimerize, autophosphorylate, and initiate other intracellular signal transduction pathways. Most receptor tyrosine kinases couple, via ligand binding, to Ras and subsequently activate MAP kinase. MAP kinase is then able to modulate the regulation of other cellular proteins, including transcription factors. Members of the receptor tyrosine kinase family include the insulin receptor, growth factor receptors (vascular endothelial growth factor, PDGF, epidermal growth factor [EGF], fibroblast growth factor [FGF], insulin-like growth factor I [IGF] I, macrophage-CSF, nerve growth factor), and receptors involved in development.134 Receptor tyrosine kinases are discussed further in Chapter 3 in relation to cellular growth and neoplasia.
Activated tyrosine kinase receptors participate in a number of intracellular signaling events that involve the phosphorylated cytoplasmic domain. Specific phosphorylated tyrosine residues serve as binding sites for Src homology regions 2 and 3 (SH2 and SH3 domains). The result of SH2 domain binding is activation or modulation of the signaling protein that contains this binding domain. In this manner, receptor tyrosine kinases activate diverse signaling pathways.135
Receptor Guanylate Cyclases
Receptor guanylate cyclases use cyclic GMP (cGMP) as a direct intracellular mediator. These cell surface receptors contain an extracellular ligand-binding region, a single transmembrane domain, and a cytoplasmic guanylate cyclase catalytic domain.136 Ligand stimulation of a receptor guanylate cyclase results in activation of cGMP-dependent protein kinase, which is a serine-threonine protein kinase. The atrial natriuretic peptide (ANP) receptor is a representative receptor guanylate kinase, which mediates the potent smooth muscle relaxing activity of ANP.
Nonreceptor Tyrosine Kinases
Some cell surface receptors involved in inflammation and hematopoietic cell regulation work through tyrosine kinases but do not contain a cytoplasmic catalytic domain. The Src family of kinases is the primary component of this receptor signaling system.137
Receptor Tyrosine Phosphatases
Leukocyte regulation is modulated by surface receptors whose function is to remove phosphate groups from specific phosphotyrosines. CD45 is a surface protein found in white blood cells that participates in T and B cell activation.138 CD45 contains a single membrane-spanning domain and a cytoplasmic region with tyrosine phosphatase activity. Depending on the substrate, dephosphorylation of signaling proteins may result in reduced or enhanced activity. Receptors in this family are important in inflammation and immune regulation and have been shown to participate in GI development, growth, and cancer.
Receptor Serine-Threonine Kinases
TGF-β (see “Growth Factor Receptors”) receptors are a unique group of surface proteins that are involved in various cell functions, including chemotaxis, inflammation, and proliferation. These receptors contain a single membrane domain and a cytoplasmic serine-threonine kinase region. Receptor stimulation initiates activation of the serine-threonine kinase and subsequent modulation of cellular protein function.139
HORMONE AND TRANSMITTER REGULATION OF GASTROINTESTINAL GROWTH
Growth of GI tissues is a balance between cellular proliferation and senescence. Many factors participate in maintenance of the GI mucosa. Nutrients and other luminal factors stimulate growth of the intestinal mucosa and are necessary to maintain normal digestive and absorptive functions. Hormones and transmitters serve as secondary messengers that are normally secreted in response to food ingestion and mediate many of the nutrient effects on the GI tract. They play a key role in cellular proliferation. Alterations in intestinal proliferation are manifested by atrophy, hyperplasia, dysplasia, or malignancy (see Chapter 3).
Growth factors that have important effects on the GI tract include peptides of the EGF, TGF-β, IGF, FGF, and PDGF families, hepatocyte growth factors, trefoil factors, and many cytokines (including interleukins).140
GROWTH FACTOR RECEPTORS
Growth factors regulate cellular proliferation by interacting with specific cell surface receptors. These receptors are membrane proteins that possess specific binding sites for the growth factor ligand. An unusual form of signaling occurs when the ligand interacts with its receptor within the same cell. For example, PDGF receptors present on the intracellular surface of fibroblast cell lines are activated by intracellular ligand. This process is known as intracrine signaling. Most peptide growth factors, however, interact with receptors on different cells to regulate proliferation. Growth factor receptors can be single polypeptide chains containing one membrane-spanning region, such as the receptor for EGF, or they may be composed two subunit heterodimers, with one subunit containing a transmembrane domain and the other residing intracellularly but covalently bound to the transmembrane subunit (Fig. 1-9). Heterodimers may also dimerize to form a receptor composed of four subunits (e.g., IGF receptor). Binding of the ligand to its receptor usually causes aggregation of two or more receptors and activation of intrinsic tyrosine kinase activity. Growth factor receptors also have the ability to autophosphorylate when bound to ligand. In addition, receptor tyrosine kinase activity may phosphorylate other intracellular proteins important in signal transduction. Autophosphorylation attenuates the receptor’s kinase activity and often leads to down-regulation and internalization of the receptor. Mutation of the receptor at its autophosphorylation site may lead to constitutive receptor activity and cellular transformation. Growth factor receptors may couple to various intracellular signaling pathways, including adenylate cyclase, phospholipase C, calcium-calmodulin protein kinases, MAP kinase, and nuclear transcription factors. Thus, growth factors play important and varied roles in most cells of the GI tract. It is not surprising, therefore, that mutations in growth factor receptors or downstream signaling proteins can lead to unregulated cell growth and neoplasia (see Chapter 3).
An important action of growth factors is their ability to modulate the expression of transacting transcription factors that can regulate expression of many other genes.141 Early response genes such as jun and fos are activated rapidly after ligand binding and control the expression of many other genes involved in cellular proliferation. Other important transcriptional factors include c-myc and nuclear factor κB (NF-κB). The latter is found in the cytoplasm in an inactive form and, following ligand binding, translocates to the nucleus, where it activates other transcription factors. NF-κB is a key target for strategies to regulate cellular proliferation and inflammation. In its phosphorylated form Rb-1, originally identified in retinoblastoma, is an inhibitor of cellular proliferation that complexes with the transcription factor p53. Dephosphorylation of Rb-1 releases p53, which activates other genes leading to cellular proliferation.
EPIDERMAL GROWTH FACTOR
EGF was the first growth factor to be discovered. It is the prototype for a family of growth factors that are structurally related and have similarly related receptors. Other members of the family include TGF-α, amphiregulin, and heparin-binding EGF. EGF is identical to urogastrone (originally isolated from urine), which was shown to inhibit gastric acid secretion and promote healing of gastric ulcers. EGF is secreted from submaxillary glands and Brunner’s glands of the duodenum. It is likely that EGF interacts with luminal cells of the GI tract to regulate proliferation. EGF has important trophic effects on gastric mucosa, and the wide distribution of EGF receptors suggests that EGF has mitogenic actions on various cells throughout the gut. The EGF receptor has been reported to be responsible for gastric hyperplasia in patients with Ménétrier’s disease.142 Moreover, two patients were effectively treated with a monoclonal antibody that blocks ligand binding to the EGF receptor.143
EGF receptors are considered important targets for the experimental treatment of human cancer based on the evidence that they play a critical role in the growth and survival of certain tumors. Monoclonal antibodies as well as small tyrosine kinase inhibitors have been undergoing clinical evaluation for the treatment of human tumors.144
TRANSFORMING GROWTH FACTOR-α
TGF-α is produced by most epithelial cells of the GI tract and acts through the EGF receptor. Therefore, it shares trophic properties with EGF. It is believed to play a key role in gastric reconstitution after mucosal injury. Moreover, it appears to be important in intestinal neoplasia because most gastric and colon cancers produce TGF-α (see Chapters 54 and 123).
TRANSFORMING GROWTH FACTOR-β
A family of TGF-β peptides exerts various biological actions, including stimulation of proliferation, differentiation, embryonic development, and formation of extracellular matrix.139 In contrast with the TGF-α receptor, there are three distinct TGF-β receptors (see Fig. 1-9).145 TGF-β modulates cell growth and proliferation in nearly all cell types and can enhance its own production from cells. It is likely that TGF-β plays a critical role in inflammation and tissue repair. TGF-β augments collagen production by recruitment of fibroblasts through its chemoattractant properties. This action can have beneficial or deleterious effects, depending on its site of deposition and abundance. For example, TGF-β may play a key role in the development of adhesions following surgery.146
INSULIN-LIKE GROWTH FACTORS
Alternative splicing of the insulin gene produces two structurally related peptides, IGF I and IGF II.147 IGFs signal through at least three different IGF receptors. The IGF I receptor is a tyrosine kinase, and the IGF II receptor is identical to the mannose 6-phosphate receptor. Although the exact function of IGFs in the GI tract is not clearly understood, they have potent mitogenic activity in intestinal epithelium. IGF II appears to be critical for embryonic development.
FIBROBLAST GROWTH FACTOR AND PLATELET-DERIVED GROWTH FACTOR
At least seven related FGFs have been identified.148 These peptides have mitogenic effects on various cell types, including mesenchymal cells, and likely play an important role in organogenesis and neovascularization.149 Although not unique to the GI tract, PDGF is one of the most thoroughly studied growth factors. It is important for fibroblast growth, and its receptor is expressed in the liver and throughout the GI tract, where it appears to promote wound healing.
TREFOIL FACTORS
Trefoil factors (pS2, spasmolysin, and intestinal trefoil factor, also known as TTF1, 2, and 3, respectively) are a family of proteins expressed throughout the GI tract.150 They share a common structure, having six cysteine residues and three disulfide bonds, creating a cloverleaf appearance that stabilizes the peptide within the gut lumen. The pS2 peptide is produced in the gastric mucosa, spasmolysin is found in the antrum and pancreas, and intestinal trefoil factor is produced throughout the small and large intestines. These peptides are produced by mucous neck cells in the stomach or goblet cells in the intestine and are secreted onto the mucosal surface of the gut. It is likely that trefoil factors act on the apical surface of the epithelial cells, where they have growth-promoting properties on the GI mucosa.
OTHER G PROTEIN–COUPLED RECEPTORS
Other peptides signaling through GPCRs may also have growth-promoting effects. Three important examples include gastrin, CCK, and gastrin-releasing peptide (GRP). Gastrin stimulates the growth of enterochromaffin-like cells of the stomach and induces proliferation of the oxyntic mucosa containing parietal cells.151 Gastrin binds to CCK-2 receptors of the stomach and activates PLC and Ras pathways, which ultimately results in activation of protein kinase C and MAP kinase, respectively. MAP kinase, which can also be activated by tyrosine kinase receptors typical of growth factors, causes the phosphorylation of transcription factors that are involved in cellular proliferation. In some cells, cAMP and protein kinase A exert synergistic effects on cellular growth through activation of nuclear transcription factors such as cAMP-responsive element binding (protein) (CREB). However, in other cells, cAMP antagonizes proliferation. Therefore, depending on the cell type, the effects of growth factors such as EGF, IGF, and PDGF may be enhanced by hormones that stimulate cAMP production. Certain colon cancer cells possess CCK-2 receptors and respond to the proliferative effects of gastrin. Moreover, gastrin may be produced by some colon cancers, enabling it to exert an autocrine effect to promote cancer growth.152 Whether circulating gastrin initiates colon cancer development is unknown.
CCK binds preferentially to the CCK-1 type receptor, which is abundant in gallbladder, the pancreas of many species, brain, and peripheral nerves of the gut. In the rodent, but not human, pancreas, CCK causes hypertrophy and hyperplasia of pancreatic acinar cells. Similar to the effects of gastrin, CCK activates phospholipase C and small GTP-binding proteins to activate MAP kinase. In animal models, CCK can promote pancreatic cancer growth.16
GRP (the mammalian analog of bombesin) was first recognized for its ability to stimulate gastrin secretion from the stomach. Neurons containing GRP are abundant in the oxyntic mucosal of the proximal stomach and, based on studies with a specific GRP antagonist, appear to play a major role in the cephalic phase of gastric acid secretion.153 It was later appreciated that GRP stimulates proliferation of G cells. GRP has received considerable attention for its growth-promoting effects on small cell lung cancer, pancreatic cancer, and certain colon cancers.154
TASTE RECEPTORS
The GI tract contains specialized taste receptor cells that detect chemicals and toxins. These cells are best characterized in the tongue, where they are concentrated in taste buds. Taste receptor cells can detect nutrients such as proteins, fats, sugars, and salt at submolar concentrations and other chemicals such as drugs and toxins at submicromolar concentrations.155 Detection of these chemical signals is important for several reasons. Distinguishing among chemicals can warn of dangerous chemical ingestants and induce a vomiting reflex, thus protecting the organism against poisoning. Alternatively, sensing various foods through taste can be pleasant and encourage food intake as well as facilitate digestion by stimulating salivary, gastric, and pancreatic secretions. Taste receptor cells contain a wide number of receptors and ion channels that serve as targets for different tastants.156 The membrane proteins can respond to molecules as simple as ions or as complex as fatty acids and proteins. Ions and amino acids activate taste receptor cells by directly interacting with ion channels. Such interactions often cause membrane depolarization and induce Ca2+ entry into the cell. Other types of molecules such as sweet and bitter tastants activate G protein–coupled receptors and stimulate production of second messengers, such as cAMP or inositol trisphosphate, which stimulate intracellular signaling processes.
A large number of taste receptors have been identified. A family of receptors that detects sweet compounds, including certain l amino acids, are known as T1R1, 2, and 3. A larger family, known as T2Rs, consisting of over 30 members, mediates bitter gustatory signals. The T1R and T2R families of receptors are G protein–coupled receptors that couple to α subunits, including α-gustducin and α-transducin. Interestingly, although taste receptors are abundant in the tongue, T2Rs, α-gustducin, and α-transducin have been identified in the gastric and intestinal mucosa and in the pancreas. In the intestine, T2Rs have been localized in some enteroendocrine and brush border cells. It is possible that chemical messengers stimulate T2Rs in these cells to secrete hormones or induce cellular responses, such as production of nitric oxide.157
INTRALUMINAL RELEASING FACTOR REGULATION OF GASTROINTESTINAL HORMONES
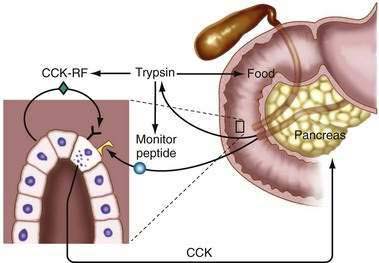
Most GI hormones are secreted into the blood following the ingestion of a meal. However, the exact mechanism whereby luminal nutrients stimulate hormone secretion is unknown. Although the apical surface of most enteric endocrine cells is exposed to the intestinal lumen (“open cells”), it is unclear whether nutrients interact with specific receptors on the surface of endocrine cells or whether they are absorbed and then stimulate hormone secretion. It has been recognized that specific releasing factors for GI hormones are present in the lumen of the gut (Fig. 1-10). CCK was the first hormone shown to be regulated by an intraluminal releasing factor.158,159 Luminal CCK-releasing factor was purified from intestinal washings and shown to stimulate CCK release when instilled into the lumen of animals. Diazepam-binding inhibitor has also been shown to stimulate CCK release, as has a pancreatic peptide known as monitor peptide.160,161 Secretin may also be regulated by an intraluminal releasing factor.28 The existence of these releasing factors underscores the significance of bioactive peptides within the lumen of the gut.
GASTROINTESTINAL PEPTIDES THAT REGULATE SATIETY AND HUNGER
During a meal, ingested nutrients interact with cells of the mouth and GI tract. Endocrine cells of the stomach and small intestine possess receptors that are linked to the secretion of GI hormones. GI peptides (see Chapters 6 and 8) are then released into the surrounding space, where they exert paracrine actions or are taken up into the circulation, where they function as hormones.162 Each of these transmitters facilitates the ingestion, digestion, absorption, or distribution of nutrients that are essential for the organism. Some GI hormones control the size of an ingested meal and are known as satiety signals. Satiety hormones share several qualities.163 First, they decrease meal size. Second, blocking their endogenous activity leads to increased meal size. Third, reduction of food intake is not the result of an aversion to food. Fourth, secretion of the hormone is caused by ingestion of food that normally causes cessation of eating (Table 1-3). Most satiety signals interact with specific receptors on nerves leading from the GI tract to the hindbrain.
Table 1-3 Gastrointestinal Peptides That Regulate Satiety and Food Intake
| REDUCE FOOD INTAKE | INCREASE FOOD INTAKE |
|---|---|
| Cholecystokinin (CCK) | Ghrelin |
| Glucagon-like peptide-1 | |
| Peptide tyrosine tyrosine (PYY3-36) | |
| Gastrin-releasing peptide | |
| Amylin | |
| Apolipoprotein A-IV | |
| Somatostatin |
CCK is one of the most extensively studied satiety hormones. In a time- and dose-dependent manner, CCK reduces food intake in animals and humans,164 an effect that is mediated by CCK-1 receptors residing on vagus nerve endings.165 The effect of CCK on food intake is a proven physiologic action because administration of a CCK receptor antagonist induces hunger and results in larger meal sizes. CCK also delays the rate at which food empties from the stomach, which may explain why the satiety actions of CCK are most apparent when the stomach is distended. Together, these findings indicate that CCK provides a signal for terminating a meal.
GLP-1 is produced by L cells of the ileum and colon and is released in response to food in the intestine. Although the primary action of GLP-1 is to stimulate insulin secretion, it also delays gastric emptying. Moreover, infusion of GLP-1 increases satiety and produces feelings of fullness, thereby reducing food intake without causing aversion.166 GLP-1 receptors are found in the periventricular nucleus, dorsal medial hypothalamus, and arcuate nucleus of the hypothalamus, which are important areas in the regulation of hunger. Like CCK, central administration of GLP-1 suppresses food intake.
PYY is also produced by L cells of the ileum and colon. Two forms of PYY are released into the circulation, PYY1-36 and PYY3-36. PYY1-36 binds to all subtypes of the neuropeptide Y family of receptors, whereas PYY3-36 has strong affinity for the Y2 receptor. When administered to animals, PYY3-36 causes a reduction in food intake, and mice lacking the Y2 receptor are resistant to the anorexigenic effects of PYY3-36, indicating that PYY3-36 signals satiety through this receptor.167 PYY3-36 has been shown in humans to decrease hunger scores and caloric intake.168 Interestingly, most of the GI peptide receptors involved in satiety are also found in the brain, where they mediate similar satiety effects. This may represent conservation of peptide signals that serve similar purposes.
Leptin is referred to as an adiposity signal because it is released into the blood in proportion to the amount of body fat and is considered a long-term regulator of energy balance. Together with CCK, leptin reduces food intake and produces a greater reduction in body weight than either agent alone.86 Therefore, it appears that long-term regulators of energy balance can affect short-term regulators through a decrease in meal size, which may promote weight reduction.
Hunger and initiation of a meal are intimately related. Ghrelin is intriguing because it is the only known circulating GI hormone that has orexigenic effects.102 Produced by the stomach, ghrelin levels increase abruptly before the onset of a meal and decrease rapidly after eating, suggesting that it signals initiation of a meal. Consistent with this role are studies demonstrating that administration of antighrelin antibodies or a ghrelin receptor antagonist suppresses food intake.169 It is not known if ghrelin is responsible for the hunger pains and audible bowel sounds that occur in people who are hungry.
Bariatric surgery, in particular Roux-en-Y gastric bypass, is the most effective procedure for long-term weight loss in morbid obesity. Although it had been assumed that weight loss accompanying this procedure was the result of reduced gastric capacity and calorie malabsorption, recent evidence of reduced ghrelin release and exaggerated PYY release after a meal has suggested that hormonal factors may contribute to reduced calorie intake.170
ENTEROINSULAR AXIS
GI hormones play an important role in the regulation of insulin secretion and glucose homeostasis. These hormones control processes that facilitate the digestion and absorption of nutrients, as well as disposal of nutrients that have reached the bloodstream. In particular, gut peptides control postprandial glucose levels through three different mechanisms: (1) stimulation of insulin secretion from pancreatic beta cells; (2) inhibition of hepatic gluconeogenesis by suppression of glucagon secretion; and (3) delaying the delivery of carbohydrates to the small intestine by inhibiting gastric emptying.171 Each of these actions reduces the blood glucose excursions that normally occur after eating.
Approximately 50% of the insulin released after a meal is the result of GI hormones that potentiate insulin secretion.172 This interaction is known as the enteroinsular axis and the gut peptides that stimulate insulin release are known as incretins. The major incretins are GLP-1 and GIP. GLP-1 not only stimulates insulin secretion but also increases beta cell mass, inhibits glucagon secretion, and delays gastric emptying. GIP stimulates insulin secretion when glucose levels are elevated and decreases glucagon-stimulated hepatic glucose production.173 Thus, on ingestion of a meal, glucose, as it is absorbed, stimulates GLP-1 and GIP secretion. Circulating glucose then stimulates beta cell production of insulin, and this effect is substantially augmented by incretins acting in conjunction with glucose to increase insulin levels.
Postprandial hyperglycemia may also be controlled by delaying the delivery of food from the stomach to the small intestine, allowing the rise in insulin to keep pace with the rate of glucose absorption. Several gut hormones that delay gastric emptying have been shown to reduce postprandial glucose excursions (Table 1-4).171 Amylin (islet amyloid polypeptide) is a 37–amino acid peptide synthesized primarily in the beta cells of the pancreatic islets together with insulin. Although it was originally recognized for its ability to form amyloid deposits in association with beta cell loss, it has more recently been found to suppress glucagon secretion, delay gastric emptying, and induce satiety.174 Insulin resistance in obese patients is associated with increased levels of both insulin and amylin.
Table 1-4 Gastrointestinal Peptides That Regulate Postprandial Blood Glucose Levels
Type II diabetes mellitus is characterized by high circulating insulin levels and insulin resistance. In addition, insulin levels do not increase appropriately after a meal and significant hyperglycemia occurs, which is consistent with an impaired incretin effect. GIP secretion is preserved in type II diabetes; however, the insulinotropic effect of GIP is reduced.175 Although the precise cause is unknown, the defect in GIP-stimulated insulin release is most pronounced in the late phase of insulin secretion. In contrast to GIP, GLP-1 secretion has been shown to be reduced in insulin-resistant type II diabetics. The lower GLP-1 levels are caused by impaired secretion rather than increased degradation of the hormone.176 Unlike GIP, the insulin response to infusion of GLP-1 is preserved, indicating that the beta cell can respond normally to this incretin hormone. These observations suggest that GLP-1 administration could be a viable treatment for the hyperglycemia associated with diabetes.177 The growing evidence that beta cell failure may develop in type II diabetes supports the use of incretin hormones, such as GLP-1, or agents that delay GLP-1 degradation by the enzyme dipeptidyl peptidase-4 (DPP-4) to enhance beta cell function.178,179
Batterham RL, Cowley MA, Small CJ, et al. Gut hormone PYY(3-36) physiologically inhibits food intake. Nature. 2002;418:650-4. (Ref 60.)
Boehning D, Snyder SH. Novel neural modulators. Annu Rev Neurosci. 2003;26:105-31. (Ref 119.)
Burdick JS, Chung E, Tanner G, et al. Treatment of Ménétrier’s disease with a monoclonal antibody against the epidermal growth factor receptor. N Engl J Med. 2000;343:1697-701. (Ref 142.)
Cummings DE, Weigle DS, Frayo RS, et al. Plasma ghrelin levels after diet-induced weight loss or gastric bypass surgery. N Engl J Med. 2002;346:1623-30. (Ref 100.)
de Herder WW, Lamberts SW. Somatostatin and somatostatin analogues: Diagnostic and therapeutic uses. Curr Opin Oncol. 2002;14:53-7. (Ref 75.)
Drucker DJ. Enhancing incretin action for the treatment of type 2 diabetes. Diabetes Care. 2003;26:2929-40. (Ref 175.)
Joseph IM, Zavros Y, Merchant JL, Kirschner D. A model for integrative study of human gastric acid secretion. J Appl Physiol. 2003;94:1602-18. (Ref 70.)
Miyawaki K, Yamada Y, Ban N, et al. Inhibition of gastric inhibitory polypeptide signaling prevents obesity. Nat Med. 2002;8:738-42. (Ref 48.)
Nakazato M, Murakami N, Date Y, et al. A role for ghrelin in the central regulation of feeding. Nature. 2001;409:194-8. (Ref 94.)
Pennefather JN, Lecci A, Candenas ML, et al. Tachykinins and tachykinin receptors: a growing family. Life Sci. 2004;74:1445-63. (Ref 65.)
Schlessinger J. Cell signaling by receptor tyrosine kinases. Cell. 2000;103:211-25. (Ref 133.)
Thomas RP, Hellmich MR, Townsend CMJr, Evers BM. Role of GI hormones in the proliferation of normal and neoplastic tissues. Endocr Rev. 2003;24:571-99. (Ref 73.)
Tracey KJ. The inflammatory reflex. Nature. 2002;420:853-9. (Ref 123.)
Vilsboll T, Zdravkovic M, Le-Thi T, et al. Liraglutide, a long-acting human glucagon-like peptide-1 analog, given as monotherapy significantly improves glycemic control and lowers body weight without risk of hypoglycemia in patients with type 2 diabetes. Diabetes Care. 2007;30:1608-10. (Ref 44.)
Woods SC. GI satiety signals I. An overview of GI signals that influence food intake. Am J Physiol Gastrointest Liver Physiol. 2004;286:G7-13. (Ref 163.)
1. Del Valle J, Yamada T. The gut as an endocrine organ. Annu Rev Med. 1990;41:447-55.
2. Larsson LI, Goltermann N, de Magistris L, et al. Somatostatin cell processes as pathways for paracrine secretion. Science. 1979;205:1393-5.
3. Grabowska AM, Watson SA. Role of gastrin peptides in carcinogenesis. Cancer Lett. 2007;257:1-15.
4. Gershon VMD. Genes, lineages, and tissue interactions in the development of the enteric nervous system. Am J Physiol. 1998;275(Pt 1):G869-73.
5. Dockray GJ. Physiology of enteric neuropeptides. In: Johnson LR, editor. Physiology of the GI tract. New York: Raven Press; 1994:129.
6. Murthy KS, Grider JR, Jin JG, Makhlouf GM. Interplay of VIP and nitric oxide in the regulation of neuromuscular activity in the gut. Arch Int Pharmacodyn Ther. 1995;329:27-38.
7. Furness JB, Clerc N. Responses of afferent neurons to the contents of the digestive tract, and their relation to endocrine and immune responses. Prog Brain Res. 2000;122:159-72.
8. Smith GP, Gibbs J. Satiating effect of cholecystokinin. Ann N Y Acad Sci. 1994;713:236-41.
9. Corsi AK, Schekman R. Mechanism of polypeptide translocation into the endoplasmic reticulum. J Biol Chem. 1996;271:30299-302.
10. Rehfeld JF. Processing of precursors of gastroenteropancreatic hormones: Diagnostic significance. J Mol Med. 1998;76:338-45.
11. Velander WH, Lubon H, Drohan WN. Transgenic livestock as drug factories. Sci Am. 1997;276:70-4.
12. Crooke ST. Progress in antisense technology. Annu Rev Med. 2004;55:61-95.
13. Kim PS, Arvan P. Endocrinopathies in the family of endoplasmic reticulum (ER) storage diseases: Disorders of protein trafficking and the role of ER molecular chaperones. Endocr Rev. 1998;19:173-202.
14. Liu X, Jiang Q, Mansfield SG, et al. Partial correction of endogenous DeltaF508 CFTR in human cystic fibrosis airway epithelia by spliceosome-mediated RNA trans-splicing. Nat Biotechnol. 2002;20:47-52.
15. Joshi SN, Gardner JD. Gastrin and colon cancer: A unifying hypothesis. Dig Dis. 1996;14:334-44.
16. Aly A, Shulkes A, Baldwin GS. Gastrins, cholecystokinins and GI cancer. Biochim Biophys Acta. 2004;1704:1-10.
17. Rehfeld JF. The new biology of GI hormones. Physiol Rev. 1998;78:1087-108.
18. Kopin AS, Lee YM, McBride EW, et al. Expression cloning and characterization of the canine parietal cell gastrin receptor. Proc Natl Acad Sci U S A. 1992;89:3605-9.
19. Liddle RA. Cholecystokinin. In: Walsh JH, Dockray GJ, editors. Gut peptides: Biochemistry and physiology. New York: Raven Press; 1994:175.
20. Reeve JRJr, Eysselein VE, Ho FJ, et al. Natural and synthetic CCK-58. Novel reagents for studying cholecystokinin physiology. Ann N Y Acad Sci. 1994;713:11-21.
21. Liddle RA, Morita ET, Conrad CK, Williams JA. Regulation of gastric emptying in humans by cholecystokinin. J Clin Invest. 1986;77:992-6.
22. Schmitz F, Göke MN, Otte JM, et al. Cellular expression of CCK-A and CCK-B/gastrin receptors in human gastric mucosa. Regul Pept. 2001;102:101-10.
23. Calam J, Ellis A, Dockray GJ. Identification and measurement of molecular variants of cholecystokinin in duodenal mucosa and plasma. Diminished concentrations in patients with celiac disease. J Clin Invest. 1982;69:218-25.
24. Geracioti TDJr, Liddle RA. Impaired cholecystokinin secretion in bulimia nervosa. N Engl J Med. 1988;319:683-8.
25. Slaff JI, Wolfe MM, Toskes PP. Elevated fasting cholecystokinin levels in pancreatic exocrine impairment: Evidence to support feedback regulation. J Lab Clin Med. 1985;105:282-5.
26. Bayliss WM, Starling EH. The mechanism of pancreatic secretion. J Physiol. 1902;28:325-53.
27. Leiter AB, Chey WY, Kopin AS. Secretin. In: Walsh JH, Dockray GJ, editors. Gut peptides: Biochemistry and physiology. New York: Raven Press; 1994:144.
28. Li P, Lee KY, Chang TM, Chey WY. Mechanism of acid-induced release of secretin in rats. Presence of a secretin-releasing peptide. J Clin Invest. 1990;86:1474-9.
29. You CH, Chey WY. Secretin is an enterogastrone in humans. Dig Dis Sc. 1987;32:466-71.
30. McGuigan JE, Wolfe MM. Secretin injection test in the diagnosis of gastrinoma. Gastroenterology. 1980;79:1324-31.
31. Said SI, Mutt V. Polypeptide with broad biological activity: Isolation from small intestine. Science. 1970;169:1217-18.
32. Dockray GJ. Vasoactive intestinal polypeptide and related peptides. In: Walsh JH, Dockray GJ, editors. Gut peptides: Biochemistry and physiology. New York: Raven Press; 1994:447.
33. Holst JJ, Fahrenkrug J, Knuhtsen S, et al. VIP and PHI in the pig pancreas: Coexistence, corelease, and cooperative effects. Am J Physiol. 1987;252(Pt 1):G182-9.
34. Fahrenkrug J. Transmitter role of vasoactive intestinal peptide. Pharmacol Toxicol. 1993;72:354-63.
35. Bloom SR. Vasoactive intestinal peptide, the major mediator of the WDHA (pancreatic cholera) syndrome: Value of measurement in diagnosis and treatment. Am J Dig Dis. 1978;23:373-6.
36. Cho WK, Boyer JL. Vasoactive intestinal polypeptide is a potent regulator of bile secretion from rat cholangiocytes. Gastroenterology. 1999;117:420-8.
37. Grider JR. Interplay of VIP and nitric oxide in regulation of the descending relaxation phase of peristalsis. Am J Physiol. 1993;264(Pt 1):G334-40.
38. Aggestrup S, Uddman R, Sundler F, et al. Lack of vasoactive intestinal polypeptide nerves in esophageal achalasia. Gastroenterology. 1983;84(Pt 1):924-7.
39. Larsson LT, Sundler F. Is the reduction of VIP the clue to the pathophysiology of Hirschsprung’s disease? Z Kinderchir. 1990;45:164-6.
40. Verner JV, Morrison AB. Endocrine pancreatic islet disease with diarrhea. Report of a case due to diffuse hyperplasia of nonbeta islet tissue with a review of 54 additional cases. Arch Intern Med. 1974;133:492-9.
41. Smith SL, Branton SA, Avino AJ, et al. Vasoactive intestinal polypeptide secreting islet cell tumors: A 15-year experience and review of the literature. Surgery. 1998;124:1050-5.
42. Mojsov S, Heinrich G, Wilson IB, et al. Preproglucagon gene expression in pancreas and intestine diversifies at the level of post-translational processing. J Biol Chem. 1986;261:11880-9.
43. Drucker DJ. Glucagon-like peptides. Diabetes. 1998;47:159-69.
44. Vilsboll T, Zdravkovic M, Le-Thi T, et al. Liraglutide, a long-acting human glucagon-like peptide-1 analog, given as monotherapy significantly improves glycemic control and lowers body weight without risk of hypoglycemia in patients with type 2 diabetes. Diabetes Care. 2007;30:1608-10.
45. Vilsboll T, Brock B, Perrild H, et al. Liraglutide, a once-daily human GLP-1 analogue, improves pancreatic B-cell function and arginine-stimulated insulin secretion during hyperglycaemia in patients with type 2 diabetes mellitus. Diabet Med. 2008;25:152-6.
46. Pederson RA. Gastric inhibitory polypeptide. In: Walsh JH, Dockray GJ, editors. Gut peptides: Biochemistry and physiology. New York: Raven Press; 1994:217.
47. Kieffer TJ. GIP or not GIP? That is the question. Trends Pharmacol Sci. 2003;24:110-12.
48. Miyawaki K, Yamada Y, Ban N, et al. Inhibition of gastric inhibitory polypeptide signaling prevents obesity. Nat Med. 2002;8:738-42.
49. Lacroix A, Bolte E, Tremblay J, et al. Gastric inhibitory polypeptide-dependent cortisol hypersecretion—a new cause of Cushing’s syndrome. N Engl J Med. 1992;327:974-80.
50. Reznik Y, Allali-Zerah V, Chayvialle JA, et al. Food-dependent Cushing’s syndrome mediated by aberrant adrenal sensitivity to gastric inhibitory polypeptide. N Engl J Med. 1992;327:981-6.
51. Kimmel JR, Hayden LJ, Pollock HG. Isolation and characterization of a new pancreatic polypeptide hormone. J Biol Chem. 1975;250:9369-76.
52. Tatemoto K, Mutt V. Chemical determination of polypeptide hormones. Proc Natl Acad Sci U S A. 1978;75:4115-19.
53. Tatemoto K, Carlquist M, Mutt V. Neuropeptide Y—a novel brain peptide with structural similarities to peptide YY and pancreatic polypeptide. Nature. 1982;296:659-60.
54. Larsson LI, Sundler F, Hakanson R. Pancreatic polypeptide—a postulated new hormone: Identification of its cellular storage site by light and electron microscopic immunocytochemistry. Diabetologia. 1976;12:211-26.
55. Wahlestedt C, Reis DJ. Neuropeptide Y-related peptides and their receptors—are the receptors potential therapeutic drug targets? Annu Rev Pharmacol Toxicol. 1993;33:309-52.
56. Lundberg JM, Tatemoto K, Terenius L, et al. Localization of peptide YY (PYY) in GI endocrine cells and effects on intestinal blood flow and motility. Proc Natl Acad Sci U S A. 1982;79:4471-5.
57. Gehlert DR. Multiple receptors for the pancreatic polypeptide (PP-fold) family: Physiological implications. Proc Soc Exp Biol Med. 1998;218:7-22.
58. Mannon P, Taylor IL. The pancreatic polypeptide family. In: Walsh JH, Dockray GJ, editors. Gut peptides: Biochemistry and physiology. New York: Raven Press; 1994:341.
59. Lloyd KC, Grandt D, Aurang K, et al. Inhibitory effect of PYY on vagally stimulated acid secretion is mediated predominantly by Y1 receptors. Am J Physiol. 1996;270(Pt 1):G123-7.
60. Batterham RL, Cowley MA, Small CJ, et al. Gut hormone PYY(3-36) physiologically inhibits food intake. Nature. 2002;418:650-4.
61. Hwa JJ, Witten MB, Williams P, et al. Activation of the NPY Y5 receptor regulates both feeding and energy expenditure. Am J Physiol. 1999;277(Pt 2):R1428-34.
62. Hazelwood RL. The pancreatic polypeptide (PP-fold) family: Gastrointestinal, vascular, and feeding behavioral implications. Proc Soc Exp Biol Med. 1993;202:44-63.
63. Cao T, Pinter E, Al-Rashed S, et al. Neurokinin-1 receptor agonists are involved in mediating neutrophil accumulation in the inflamed, but not normal, cutaneous microvasculature: An in vivo study using neurokinin-1 receptor knockout mice. J Immunol. 2000;164:5424-9.
64. Mantyh PW, DeMaster E, Malhotra A, et al. Receptor endocytosis and dendrite reshaping in spinal neurons after somatosensory stimulation. Science. 1995;268:1629-32.
65. Pennefather JN, Lecci A, Candenas ML, et al. Tachykinins and tachykinin receptors: A growing family. Life Sci. 2004;74:1445-63.
66. Mantyh CR, Pappas TN, Lapp JA, et al. Substance P activation of enteric neurons in response to intraluminal Clostridium difficile toxin A in the rat ileum. Gastroenterology. 1996;111:1272-80.
67. Mantyh PW, Mantyh CR, Gates T, et al. Receptor binding sites for substance P and substance K in the canine gastrointestinal tract and their possible role in inflammatory bowel disease. NeuroScience. 1988;25:817-37.
68. Chiba T, Yamada T. Gut somatostatin. In: Walsh JH, Dockray GJ, editors. Gut peptides: Biochemistry and physiology. New York: Raven Press; 1994:123.
69. Kolivas S, Shulkes A. Regulation of expression of the receptors controlling gastric acidity. Regul Pept. 2004;121:1-9.
70. Joseph IM, Zavros Y, Merchant JL, Kirschner D. A model for integrative study of human gastric acid secretion. J Appl Physiol. 2003;94:1602-18.
71. Patel YC, Greenwood MT, Panetta R, et al. The somatostatin receptor family. Life Sci. 1995;57:1249-65.
72. Chey WY, Chang T. Neural hormonal regulation of exocrine pancreatic secretion. Pancreatology. 2001;1:320-35.
73. Thomas RP, Hellmich MR, Townsend CMJr, Evers BM. Role of gastrointestinal hormones in the proliferation of normal and neoplastic tissues. Endocr Rev. 2003;24:571-99.
74. Florio T. Molecular mechanisms of the antiproliferative activity of somatostatin receptors (SSTRs) in neuroendocrine tumors. Front Biosci. 2008;13:822-40.
75. de Herder WW, Lamberts SW. Somatostatin and somatostatin analogues: Diagnostic and therapeutic uses. Curr Opin Oncol. 2002;14:53-7.
76. de Franchis R. Somatostatin, somatostatin analogues and other vasoactive drugs in the treatment of bleeding oesophageal varices. Dig Liver Dis. 2004;36(Suppl 1):S93-100.
77. Poitras P. Motilin. In: Walsh JH, Dockray GJ, editors. Gut peptides: Biochemistry and physiology. New York: Raven Press; 1994:261.
78. Depoortere I. Motilin and motilin receptors: characterization and functional significance. Verh K Acad Geneeskd Belg. 2001;63:511-29.
79. Pilot MA. Macrolides in roles beyond antibiotic therapy. Br J Surg. 1994;81:1423-9.
80. Itoh Z. Motilin and clinical application. Peptides. 1997;18:593-608.
81. Zhang Y, Proenca R, Maffei M, et al. Positional cloning of the mouse obese gene and its human homologue. Nature. 1994;372:425-32.
82. Tartaglia LA, Dembski M, Weng X, et al. Identification and expression cloning of a leptin receptor, OB-R. Cell. 1995;83:1263-71.
83. Vaisse C, Halaas JL, Horvath CM, et al. Leptin activation of Stat3 in the hypothalamus of wild-type and ob/ob mice but not db/db mice. Nat Genet. 1996;14:95-7.
84. Banks WA, Kastin AJ, Huang W, et al. Leptin enters the brain by a saturable system independent of insulin. Peptides. 1996;17:305-11.
85. Schwartz MW, Seeley RJ, Campfield LA, et al. Identification of targets of leptin action in rat hypothalamus. J Clin Invest. 1996;98:1101-6.
86. Lewin MJ, Bado A. Gastric leptin. Microsc Res Tech. 2001;53:372-6.
87. Morton GJ, Blevins JE, Williams DL, et al. Leptin action in the forebrain regulates the hindbrain response to satiety signals. J Clin Invest. 2005;115:703-10.
88. Ostlund REJr, Yang JW, Klein S, Gingerich R. Relation between plasma leptin concentration and body fat, gender, diet, age, and metabolic covariates. J Clin Endocrinol Metab. 1996;81:3909-13.
89. Montague CT, Farooqi IS, Whitehead JP, et al. Congenital leptin deficiency is associated with severe early-onset obesity in humans. Nature. 1997;387:903-8.
90. Strobel A, Issad T, Camoin L, et al. A leptin missense mutation associated with hypogonadism and morbid obesity. Nat Genet. 1998;18:213-15.
91. Clement K, Vaisse C, Lahlou N, et al. A mutation in the human leptin receptor gene causes obesity and pituitary dysfunction. Nature. 1998;392:398-401.
92. Kojima M, Hosoda H, Date Y, et al. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature. 1999;402:656-60.
93. Takaya K, Ariyasu H, Kanamoto N, et al. Ghrelin strongly stimulates growth hormone release in humans. J Clin Endocrinol Metab. 2000;85:4908-11.
94. Nakazato M, Murakami N, Date Y, et al. A role for ghrelin in the central regulation of feeding. Nature. 2001;409:194-8.
95. Ariyasu H, Takaya K, Tagami T, et al. Stomach is a major source of circulating ghrelin, and feeding state determines plasma ghrelin-like immunoreactivity levels in humans. J Clin Endocrinol Metab. 2001;86:4753-8.
96. Date Y, Kojima M, Hosoda H, et al. Ghrelin, a novel growth hormone-releasing acylated peptide, is synthesized in a distinct endocrine cell type in the gastrointestinal tracts of rats and humans. Endocrinology. 2000;141:4255-61.
97. Hosoda H, Kojima M, Kangawa K. Ghrelin and the regulation of food intake and energy balance. Mol Interv. 2002;2:494-503.
98. Tschop M, Weyer C, Tataranni PA, et al. Circulating ghrelin levels are decreased in human obesity. Diabetes. 2001;50:707-9.
99. Otto B, Cuntz U, Fruehauf E, et al. Weight gain decreases elevated plasma ghrelin concentrations of patients with anorexia nervosa. Eur J Endocrinol. 2001;145:669-73.
100. Cummings DE, Weigle DS, Frayo RS, et al. Plasma ghrelin levels after diet-induced weight loss or gastric bypass surgery. N Engl J Med. 2002;346:1623-30.
101. Kamegai J, Tamura H, Shimizu T, et al. Chronic central infusion of ghrelin increases hypothalamic neuropeptide Y and Agouti-related protein mRNA levels and body weight in rats. Diabetes. 2001;50:2438-43.
102. Inui A, Asakawa A, Bowers CY, et al. Ghrelin, appetite, and gastric motility: The emerging role of the stomach as an endocrine organ. FASEB J. 2004;18:439-56.
103. Cummings DE, Clement K, Purnell JQ, et al. Elevated plasma ghrelin levels in Prader Willi syndrome. Nat Med. 2002;8:643-4.
104. DelParigi A, Tschop M, Heiman ML, et al. High circulating ghrelin: A potential cause for hyperphagia and obesity in Prader-Willi syndrome. J Clin Endocrinol Metab. 2002;87:5461-4.
105. Wren AM, Seal LJ, Cohen MA, et al. Ghrelin enhances appetite and increases food intake in humans. J Clin Endocrinol Metab. 2001;86:5992.
106. Dajas-Bailador F, Wonnacott S. Nicotinic acetylcholine receptors and the regulation of neuronal signalling. Trends Pharmacol Sci. 2004;25:317-24.
107. Velasco M, Luchsinger A. Dopamine: Pharmacologic and therapeutic aspects. Am J Ther. 1998;5:37-43.
108. Missale C, Nash SR, Robinson SW, et al. Dopamine receptors: From structure to function. Physiol Rev. 1998;78:189-225.
109. Gershon MD. Review article: Roles played by 5-hydroxytryptamine in the physiology of the bowel. Aliment Pharmacol Ther. 1999;13(Suppl 2):15-30.
110. Kim DY, Camilleri M. Serotonin: A mediator of the brain-gut connection. Am J Gastroenterol. 2000;95:2698-709.
111. Talley NJ. Serotoninergic neuroenteric modulators. Lancet. 2001;358:2061-8.
112. Pandolfino JE, Howden CW, Kahrilas PJ. Motility-modifying agents and management of disorders of gastrointestinal motility. Gastroenterology. 2000;118(Suppl 1):S32-47.
113. Camilleri M. Management of the irritable bowel syndrome. Gastroenterology. 2001;120:652-68.
114. Bouras EP, Camilleri M, Burton DD, et al. Prucalopride accelerates gastrointestinal and colonic transit in patients with constipation without a rectal evacuation disorder. Gastroenterology. 2001;120:354-60.
115. Zheng W, Cole PA. Serotonin N-acetyltransferase: Mechanism and inhibition. Curr Med Chem. 2002;9:1187-99.
116. Bubenik GA. Localization, physiological significance and possible clinical implication of gastrointestinal melatonin. Biol Signals Recept. 2001;10:350-66.
117. Bazar KA, Yun AJ, Lee PY. Debunking a myth: Neurohormonal and vagal modulation of sleep centers, not redistribution of blood flow, may account for postprandial somnolence. Med Hypotheses. 2004;63:778-82.
118. Hill SJ, Ganellin CR, Timmerman H, et al. International Union of Pharmacology. XIII. Classification of histamine receptors. Pharmacol Rev. 1997;49:253-78.
119. Boehning D, Snyder SH. Novel neural modulators. Annu Rev Neurosci. 2003;26:105-31.
120. Bornstein JC, Costa M, Grider JR. Enteric motor and interneuronal circuits controlling motility. Neurogastroenterol Motil. 2004;16(Suppl 1):34-8.
121. Wallace JL, Miller MJ. Nitric oxide in mucosal defense: A little goes a long way. Gastroenterology. 2000;119:512-20.
122. Schulte G, Fredholm BB. Signalling from adenosine receptors to mitogen-activated protein kinases. Cell Signal. 2003;15:813-27.
123. Tracey KJ. The inflammatory reflex. Nature. 2002;420:853-9.
124. Lefkowitz RJ. The superfamily of heptahelical receptors. Nat Cell Biol. 2000;2:E133-6.
125. Palczewski K, Kumasaka T, Hori T, et al. Crystal structure of rhodopsin: A G protein–coupled receptor. Science. 2000;289:739-45.
126. Patel TB. Single transmembrane spanning heterotrimeric G protein–coupled receptors and their signaling cascades. Pharmacol Rev. 2004;56:371-85.
127. Pierce KL, Premont RT, Lefkowitz RJ. Seven-transmembrane receptors. Nat Rev Mol Cell Biol. 2002;3:639-50.
128. Simonds WF. G protein regulation of adenylate cyclase. Trends Pharmacol Sci. 1999;20:66-73.
129. Robishaw JD, Berlot CH. Translating G protein subunit diversity into functional specificity. Curr Opin Cell Biol. 2004;16:206-9.
130. Clapham DE, Neer EJ. G protein beta gamma subunits. Annu Rev Pharmacol Toxicol. 1997;37:167-203.
131. Litosch I. Novel mechanisms for feedback regulation of phospholipase C-beta activity. IUBMB Life. 2002;54:253-60.
132. Kohout TA, Lefkowitz RJ. Regulation of G protein–coupled receptor kinases and arrestins during receptor desensitization. Mol Pharmacol. 2003;63:9-18.
133. Schlessinger J. Cell signaling by receptor tyrosine kinases. Cell. 2000;103:211-25.
134. Simon MA. Receptor tyrosine kinases: Specific outcomes from general signals. Cell. 2000;103:13-15.
135. Pawson T. Specificity in signal transduction: From phosphotyrosine-SH2 domain interactions to complex cellular systems. Cell. 2004;116:191-203.
136. Kuhn M. Structure, regulation, and function of mammalian membrane guanylyl cyclase receptors, with a focus on guanylyl cyclase-A. Circ Res. 2003;93:700-9.
137. Yeatman TJ. A renaissance for SRC. Nat Rev Cancer. 2004;4:470-80.
138. Irie-Sasaki J, Sasaki T, Penninger JM. CD45-regulated signaling pathways. Curr Top Med Chem. 2003;3:783-96.
139. Shi Y, Massague J. Mechanisms of TGF-beta signaling from cell membrane to the nucleus. Cell. 2003;113:685-700.
140. Podolsky DK. Peptide growth factors in the gastrointestinal tract. In: Johnson LR, editor. Physiology of the gastrointestinal tract. New York: Raven Press; 1994:129.
141. Zimmerman CM, Padgett RW. Transforming growth factor beta signaling mediators and modulators. Gene. 2000;249:17-30.
142. Burdick JS, Chung E, Tanner G, et al. Treatment of Ménétrier’s disease with a monoclonal antibody against the epidermal growth factor receptor. N Engl J Med. 2000;343:1697-701.
143. Settle SH, Washington K, Lind C, et al. Chronic treatment of Ménétrier’s disease with Erbitux: Clinical efficacy and insight into pathophysiology. Clin Gastroenterol Hepatol. 2005;3:654-9.
144. Caraglia M, Marra M, Meo G, Addeo SR, et al. EGF-R small inhibitors and anti-EGF-R antibodies: Advantages and limits of a new avenue in anticancer therapy. Recent. Patents Anticancer Drug Discov. 2006;1:209-22.
145. Massague J. How cells read TGF-beta signals. Nat Rev Mol Cell Biol. 2000;1:169-78.
146. Williams RS, Rossi AM, Chegini N, Schultz G. Effect of transforming growth factor beta on postoperative adhesion formation and intact peritoneum. J Surg Res. 1992;52:65-70.
147. Adams TE, Epa VC, Garrett TP, Ward CW. Structure and function of the type 1 insulin-like growth factor receptor. Cell Mol Life Sci. 2000;57:1050-93.
148. Ornitz DM. FGFs, heparan sulfate and FGFRs: Complex interactions essential for development. Bioessays. 2000;22:108-12.
149. Kim SK, MacDonald RJ. Signaling and transcriptional control of pancreatic organogenesis. Curr Opin Genet Dev. 2002;12:540-7.
150. Podolsky DK. Mechanisms of regulatory peptide action in the gastrointestinal tract: Trefoil peptides. J Gastroenterol. 2000;35(Suppl 12):69-74.
151. Koh TJ, Chen D. Gastrin as a growth factor in the gastrointestinal tract. Regul Pept. 2000;93:37-44.
152. Smith AM, Watson SA. Gastrin and gastrin receptor activation: An early event in the adenoma-carcinoma sequence. Gut. 2000;47:820-4.
153. Hildebrand P, Lehmann FS, Ketterer S, et al. Regulation of gastric function by endogenous gastrin-releasing peptide in humans: Studies with a specific gastrin-releasing peptide antagonist. Gut. 2001;49:23-8.
154. Schally AV, Szepeshazi K, Nagy A, et al. New approaches to therapy of cancers of the stomach, colon and pancreas based on peptide analogs. Cell Mol Life Sci. 2004;61:1042-68.
155. Sternini C. Taste receptors in the gastrointestinal tract. IV. Functional implications of bitter taste receptors in gastrointestinal chemosensing. Am J Physiol Gastrointest Liver Physiol. 2007;292:G457-61.
156. Rozengurt E, Sternini C. Taste receptor signaling in the mammalian gut. Curr Opin Pharmacol. 2007;7:557-62.
157. Hofer D, Asan E, Drenckhahn D. Chemosensory perception in the gut. News Physiol Sci. 1999;14:18-23.
158. Spannagel AW, Green GM, Guan D, et al. Purification and characterization of a luminal cholecystokinin-releasing factor from rat intestinal secretion. Proc Natl Acad Sci U S A. 1996;93:4415-20.
159. Herzig KH, Schon I, Tatemoto K, et al. Diazepam binding inhibitor is a potent cholecystokinin-releasing peptide in the intestine. Proc Natl Acad Sci U S A. 1996;93:7927-32.
160. Iwai K, Fukuoka S, Fushiki T, et al. Purification and sequencing of a trypsin-sensitive cholecystokinin-releasing peptide from rat pancreatic juice. Its homology with pancreatic secretory trypsin inhibitor. J Biol Chem. 1987;262:8956-9.
161. Li Y, Hao Y, Owyang C. Diazepam-binding inhibitor mediates feedback regulation of pancreatic secretion and postprandial release of cholecystokinin. J Clin Invest. 2000;105:351-9.
162. Cummings DE, Overduin J. Gastrointestinal regulation of food intake. J Clin Invest. 2007;117:13-23.
163. Woods SC. GI satiety signals I. An overview of GI signals that influence food intake. Am J Physiol Gastrointest Liver Physiol. 2004;286:G7-13.
164. Moran TH, Kinzig KP. Gastrointestinal satiety signals II. Cholecystokinin. Am J Physiol Gastrointest Liver Physiol. 2004;286:G183-8.
165. Ritter RC, Covasa M, Matson CA. Cholecystokinin: Proofs and prospects for involvement in control of food intake and body weight. Neuropeptides. 1999;33:387-99.
166. Stanley S, Wynne K, Bloom S. Gastrointestinal satiety signals III. Glucagon-like peptide 1, oxyntomodulin, peptide YY, and pancreatic polypeptide. Am J Physiol Gastrointest Liver Physiol. 2004;286:G693-7.
167. Sainsbury A, Schwarzer C, Couzens M, et al. Important role of hypothalamic Y2 receptors in body weight regulation revealed in conditional knockout mice. Proc Natl Acad Sci U S A. 2002;99:8938-43.
168. Batterham RL, Cohen MA, Ellis SM, et al. Inhibition of food intake in obese subjects by peptide YY3-36. N Engl J Med. 2003;349:941-8.
169. Korbonits M, Goldstone AP, Gueorguiev M, Grossman AB. Ghrelin—a hormone with multiple functions. Front Neuroendocrinol. 2004;25:27-68.
170. Korner J, Bessler M, Cirilo LJ, et al. Effects of Roux-en-Y gastric bypass surgery on fasting and postprandial concentrations of plasma ghrelin, peptide YY, and insulin. J Clin Endocrinol Metab. 2005;90:359-65.
171. Perfetti R, Brown TA, Velikina R, Busselen S. Control of glucose homeostasis by incretin hormones. Diabetes Technol Ther. 1999;1:297-305.
172. Fehmann HC, Gherzi R, Goke B. Regulation of islet hormone gene expression by incretin hormones. Exp Clin Endocrinol Diabetes. 1995;103(Suppl 2):56-65.
173. Fehmann HC, Goke R, Goke B. Cell and molecular biology of the incretin hormones glucagon-like peptide-I and glucose-dependent insulin-releasing polypeptide. Endocr Rev. 1995;16:390-410.
174. Reda TK, Geliebter A, Pi-Sunyer FX. Amylin, food intake, and obesity. Obes Res. 2002;10:1087-91.
175. Drucker DJ. Enhancing incretin action for the treatment of type 2 diabetes. Diabetes Care. 2003;26:2929-40.
176. Vilsboll T, Agerso H, Krarup T, Holst JJ. Similar elimination rates of glucagon-like peptide-1 in obese type 2 diabetic patients and healthy subjects. J Clin Endocrinol Metab. 2003;88:220-4.
177. Ahren B. Gut peptides and type 2 diabetes mellitus treatment. Curr Diab Rep. 2003;3:365-72.
178. Ranganath LR. Incretins: Pathophysiological and therapeutic implications of glucose-dependent insulinotropic polypeptide and glucagon-like peptide-1. J Clin Pathol. 2008;61:401-9.
179. Raz I, Hanefeld M, Xu L, et al. Efficacy and safety of the dipeptidyl peptidase-4 inhibitor sitaglyptin as monotheapy in patients with typer 2 diabetes mellitus. Diabetologia. 2006;49:2564-71.