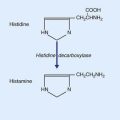
1 Embryology
This chapter aims to give you sufficient insight into development to account for the arrangement of structures in the mature nervous system. If not already familiar with adult brain anatomy, we suggest you read this chapter again following study of Chapters 2 and 3.
Spinal Cord
Neurulation
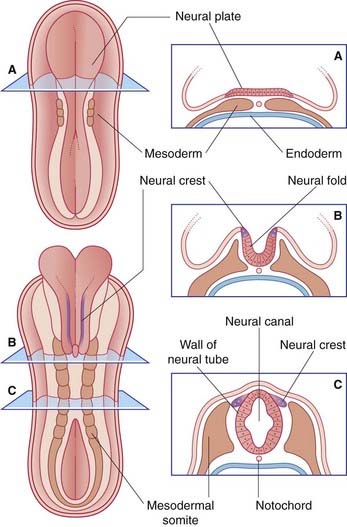
The entire nervous system originates from the neural plate, an ectodermal thickening in the floor of the amniotic sac (Figure 1.1). During the third week after fertilization, the plate forms paired neural folds, which unite to create the neural tube and neural canal. Union of the folds commences in the future neck region of the embryo and proceeds rostrally and caudally from there. The open ends of the tube, the neuropores, are closed off before the end of the fourth week. The process of formation of the neural tube from the ectoderm is known as neurulation.
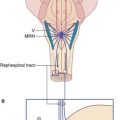
Spinal nerves
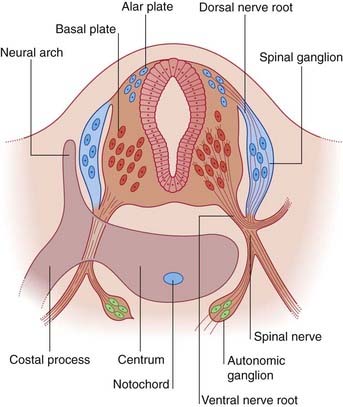
The dorsal part of the neural tube is called the alar plate; the ventral part is the basal plate (Figure 1.2). Neurons developing in the alar plate are predominantly sensory in function and receive dorsal nerve roots growing in from the spinal ganglia. Neurons in the basal plate are predominantly motor and give rise to ventral nerve roots. At appropriate levels of the spinal cord, the ventral roots also contain autonomic fibers. The dorsal and ventral roots unite to form the spinal nerves, which emerge from the vertebral canal in the interval between the neural arches being formed by the mesenchymal vertebrae.
Brain
Brain parts
Late in the fourth week, the rostral part of the neural tube undergoes flexion at the level of the future midbrain (Figure 1.3A). This region is the mesencephalon; slight constrictions mark its junction with the prosencephalon (future forebrain) and rhombencephalon (future hindbrain).
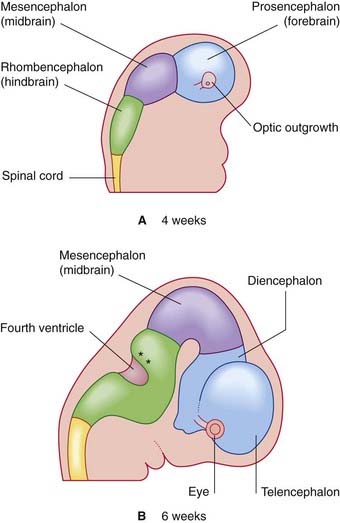
The alar plate of the prosencephalon expands on each side (Figure 1.3A) to form the telencephalon (cerebral hemispheres). The basal plate remains in place here as the diencephalon. Finally, an optic outgrowth from the diencephalon is the forerunner of the retina and optic nerve.
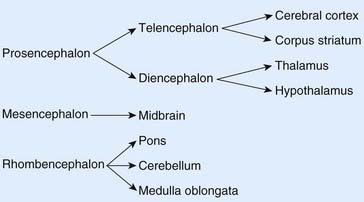
The diencephalon, mesencephalon, and rhombencephalon constitute the embryonic brainstem.
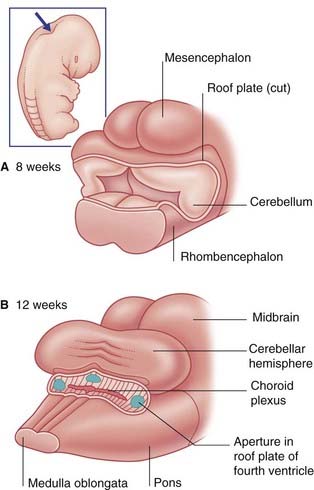
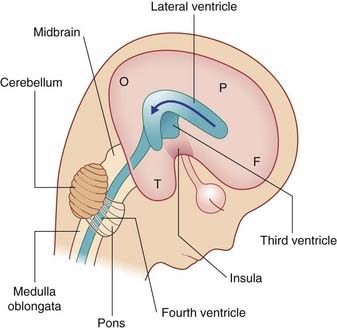
The brainstem buckles as development proceeds. As a result, the mesencephalon is carried to the summit of the brain. The rhombencephalon folds on itself, causing the alar plates to flare and creating the rhomboid (diamond-shaped) fourth ventricle of the brain. The rostral part of the rhombencephalon gives rise to the pons and cerebellum. The caudal part gives rise to the medulla oblongata (Figure 1.4).
Ventricular system and choroid plexuses
The neural canal dilates within the cerebral hemispheres, forming the lateral ventricles; these communicate with the third ventricle contained within the diencephalon. The third and fourth ventricles communicate through the aqueduct of the midbrain (Figure 1.5).
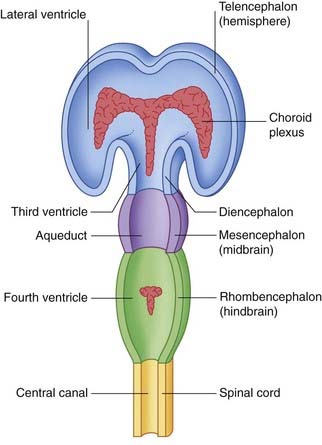
The thin roofs of the forebrain and hindbrain are invaginated by tufts of capillaries, which form the choroid plexuses of the four ventricles. The choroid plexuses secrete cerebrospinal fluid (CSF), which flows through the ventricular system. The fluid leaves the fourth ventricle through three apertures in its roof (Figure 1.6).
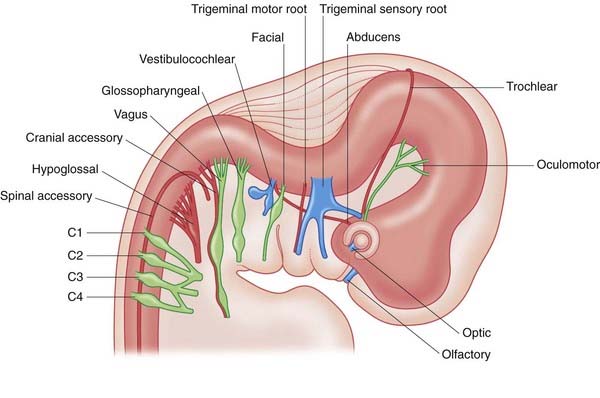
Cranial nerves
Figure 1.7 illustrates the state of development of the cranial nerves during the sixth week after fertilization.
Cerebral hemispheres
Expansion of the cerebral hemispheres is not uniform. A region on the lateral surface, the insula (L. ‘island’), is relatively quiescent and forms a pivot around which the expanding hemisphere rotates. Frontal, parietal, occipital, and temporal lobes can be identified at 14 weeks’ gestational age (Figure 1.8).
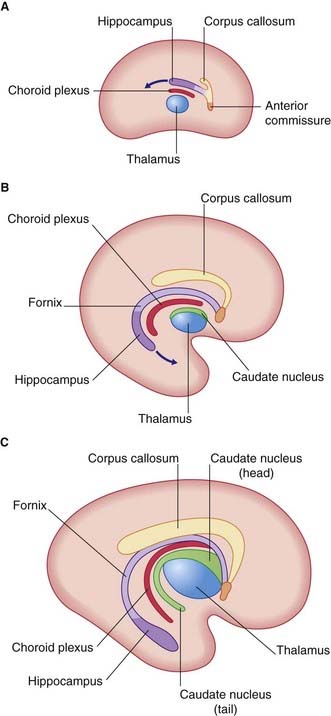
On the medial surface of the hemisphere, a patch of cerebral cortex, the hippocampus, belongs to a fifth, limbic lobe of the brain. The hippocampus is drawn into the temporal lobe, leaving in its wake a strand of fibers called the fornix. Within the concavity of this arc is the choroid fissure, through which the choroid plexus invaginates into the lateral ventricle (Figure 1.9).
Coronal sections of the telencephalon reveal a mass of gray matter in the base of each hemisphere, which is the forerunner of the corpus striatum. Beside the third ventricle, the diencephalon gives rise to the thalamus and hypothalamus (Figure 1.10).
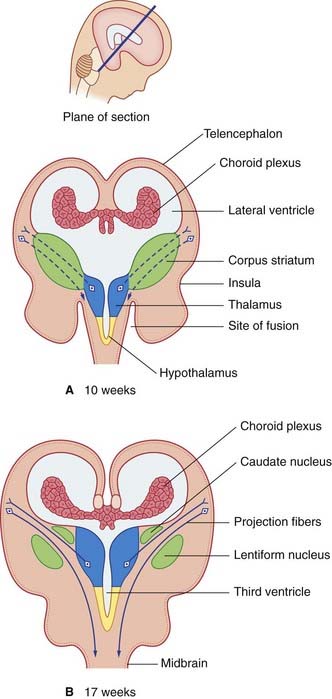
The expanding cerebral hemispheres come into contact with the diencephalon, and they fuse with it (see ‘site of fusion’ in Figure 1.10A). One consequence is that the term ‘brainstem’ is restricted thereafter to the remaining, free parts: midbrain, pons, and medulla oblongata. A second consequence is that the cerebral cortex is able to project fibers direct to the brainstem. Together with fibers projecting from thalamus to cortex, they split the corpus striatum into caudate and lentiform nuclei (Figure 1.10B).
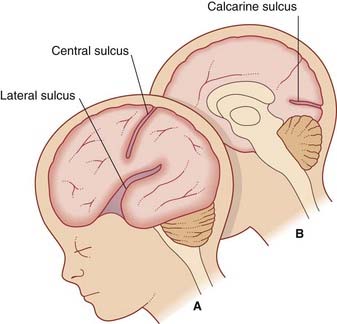
By the 28th week of development, several sulci (fissures) have appeared on the surface of the brain, notably the lateral, central, and calcarine sulci (Figure 1.11).
Bossy J, O’Rahilly R, Müller F. Ontogenese du systeme nerveux. In: Bossy J, editor. Anatomie clinique: neuroanatomie. Paris: Springer-Verlag; 1990:357-388.
Dubois J, Benders M, Borradori-Tolsa, et al. Primary cortical folding in the human newborn: an early marker of later functional development. Brain. 2008;131:2028-4041.
Le Douarin, Britto JM, Creuzer S. Role of the neural crest in face and brain development. Brain Res Dev. 2007;55:237-247.
O’Rahilly R, Muller F. Significant features in the early prenatal development of the human brain. Ann Anat. 2008;104:105-118.