[level-membership-for-basic-science-category]
23 Drives and Emotions: The Hypothalamus and Limbic System
The Hypothalamus Coordinates Drive-Related Behaviors
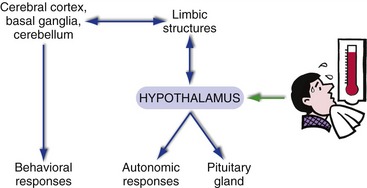
The hypothalamus is a nodal point in the neural circuits underlying drive-related behaviors (Fig. 23-1). It’s got interconnections with visceral parts of the nervous system, through which it is informed of and controls things like blood glucose, blood pressure, and body temperature. It’s also got interconnections with limbic structures, through which you become aware of homeostatic needs (“I’m hungry”). Finally, the hypothalamus has not just neural outputs but also ways to control the pituitary gland.
The Hypothalamus Can Be Subdivided in Both Longitudinal and Medial-Lateral Directions
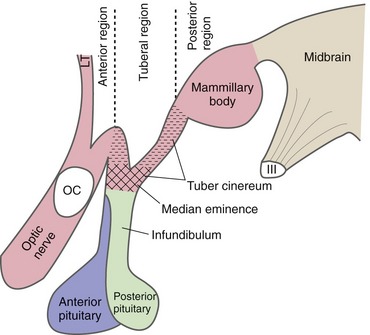
Parts of the hypothalamus are exposed at the base of the brain, surrounded by the circle of Willis. The mammillary bodies form the most posterior part of the hypothalamus and lie adjacent to the cerebral peduncles. Between the mammillary bodies and the optic chiasm and tract is a small swelling called the tuber cinereum. The median eminence arises from the tuber cinereum and narrows into the infundibulum, to which the pituitary gland is attached. These landmarks on the base of the brain are used to divide the hypothalamus longitudinally (Fig. 23-2) into an anterior region (above the optic chiasm, extending anteriorly to the lamina terminalis), a tuberal region (above and including the tuber cinereum), and a posterior region (above and including the mammillary bodies).
Hypothalamic Inputs Arise in Widespread Neural Sites
Key Concepts
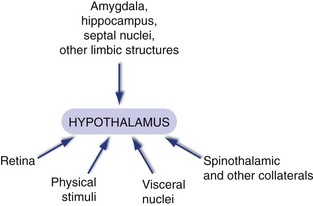
The hypothalamus receives lots of inputs (Fig. 23-3), but most of them are from two general categories: those from nuclei in the brainstem and spinal cord conveying information about the state of your body, and those from limbic structures like the amygdala, hippocampus, and septal nuclei. Inputs about the state of the body (“It’s getting warm in here,” or “Blood glucose is getting low”) arrive from places like the nucleus of the solitary tract by way of the dorsal longitudinal fasciculus, which travels through the periaqueductal gray into the periventricular zone; through the medial forebrain bundle, which travels through the reticular formation into the lateral hypothalamus; and as branches from tracts like the spinothalamic tract. Limbic inputs arrive from the amygdala, from the hippocampus (through the fornix), and from the septal nuclei and other sites (through the medial forebrain bundle); collectively they keep the hypothalamus updated on other aspects of the environment (“Not a good place to take off my shirt”).
Hypothalamic Outputs Largely Reciprocate Inputs
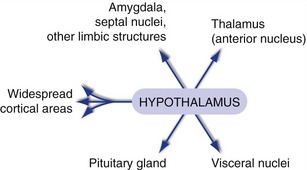
Hypothalamic connections with visceral nuclei and limbic structures are largely reciprocal (Fig. 23-4). Projections through the dorsal longitudinal fasciculus and the medial forebrain bundle reach sites like the nucleus of the solitary tract, the dorsal motor nucleus of the vagus, and the intermediolateral cell column of the spinal cord (“Better start sweating”). Projections through the medial forebrain bundle and other routes reach the amygdala, septal nuclei, and other limbic structures (“Maybe I can find the thermostat”). (Hypothalamic output reaches the hippocampus through a more circuitous route utilizing the thalamus, as described in Chapter 24.) In addition, diffuse modulatory projections to the thalamus and cerebral cortex play a key role in sleep-wake cycles (see Fig. 22-5Fig. 22-6).
The Hypothalamus Controls Both Lobes of the Pituitary Gland
The final, and major, hypothalamic outputs control the pituitary gland (hypophysis) through two separate mechanisms (Fig. 23-5). (1) Hypothalamic neurons in the supraoptic and paraventricular nuclei are the source of antidiuretic hormone (vasopressin) and oxytocin. They transport these hormones down their axons to the posterior lobe of the pituitary (most of the neurohypophysis), where they are released into the circulation. (2) Hypothalamic neurons in and near the tuber cinereum produce small peptides that serve as releasing and inhibiting factors for the anterior lobe of the pituitary (most of the adenohypophysis). They transport these factors down their axons and release them into capillaries in the median eminence. These capillaries then converge into pituitary portal vessels that travel down the infundibular stalk to a second capillary bed in the anterior pituitary. The releasing and inhibiting factors leave the second capillary bed and control the production of anterior pituitary hormones.
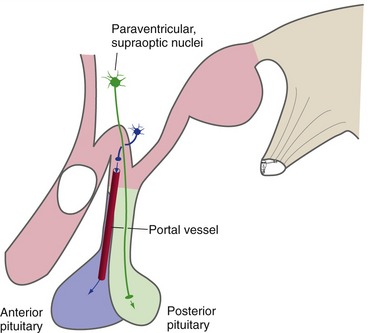
The Hypothalamus Collaborates with a Network of Brainstem and Spinal Cord Neurons
The connections of the hypothalamus position it to control the whole gamut of homeostatic mechanisms and drive-related behaviors—temperature regulation, feeding and drinking, cardiovascular function, sexual behavior, hormonal regulation, aggression, and on and on. Although some of these activities mainly involve autonomic adjustments (e.g., cardiovascular reflexes), skeletal muscle plays a major role in others (e.g., breathing). Normal control of micturition (urination) provides a nice example of coordination among visceral afferents, conscious awareness, smooth muscle, skeletal muscle, and voluntary control.
Normal Micturition Involves a Central Pattern Generator in the Pons
The bladder is a container that spends most of its time storing urine; it can do this because the pressure inside the bladder is usually low—the smooth muscle in its wall (the detrusor) is relaxed, and the internal (smooth muscle) and external (skeletal muscle) sphincters in its neck are contracted. This storage mode is maintained by sympathetic inputs and by tonic firing of motor neurons to the external sphincter (Fig. 23-6A). Periodically the sphincters relax, the detrusor contracts, and urine is eliminated. Infants do this automatically, using spinal cord reflex circuitry that is suppressed later in life. For adults, entering this elimination mode is more complicated because decisions need to be made about appropriate times and places to urinate. The periaqueductal gray sums up inputs from the spinal cord (tension in the bladder wall), the hypothalamus, and cerebral cortex, then passes the decision along to the pontine micturition center, which executes it (Fig. 23-6B).
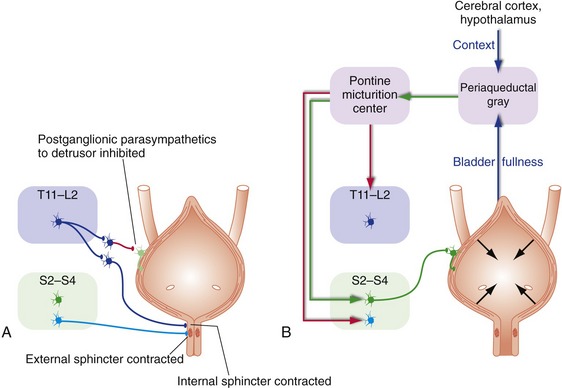
The Hypothalamus and Associated Central Pattern Generators Keep Physiological Variables within Narrow Limits
In a similar way, the hypothalamus monitors a host of other physiological variables and is involved in initiating the right mix of autonomic, hormonal, and behavioral responses to changes in these variables—secreting more antidiuretic hormone in response to increased plasma osmolality, for example, while at the same time contributing to a feeling of thirst through interactions with the limbic system.
Limbic Structures Are Interposed between the Hypothalamus and Neocortex
We perceive multiple attributes of things—not only their physical attributes, but also whether they are attractive, frightening, and so on. Integration of these multiple attributes occurs in the multimodal association areas of the cerebral cortex (see Fig. 22-3), using multiple inputs. Information about physical attributes comes from the unimodal association areas, whereas information about their drive-related attributes comes from limbic structures. Drive-related attributes of objects or situations also have implications for autonomic and behavioral responses, and these are mediated by the same limbic structures in conjunction with the hypothalamus and the adjacent septal area. Hence, the general notion of the limbic system is that limbic structures serve as a sort of bridge between neocortex and behavior when drives and emotions are involved (see Fig. 23-1).
The Hippocampus and Amygdala Are the Central Components of the Two Major Limbic Subsystems
The limbic system includes both cortical areas and noncortical structures and is divided into two subsystems, one centered on the hippocampus and the other on the amygdala. Each uses different areas of cerebral cortex as part of the bridge to multimodal association areas. The hippocampal subsystem is primarily involved in learning and memory, as described in Chapter 24.
The Amygdala Is Centrally Involved in Emotional Responses
The amygdala, a collection of nuclei located in the temporal lobe at the anterior end of the hippocampus, is a key link between experiences and emotional reactions to them. It receives a great deal of sensory information of all sorts, from the brainstem, thalamus, hypothalamus, olfactory bulb, and unimodal association areas (Fig. 23-7). It also receives inputs from anterior parts of the limbic lobe and nearby areas, including orbital and anterior temporal cortex and the insula. Connections with the hypothalamus travel through both the stria terminalis and a more diffuse pathway that passes underneath the lenticular nucleus. The stria terminalis, like the fornix, curves around with the lateral ventricle, but in this case travels just medial to the caudate nucleus; for much of its course it lies in the groove between the caudate nucleus and the thalamus.
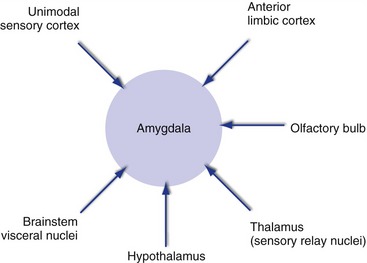
Emotional experiences are accompanied by conscious awareness of the emotion, autonomic reactions, and heightened awareness of ongoing events, and outputs from the amygdala (Fig. 23-8) help mediate all of these. Outputs to anterior limbic cortex, both directly and through the dorsomedial nucleus of the thalamus, contribute to conscious experience. Those to the hypothalamus and to brainstem visceral nuclei mediate autonomic responses. Those to sensory cortical areas are even more extensive than inputs and help to enhance performance when the pressure is on. In addition, outputs to the ventral striatum get the basal ganglia into the act, and outputs to the hippocampus affect the probability that an event will be remembered (see Chapter 24).
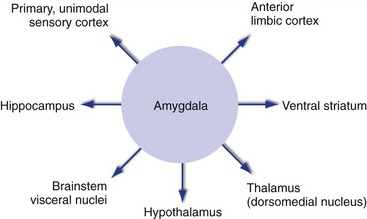
The Amygdala Is Involved in Emotion-Related Aspects of Learning
The amygdala uses its interconnections with limbic and sensory cortex and the hypothalamus both to form and to express associations between objects and the reactions they provoke. That is, while the hippocampus is important for remembering that an event occurred (see Chapter 24), the amygdala is important not only for remembering whether the event was “good” or “bad,” but also for triggering appropriate responses (“gut feelings”) the next time a similar event occurs.
[/level-membership-for-basic-science-category][not-level-membership-for-basic-science-category]
23 Drives and Emotions: The Hypothalamus and Limbic System
The Hypothalamus Coordinates Drive-Related Behaviors
The hypothalamus is a nodal point in the neural circuits underlying drive-related behaviors (Fig. 23-1). It’s got interconnections with visceral parts of the nervous system, through which it is informed of and controls things like blood glucose, blood pressure, and body temperature. It’s also got interconnections with limbic structures, through which you become aware of homeostatic needs (“I’m hungry”). Finally, the hypothalamus has not just neural outputs but also ways to control the pituitary gland.
The Hypothalamus Can Be Subdivided in Both Longitudinal and Medial-Lateral Directions
Parts of the hypothalamus are exposed at the base of the brain, surrounded by the circle of Willis. The mammillary bodies form the most posterior part of the hypothalamus and lie adjacent to the cerebral peduncles. Between the mammillary bodies and the optic chiasm and tract is a small swelling called the tuber cinereum. The median eminence arises from the tuber cinereum and narrows into the infundibulum, to which the pituitary gland is attached. These landmarks on the base of the brain are used to divide the hypothalamus longitudinally (Fig. 23-2) into an anterior region (above the optic chiasm, extending anteriorly to the lamina terminalis), a tuberal region (above and including the tuber cinereum), and a posterior region (above and including the mammillary bodies).
Hypothalamic Inputs Arise in Widespread Neural Sites
Key Concepts
The hypothalamus receives lots of inputs (Fig. 23-3), but most of them are from two general categories: those from nuclei in the brainstem and spinal cord conveying information about the state of your body, and those from limbic structures like the amygdala, hippocampus, and septal nuclei. Inputs about the state of the body (“It’s getting warm in here,” or “Blood glucose is getting low”) arrive from places like the nucleus of the solitary tract by way of the dorsal longitudinal fasciculus, which travels through the periaqueductal gray into the periventricular zone; through the medial forebrain bundle, which travels through the reticular formation into the lateral hypothalamus; and as branches from tracts like the spinothalamic tract. Limbic inputs arrive from the amygdala, from the hippocampus (through the fornix), and from the septal nuclei and other sites (through the medial forebrain bundle); collectively they keep the hypothalamus updated on other aspects of the environment (“Not a good place to take off my shirt”).
Hypothalamic Outputs Largely Reciprocate Inputs
Hypothalamic connections with visceral nuclei and limbic structures are largely reciprocal (Fig. 23-4). Projections through the dorsal longitudinal fasciculus and the medial forebrain bundle reach sites like the nucleus of the solitary tract, the dorsal motor nucleus of the vagus, and the intermediolateral cell column
[/not-level-membership-for-basic-science-category]
