12 Cranial Nerves and Their Nuclei
Cranial nerves and their central connections often look bewilderingly complicated, but for the most part they are actually arranged systematically.
Cranial Nerve Nuclei Have a Generally Predictable Arrangement
The olfactory nerve (I) is a series of thin filaments that attach directly to the olfactory bulb, part of the telencephalon. Fibers of the optic nerve (II) proceed through the optic chiasm and tract (see Chapter 17), and most end in the lateral geniculate nucleus of the thalamus, part of the diencephalon. The 10 remaining cranial nerves originate or terminate in the brainstem (or upper cervical spinal cord), as indicated in Table 12-1 and Fig. 12-1. The taste afferents in the facial, glossopharyngeal, and vagus nerves (VII, IX, X) are considered separately in Chapter 13, along with the olfactory nerve (I), and the vestibulocochlear nerve (VIII) has its own chapter (14). The rest of the brainstem cranial nerves are introduced in this chapter.
| Cranial Nerve | Main Functions | Attachment Point |
|---|---|---|
| III. Oculomotor | Eye movements, pupil, lens | Rostral midbrain (V) |
| IV. Trochlear | Eye movements (superior oblique) | Pons/midbrain junction (D) |
| V. Trigeminal | Facial sensation, chewing | Midpons (L) |
| VI. Abducens | Eye movements (lateral rectus) | Pontomedullary junction (V) |
| VII. Facial | Facial expression, taste | Pontomedullary junction (V/L) |
| VIII. Vestibulocochlear | Hearing, equilibrium | Pontomedullary junction (V/L) |
| IX. Glossopharyngeal | Taste, swallowing | Rostral medulla (L) |
| X. Vagus | Visceral sensory Speaking, swallowing Preganglionic parasympathetic |
Rostral medulla (L) |
| XI. Accessory | Head and shoulder movement | Upper cervical spinal cord (L) |
| XII. Hypoglossal | Tongue movement | Rostral medulla (V/L) |
D, L, and V indicate dorsal, lateral, and ventral attachment points.
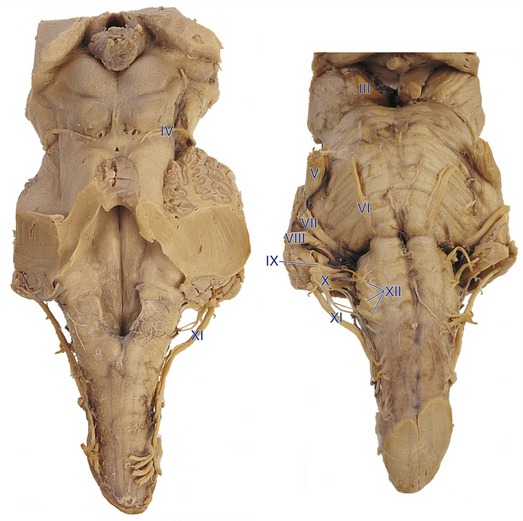
The wiring principles discussed in Chapter 3 and applied to the spinal cord in Chapter 10 also apply, for the most part, to cranial nerves and their connections. At its attachment point, each brainstem cranial nerve is carrying sensory information from ipsilateral receptors or motor output to ipsilateral muscles (except for the case of afferents and efferents for places like abdominal viscera, where the ipsilateral-contralateral concept loses much of its meaning). Thus, the right vestibulocochlear nerve (VIII) carries information from the right cochlea and the left oculomotor nerve (III) innervates muscles of the left eye. Similarly, the fibers of these cranial nerves mostly terminate in or originate from the ipsilateral side of the CNS. The principal exceptions are the trochlear (IV) and oculomotor nerves. All trochlear fibers and some oculomotor fibers originate from motor neurons in the contralateral half of the CNS.
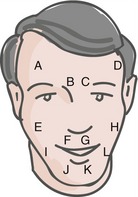
Some fairly simple rules can be used to predict the approximate locations of cranial nerve nuclei—collections of second-order sensory neurons, lower motor neurons, and preganglionic parasympathetic neurons. The medial-lateral location of a nucleus is predicted by the sulcus limitans, and the longitudinal location by the level of attachment of the cranial nerve associated with it (Fig. 12-2). Sensory nuclei are typically located lateral to the sulcus limitans and motor nuclei medial to it because of the way the neural tube opens up at the level of the fourth ventricle. Visceral nuclei are located closer to the sulcus limitans, so for example second-order taste neurons are just lateral to the sulcus limitans and visceral motor (= preganglionic parasympathetic) neurons just medial to it.
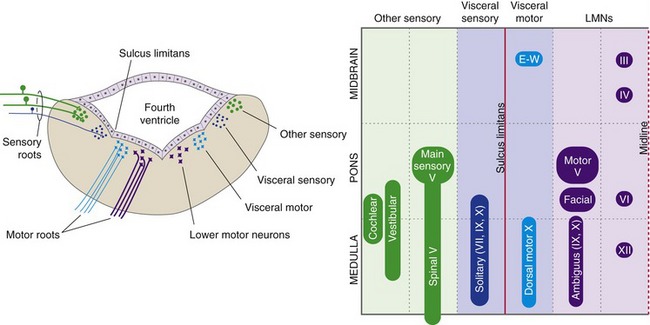
Cranial Nerves III, IV, VI, and XII Contain Somatic Motor Fibers
The trochlear nerve (IV) innervates the superior oblique muscle and the abducens nerve (VI) innervates the lateral rectus. The oculomotor nerve (III) innervates the remaining extraocular muscles (medial, superior, and inferior recti, inferior oblique) and the elevator of the eyelid, and it also contains the preganglionic parasympathetic fibers for the pupillary sphincter and the ciliary muscle. All three of these nerves, once they leave the brainstem, proceed to the ipsilateral eye (although trochlear and some oculomotor fibers cross before leaving the brainstem). The hypoglossal nerve (XII) innervates the muscles of the ipsilateral half of the tongue.
The Abducens Nucleus Also Contains Interneurons That Project to the Contralateral Oculomotor Nucleus
Any time we look to the left or the right, we need to contract the lateral rectus of one eye and the medial rectus of the other eye simultaneously. Theoretically, this could be accomplished by having separate, parallel inputs to both sets of motor neurons. However, we have evolved a different mechanism to achieve the same end. The abducens nucleus contains not only the motor neurons for the ipsilateral lateral rectus, but also an equal number of interneurons whose axons cross the midline, join the medial longitudinal fasciculus (MLF), and ascend to medial rectus motor neurons in the contralateral oculomotor nucleus (Fig. 12-3). Anything that stimulates lateral rectus motor neurons during attempted horizontal gaze also stimulates the interneurons, so coordinated gaze is achieved when we try to contract one lateral rectus.
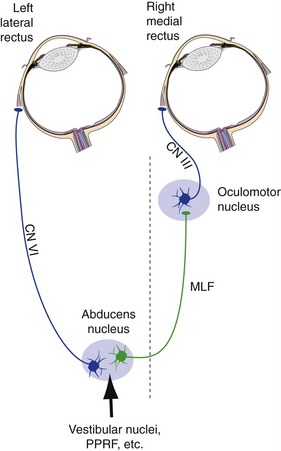
Inputs to the abducens nucleus arise in a number of sites, including the vestibular nuclei. (This allows us to generate eye movements equal and opposite to head movements—the vestibulo-ocular reflex, covered further in Chapter 14.) Ordinary voluntary eye movements are of a rapid type called saccades. Making a rapid eye movement over to a target requires a rapid initial burst of action potentials in abducens motor neurons and interneurons to get the eyes moving, followed by slower maintained firing to maintain the new position. The required timing signals are generated in the reticular formation near the abducens nucleus, a region called the paramedian pontine reticular formation (or PPRF, or pontine gaze center). Hence, damage to one abducens nucleus causes loss of all horizontal eye movements to the ipsilateral side, but damage to the PPRF causes selective loss of rapid eye movements to the ipsilateral side. Damage to one MLF causes selective weakness of the ipsilateral medial rectus during attempted lateral gaze. (Eye movements are discussed more fully in Chapter 21.)
The Hypoglossal Nerve (XII) Innervates Tongue Muscles
Upper motor neurons for tongue motor neurons, like those for spinal cord motor neurons, are located in the precentral gyrus and adjacent cortical areas. Their axons, called corticobulbar fibers, accompany corticospinal axons through the internal capsule, cerebral peduncle, and pons. In the rostral medulla, they leave the corticospinal fibers and end in the hypoglossal nucleus. As they do so, many but not all of them cross the midline. Hence, both sides of the tongue can be made to contract by one cerebral hemisphere, and unilateral damage to corticobulbar fibers does not cause profound or lasting weakness of either side of the tongue. However, most of us have substantially more crossed than uncrossed fibers, so there is often some transient weakness on the contralateral side following corticobulbar damage. In such a case, cortical damage would cause tongue deviation away from the side of the lesion because the weak side would be contralateral to the lesion.
Branchiomeric Nerves Contain Axons from Multiple Categories
The trigeminal (V), facial (VII), glossopharyngeal (IX), vagus (X), and accessory (XI) nerves are more complex than the ones considered so far. Each innervates skeletal muscle derived embryologically from the branchial arches. Except for the accessory nerve, each innervates other things, too. Nevertheless, each has only one or two major functions (see Table 12-1 at the beginning of this chapter).
The Trigeminal Nerve (V) Is the General Sensory Nerve for the Head
The Main Sensory Nucleus Receives Information about Touch and Jaw Position
Large-diameter trigeminal afferents, carrying information about touch and jaw position, end in the main sensory nucleus (Fig. 12-4). The main sensory nucleus is the trigeminal equivalent of a posterior column nucleus. Second-order fibers from the main sensory nucleus cross the midline, join the medial lemniscus, and reach the ventral posteromedial nucleus (VPM) of the thalamus. VPM neurons then project to somatosensory cortex of the postcentral gyrus and adjacent areas. In contrast to the situation in the spinal cord, large-diameter trigeminal afferents do not travel through anything comparable to the posterior columns before reaching their nucleus of termination (Fig. 12-5).
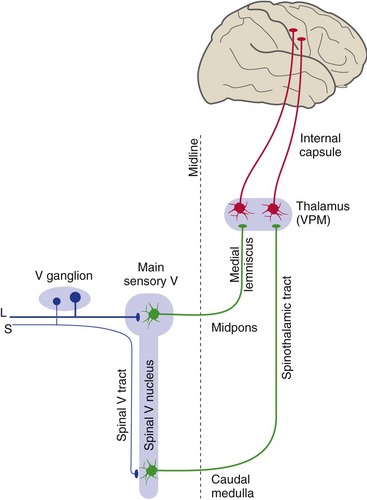
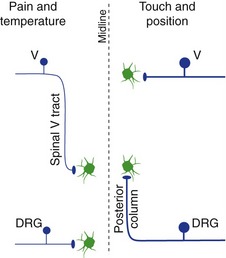
The Spinal Trigeminal Nucleus Receives Information about Pain and Temperature
Small-diameter trigeminal afferents, carrying pain and temperature and some touch information, turn caudally in the spinal trigeminal tract. The pain and temperature fibers end in portions of the spinal trigeminal nucleus in the caudal medulla and upper cervical spinal cord. Second-order fibers from the spinal trigeminal nucleus then cross the midline, join the spinothalamic tract, and reach VPM and other thalamic nuclei (see Fig. 12-4). Thalamic neurons then project to somatosensory cortex of the postcentral gyrus and other areas. In contrast to the situation in the spinal cord, small-diameter trigeminal afferents travel an appreciable distance through the CNS (in the spinal trigeminal tract) before reaching the second-order neurons on which they terminate (see Fig. 12-5).
The primary afferent fibers in the spinal trigeminal tract are arranged somatotopically (Fig. 12-6), with the ipsilateral half of the face represented upside down (i.e., ophthalmic division fibers most ventral, mandibular division fibers most dorsal). In addition to this dorsal-ventral gradient, there is a rostral-caudal order in which the pain and temperature fibers terminate in the spinal trigeminal nucleus. Fibers representing areas near the midline terminate most rostrally (i.e., in the mid-medulla), and fibers representing more lateral areas terminate most caudally (i.e., in the upper cervical spinal cord). This makes sense because it means that pain and temperature afferents from the back of the head (i.e., upper cervical dorsal roots) end near the termination sites of trigeminal afferents from nearby areas of the head.
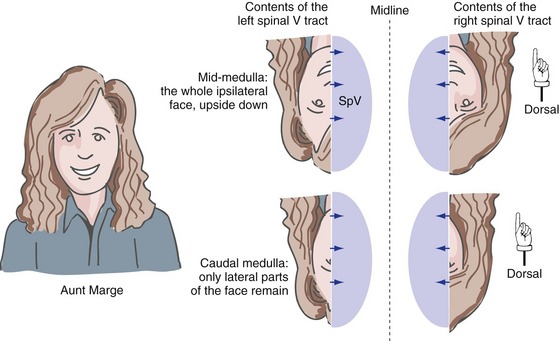
The rest of the spinal trigeminal nucleus (i.e., the parts in the rostral medulla and caudal pons) takes care of the remaining spinal-cord-like somatosensory functions for the head. These include things like getting trigeminal information to the cerebellum, as well as providing the interneurons for the blink reflex (see Fig. 12-8), which is a trigeminal equivalent of a withdrawal reflex.
The Trigeminal Motor Nucleus Innervates Muscles of Mastication
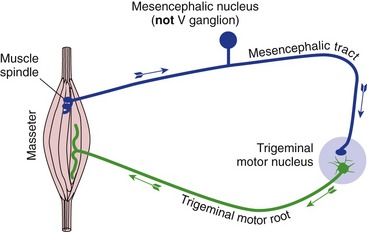
Trigeminal motor neurons are involved not just in routine chewing, but also in a jaw-jerk reflex (Fig. 12-7) homologous to the knee-jerk reflex: stretching the masseter causes a reflex contraction by way of a monosynaptic circuit parallel to that of spinal stretch reflexes. The afferent cell bodies for this reflex are peculiar in that they reside within the CNS (in the mesencephalic trigeminal nucleus).
The Facial Nerve (VII) Innervates Muscles of Facial Expression
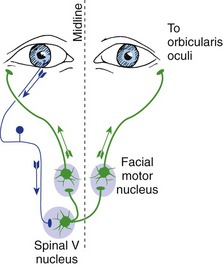
The jaw-jerk reflex is the only prominent monosynaptic cranial nerve reflex. Other brainstem reflexes, like other spinal reflexes, involve interneurons. Many of these reflexes are protective in nature, like the spinal flexor (withdrawal) reflex. An example is blinking both eyes when an object touches either cornea. The afferents for this blink reflex (Fig. 12-8) travel in the trigeminal nerve, the efferents (cell bodies in the facial motor nucleus) in the facial nerve.
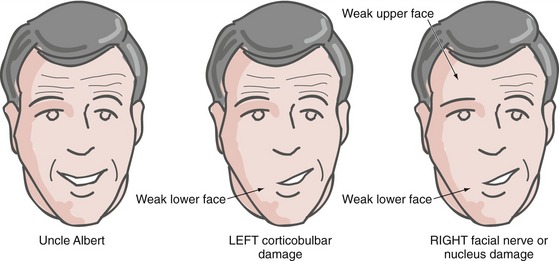
Upper Motor Neuron Damage Affects the Upper and Lower Parts of the Face Differently
Corticobulbar fibers to the motor neurons for the lower half of the face are mostly crossed, corresponding to the way we can move both sides of the lower face relatively independently. However, corticobulbar fibers to the motor neurons for the upper half of the face are distributed bilaterally, corresponding to the way both sides of the upper face usually move together. Hence, all parts of the face except the ipsilateral lower quadrant can be made to contract by one cerebral hemisphere, and so unilateral corticobulbar damage causes weakness of only the contralateral lower quadrant (Fig. 12-9). (For example, after left motor cortex damage the intact right hemisphere cannot make the muscles of the right lower quadrant contract.) In contrast, unilateral damage to the facial nerve or nucleus causes weakness of the entire ipsilateral half of the face.
The Vagus Nerve (X) Is the Principal Parasympathetic Nerve
Afferent fibers from the thoracic and abdominal viscera reach the brainstem with the vagus nerve. They travel within the brainstem in the solitary tract of the rostral medulla and end in the surrounding nucleus of the solitary tract (Fig. 12-10). A smaller number of glossopharyngeal afferents, mostly from the mouth and throat, follow a similar course in the CNS. Neurons in the nucleus of the solitary tract participate in various visceral reflexes (e.g., automatic adjustment of respiratory and cardiovascular functions) and also convey information directly or indirectly to the hypothalamus and thalamus.
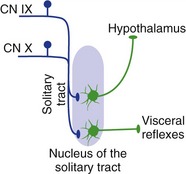
Figure 12-10 Visceral connections of the nucleus of the solitary tract. (As discussed in Chapter 13, this nucleus also has a critical role in taste.)
Motor neurons for the muscles of the larynx and pharynx are located in the reticular formation of the rostral medulla, just dorsal to the inferior olivary nucleus, as nucleus ambiguus. Most of them travel peripherally in the vagus nerve (X) to ipsilateral muscles; a few travel in the glossopharyngeal nerve (IX).
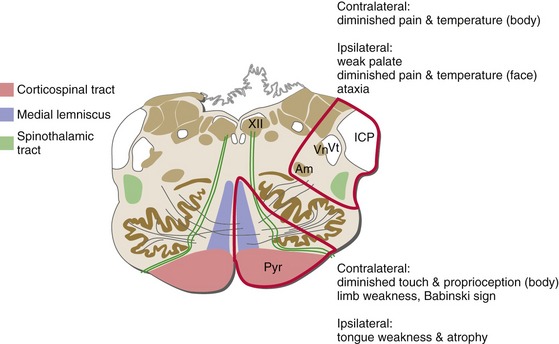
Brainstem Damage Commonly Causes Deficits on One Side of the Head and the Opposite Side of the Body
Once the medial lemniscus has formed, all the major long tracts in the brainstem are related to the contralateral side of the body. In contrast, fibers entering or leaving the brainstem in cranial nerves are hooked up to ipsilateral structures. The result is that damage on one side of the brainstem causes contralateral problems in the body, but ipsilateral problems in the head (Fig. 12-11). These kinds of crossed findings are hard to produce with damage anyplace else in the nervous system and are usually a dead giveaway that it’s a brainstem lesion. Knowing the locations of the long tracts and of cranial nerve attachment levels often lets you localize brainstem damage with amazing precision.
For questions 1-4, choose the most likely site of damage from the following list.
| LEFT | RIGHT |
| a. Medulla | d. Medulla |
| b. Pons | e. Pons |
| c. Midbrain | f. Midbrain |