[level-membership-for-basic-science-category]
21 Control of Eye Movements
Six Extraocular Muscles Move the Eye in the Orbit
We need to move each eye in various combinations of six directions. Four of them are obvious—medially (adduction), laterally (abduction), up (elevation), and down (depression). The two others are torsional movements, the kind you would make to keep an eye level as you tilt your head to one side or the other. Intorsion rotates the top of the eye closer to the nose and extorsion rotates it away. Movements in these six directions are accomplished by six small extraocular muscles, but the correspondence between movements and individual muscles is not always direct (Table 21-1).
Table 21-1 Extraocular muscles, eye movements, and cranial nerves
| Movement | Principal Muscle | Other Contributors |
|---|---|---|
| Abduction | Lateral rectus (VI) | Inferior oblique (III) Superior oblique (IV) |
| Adduction | Medial rectus (III) | Inferior rectus (III) Superior rectus (III) |
| Depression | Inferior rectus (III) | Superior oblique (IV) |
| Elevation | Superior rectus (III) | Inferior oblique (III) |
| Extorsion | Inferior oblique (III) | Inferior rectus (III) |
| Intorsion | Superior oblique (III) | Superior rectus (III) |
The Superior and Inferior Recti and the Obliques Have More Complex Actions
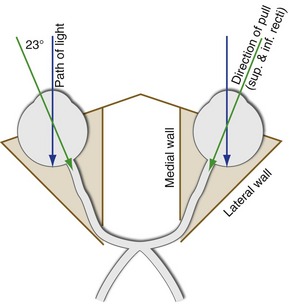
The four remaining muscles—the superior rectus, inferior rectus, superior oblique, and inferior oblique—do not lie entirely in the same plane as one of the directions of eye movement, so their actions are more complex. For example, the eye (when looking at something far away) points straight ahead in the orbit, but the axis of the orbit itself—the direction in which the superior and inferior recti pull—points not only backward but also toward the nose (Fig. 21-1). The result is that contraction of the superior rectus mainly causes elevation, but also pulls the top of the eye toward the nose (i.e., intorsion and adduction). Similarly, the inferior rectus mainly causes depression, but also causes extorsion and adduction. The superior and inferior obliques mainly cause intorsion and extorsion, respectively. However, because they insert behind the middle of the eye and pull partially anteriorly, they too cause movement in additional directions (see Table 21-1).
There Are Fast and Slow Conjugate Eye Movements
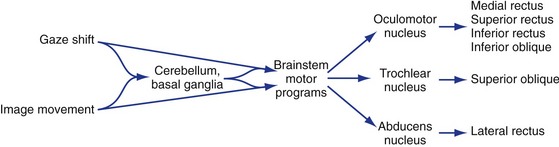
Just as there are motor programs for things like walking that can be modulated by descending projections from places like motor cortex, there are groups of subcortical neurons specialized to generate the timing signals for fast and slow eye movements and pass them along to the oculomotor, trochlear, and abducens nuclei (Fig. 21-2). These timing centers receive inputs from those parts of the brain that can initiate eye movements and then send their outputs to the appropriate motor neurons; just as in the case of other movements, the cerebellum and basal ganglia play a role in planning and coordinating eye movements. Superimposed on this arrangement are projections from the vestibular nuclei, so that we can adjust eye position to compensate for head movements.
Fast, Ballistic Eye Movements Get Images onto the Fovea
Saccades are rapid conjugate movements (Fig. 21-3), in which our eyes can move as rapidly as 700°/second. We use saccades for voluntary eye movements, to look over at something we caught a glimpse of in the periphery, to catch up with something that’s moving too fast to track, and as the fast phase of nystagmus. Moving your eyes like this is harder than it seems. It requires a very rapid burst (up to 1000 impulses/second) in the motor neurons to generate the velocity, and then a carefully calculated maintained firing rate to keep the eyes in their new position. Saccades are prepackaged movements, as though the brain calculates how far we need to move, sets up the timing, and then lets it fly. Once it starts, the saccade usually can’t be changed, for example if the target moves again. One of the few ways a saccade can be modified is through the vestibular nuclei. If you move your head during a saccade, the vestibulo-ocular reflex (VOR, see Fig. 14-11) automatically compensates for the movement.
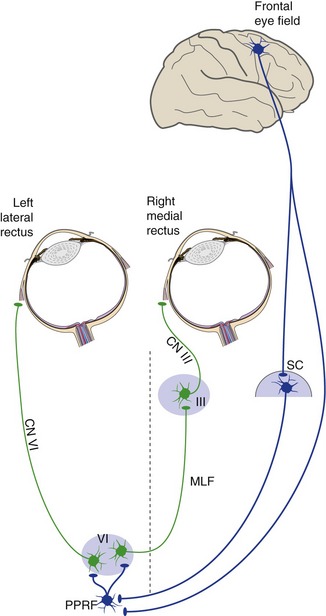
For horizontal conjugate movements, things aren’t quite that simple because we need to coordinate the lateral rectus of one eye with the medial rectus of the other eye. This is accomplished by having not only motor neurons in each abducens nucleus, but also interneurons that project through the contralateral MLF to the oculomotor nucleus (see Fig. 12-3). (These abducens interneurons are activated not just during saccades, but during all horizontal movements to the ipsilateral side.) The timing signals are generated in the paramedian pontine reticular formation (PPRF) near the abducens nucleus. The PPRF on one side of the pons sets up the signals for saccades to the ipsilateral side.
The Frontal Eye Fields and Superior Colliculus Trigger Saccades to the Contralateral Side
Because saccades are used prominently in voluntary eye movements, it is not surprising that they can be triggered from the frontal lobes. The specific area involved, called the frontal eye field, is just anterior to where the head is represented in motor cortex. A unilateral frontal lesion causes no problems with vertical saccades, so vertical eye movements are apparently represented bilaterally. In the case of horizontal saccades, each hemisphere triggers movements to the contralateral side (Fig. 21-4). However, after unilateral frontal lesions, even horizontal saccades recover quickly (usually in a matter of days). How much of this recovery depends on the other frontal lobe or other cortical areas, and how much depends on the superior colliculus, is unclear.
Slow, Guided Eye Movements Keep Images on the Fovea
Smooth Pursuit Movements Compensate for Target Movement
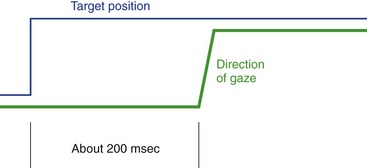
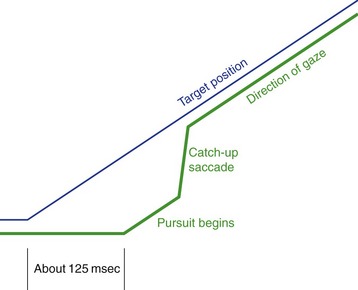
We use pursuit or tracking movements to track a moving object once its image is on or near the fovea. Pursuit movements can go at a maximum rate of only 50°/second or so. As a result, rapidly or irregularly moving objects require a combination of saccades and pursuit movements. Also, there’s a latency of about 125 msec for pursuit movements when a target starts to move, so by the time we start to track something, its image has moved off the fovea; the CNS keeps track of all this and produces a catch-up saccade when required (Fig. 21-5).
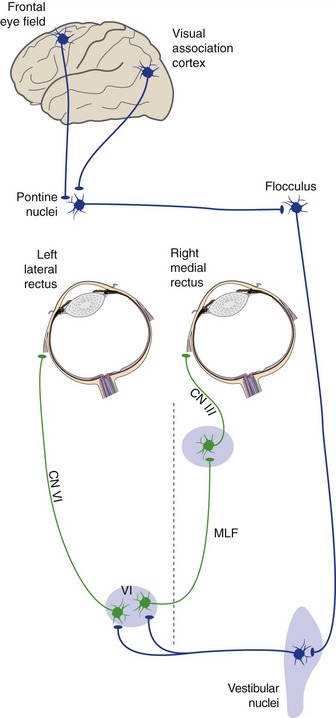
Even though pursuit movements can get started faster than saccades (125 vs. 200 msec), they use what looks like a considerably more circuitous pathway. Signals from motion-sensitive areas of visual association cortex and from the frontal eye field reach a particular small group of pontine nuclei, and then in succession the flocculus, vestibular nuclei, and the abducens, trochlear, and oculomotor nuclei (Fig. 21-6). (This is probably because of an evolutionary relationship between pursuit movements and VOR suppression.) As in the case of saccades, vertical movements are triggered bilaterally. Oddly enough, each cerebral hemisphere is more involved in triggering horizontal pursuit movements to the ipsilateral side (probably a by-product of the evolutionary relationship to cerebellar circuitry).
Changes in Object Distance Require Vergence Movements
Vergence movements are part of the near reflex (see Fig. 17-9). The afferent limb of the reflex is the normal visual pathway from eyeball to occipital lobe. Visual association cortex of the occipital lobe then projects to the efferent machinery in the midbrain.
The Basal Ganglia and Cerebellum Participate in Eye Movement Control
One of the parallel loops through the basal ganglia (see Fig. 19-4) passes through the frontal eye field and the caudate nucleus and substantia nigra (reticular part). This loop reaches not only the thalamus, but also the superior colliculus. Patients with basal ganglia disorders may have involuntary, small, or slow eye movements, much like the movements of other parts of their bodies.
[/level-membership-for-basic-science-category][not-level-membership-for-basic-science-category]
21 Control of Eye Movements
Six Extraocular Muscles Move the Eye in the Orbit
We need to move each eye in various combinations of six directions. Four of them are obvious—medially (adduction), laterally (abduction), up (elevation), and down (depression). The two others are torsional movements, the kind you would make to keep an eye level as you tilt your head to one side or the other. Intorsion rotates the top of the eye closer to the nose and extorsion rotates it away. Movements in these six directions are accomplished by six small extraocular muscles, but the correspondence between movements and individual muscles is not always direct (Table 21-1).
Table 21-1 Extraocular muscles, eye movements, and cranial nerves
| Movement | Principal Muscle | Other Contributors |
|---|---|---|
| Abduction | Lateral rectus (VI) | Inferior oblique (III) Superior oblique (IV) |
| Adduction | Medial rectus (III) | Inferior rectus (III) Superior rectus (III) |
| Depression | Inferior rectus (III) | Superior oblique (IV) |
| Elevation | Superior rectus (III) | Inferior oblique (III) |
| Extorsion | Inferior oblique (III) | Inferior rectus (III) |
| Intorsion | Superior oblique (III) | Superior rectus (III) |
The Superior and Inferior Recti and the Obliques Have More Complex Actions
The four remaining muscles—the superior rectus, inferior rectus, superior oblique, and inferior oblique—do not lie entirely in the same plane as one of the directions of eye movement, so their actions are more complex. For example, the eye (when looking at something far away) points straight ahead in the orbit, but the axis of the orbit itself—the direction in which the superior and inferior recti pull—points not only backward but also toward the nose (Fig. 21-1). The result is that contraction of the superior rectus mainly causes elevation, but also pulls the top of the eye toward the nose (i.e., intorsion and adduction). Similarly, the inferior rectus mainly causes depression, but also causes extorsion and adduction. The superior and inferior obliques mainly cause intorsion and extorsion, respectively. However, because they insert behind the middle of the eye and pull partially anteriorly, they too cause movement in additional directions (see Table 21-1).
