Chapter 9 Circulatory limits to acute exercise
Dynamic work capacity is proportional to the amount of aerobic metabolism that can take place in the cells of the locomotor skeletal muscles. Although the efficiency of these metabolic processes can be manipulated by, for example, dietary selection so as to optimize the appropriate substrates and training-induced adaptation of muscle fibre type, the end result is a reflection of oxygen availability. As such, the primary limit to exercise is the rate at which oxygen can be delivered to the muscle.
As discussed in Chapter 8 (p. 99), ventilatory function and pulmonary oxygen uptake in the pulmonary capillaries are not normally limiting factors since, even at peak workloads, arterial oxygen saturation is similar to that at rest, except in the subset of elite athletes with exceptional cardiac adaptations to training. However, we saw in Chapter 3 that the volume of blood that can be pumped around the circulation per unit time is determined by the age-limited maximum heart rate and by the decline in ventricular filling at high heart rates. These changes set an absolute limit of around 0.3 l/min/kg to cardiac output in an untrained individual, equating to around 21 l/min in a 70 kg person. The purpose of the current chapter is to examine in detail the factors that limit cardiac output at different exercise durations and intensities, under different environmental circumstances and in relation to different types of individual.
FACTORS THAT LIMIT MUSCLE PERFUSION
Effects of exercise hyperaemia
In resting muscle, where there is a relatively high precapillary vasoconstrictor tone, mean capillary hydrostatic pressure and plasma oncotic pressure are both around 25 mmHg, so that there is a balance between inwardly and outwardly directed pressure gradients and no net water movement occurs across the capillary wall. When the muscle arterioles dilate with onset of exercise, however, the resulting fall in precapillary resistance automatically raises capillary hydrostatic pressure, to values as high as 50–60 mmHg when the arterioles are maximally dilated. Because this is far above the opposing oncotic pressure value, net extravasation of plasma water takes place until the resultant increase in interstitial hydrostatic pressure is sufficient to rebalance the outward/inward forces (Fortney et al 1981). At least at high muscle workloads, the outward water movement is increased further by the rise in interstitial osmolality caused by accumulation of muscle metabolites.
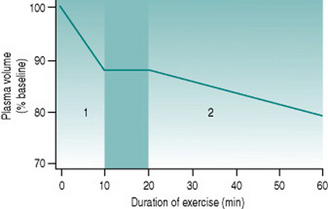
Since skeletal muscle makes up such a large proportion of total body mass, the volume of extravasated fluid within the muscles during whole-body dynamic exercise can be appreciable. At relatively heavy workloads, 10–12% of plasma volume (around 300 ml in a 70 kg person) is lost over the initial 10 min of activity (Fig. 9.1). Thus, over this period cardiac output also falls by around 300 ml provided heart rate remains constant, necessarily reducing muscle blood flow. Moreover, plasma loss results in a concomitant rise in haematocrit, elevating blood viscosity and so increasing cardiac workload.
Effect of heat production
As mentioned in Chapter 6 (p. 68), the first of these processes is withdrawal of sympathetic vasoconstrictor tone from the cutaneous arterioles. In parallel, there is activation of cholinergic sympathetic nerves to eccrine sweat glands. Both the loss of sympathetic tone and the process of sweat production cause cutaneous vasodilation. As a result, skin blood flow rises from a resting value that is typically less than 500 ml/min at environmental temperatures below 25° C to as much as 3 l/min after 15–20 min of sustained heavy exercise, with a similar fall in muscle blood flow. The dilatation due to sweating is at least partly secondary to glandular release of kinins (see Chapter 6, p. 75) but may also involve release of unknown dilator factors from the sudomotor nerve endings (Joyner & Halliwill 2000). As well as serving to convey heat to the body/environment interface, the high skin blood flow is essential to provide water and electrolytes for sweat production. This typically reaches a maximum of around 1 l/h, but may be as high as 2 l/h in a person who exercises routinely in hot conditions.
Sweating produces a second phase of plasma volume reduction that typically becomes measurable around 20 min after commencement of exercise (Fig. 9.1). Although in theory plasma and interstitial fluids are in equilibrium, high rates of sweat production draw primarily on the plasma, so that blood volume falls and haematocrit rises progressively as the exercise bout continues. Both effects make muscle oxygenation progressively more and more inefficient.
Measurement of sweating
A wide range of techniques are available for measuring sweat production, but none are suitable for all applications. Choice of technique depends critically on the intensity of sweat secretion to be measured and the duration of measurement required. All techniques in routine use involve one of four basic principles, as summarized below.
Volume collection
Except in very humid environments, sweat evaporates rapidly from the skin, even at quite high rates of secretion. If, however, a container is placed on the skin surface so as to prevent evaporative loss, then the sweat produced over that area of skin will be retained and can be measured. For obvious reasons, this technique cannot be used to detect the onset of sweating and is not practicable with very low sweat rates. But at high rates of production it can provide accurate quantitative values, provided that evaporative loss is prevented during the process of measurement and that the duration of collection is appropriate for the volume of the sampling vessel. One advantage of this method is the capacity to make repeated measurements for the same set of sweat glands over a prolonged period. A limitation is the lack of information obtained on behaviour of sweat glands elsewhere on the body.
Heart rate drift
During sustained dynamic exercise, heart rate begins to drift upwards from its previously stable level. The drift is prevented almost entirely by adequate fluid replacement during exercise, confirming that it involves reflex sympathetic compensation for the progressive fall in circulating plasma volume. In addition, prolonged muscle activity tends to involve progressive recruitment of larger numbers of muscle units as compensation for fatigue of those first recruited. Since greater descending motor drive is associated with greater activation of the cardiovascular centre, this also increases cardiac sympathetic drive.
Competition with other metabolically active tissues
One of these is that, in large-bodied, highly trained athletes, the metabolic demand of the respiratory muscles can be a potential limiting factor in locomotor muscle perfusion during intense activity (Dempsey et al 2006). In other subjects the amount of tissue involved is not sufficient to impose a significant perfusional load but, even so, it is possible that chemoreceptor inputs from metabolite build-up in respiratory muscles may contribute to the sensation of fatigue during intense activity.
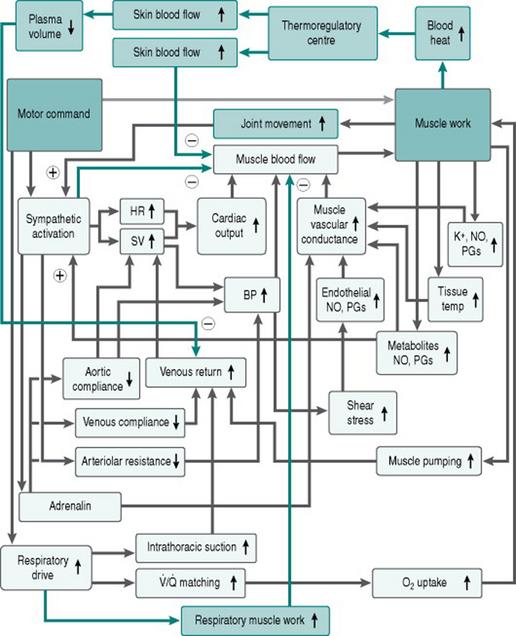
Figure 9.2 adds these limiting factors to our flow chart of the matrix of responses to exercise.
EFFECTS OF DIFFERENT ENVIRONMENTS
Temperature and humidity
Since the need for heat loss imposes major limitations on muscle blood flow, it is predictable that environments that affect thermoregulatory efficiency will influence exercise capacity. Thus, time to fatigue is increased under cold conditions due to both reduced skin blood flow and sweating (Blanchet et al 2003). During rapid exposure to a very cold environment (such as on immersion in cold water) there is profound cutaneous vasoconstriction that can result in dramatic elevation of blood pressure and it is possible that, even with less extreme activation of skin cold receptors, the pressure gradient for muscle perfusion during exercise will be somewhat higher than under thermoneutral conditions. It is, however, not known whether this has any real effect on the efficiency of muscle perfusion.
In cold environments, other issues that combine circulatory and thermoregulatory considerations may complicate the situation further. Thus, varying efficiencies of insulative clothing may create microenvironments that range from one that tends towards producing hypothermia due to evaporative and convective cooling to one that retards heat loss so much that it produces hyperthermia. In addition, extreme cold especially in windy conditions can overwhelm the cold dilator response that normally protects digits, ears and nose (see Chapter 5, p. 58) and result in freezing of cells in these tissues (frostbite).
Aquatic vs. terrestrial exercise
The relative inefficiency of swimming due to the high density of the medium is to some extent countered by the absence of some of the problems that limit terrestrial exercise. The high thermal conductivity of water means that heat loss occurs very rapidly and without the need for massive cutaneous vasodilation or sweating. As well, effective circulating blood volume is increased, both by lack of gravitational pooling and by the fact that the external pressure of the water compresses the superficial veins. Together, these factors allow muscle perfusion to be a higher proportion of cardiac output than during terrestrial activity and for it to be maintained stable for far longer periods.
EFFECTS OF AGE
Children
Cardiovascular factors
The cardiovascular systems of children and adolescents differ quantitatively from those of adults in several ways that impact on the response to exercise. First, children have lower heart sizes than adults relative to body mass, so their stroke volumes are lower. At any age, heart mass and stroke volume are slightly lower in girls than in boys. While heart size rises with age, stroke volume remains less than that in adults until late adolescence. In addition, studies that have investigated responses to maximum exercise in children and adolescents have been unable to show that the ceiling heart rate is significantly higher than the 200 beats/min typical of a 20-year-old (see Chapter 3, p. 20). Therefore, the relatively low stroke volumes in children mean that given increments of cardiac output require greater increments in heart rate than in adults, and the maximal cardiac output that can be achieved must be assumed to be lower than that in adults.
Nonetheless, there is more variability in maximum heart rate among children of any age than would be seen in adults and some children show maxima as high as 220 beats/min (Dencker et al 2006). On the basis of these findings it has been suggested that the true maximum rate is, in fact, greater than in adults and that failure to see this consistently is because children are often not motivated to reach their true maximum workload during testing. At present, the situation remains unresolved but, in the absence of an alternate consensus view, it is safest to assume that age-related changes in maximum heart rate begin only after adolescence.
A further difference between children and adults is that children of both sexes below the age of puberty have haematocrits that are slightly lower than those after puberty, at around 36–38%. Intuitively, the combination of lower cardiac output and lower oxygen carrying capacity would be thought to limit exercise in children. Nonetheless, these differences appear to confer little or no cardiovascular disadvantage on the capacity of children to produce short-term aerobic work, because they have greater capacity than adults for oxygen extraction in exercising muscles (Turley & Wilmore 1997). The mechanisms underlying this are not certain. Probable contributing factors include differences in muscle cell metabolism and the fact that smaller muscles have shorter diffusional distances between plasma and muscle cell.
Thermoregulatory factors
Nonetheless, children appear to be at least as efficient as adults in maintaining body temperature during exercise in hot conditions (Inbar et al 2004). Their higher surface area:body mass ratio contributes to this. As well, the higher sweat rates seen in adults may not provide more effective evaporation because some of the sweat drips off the skin.
Old age
Thermoregulatory factors
The elderly subject also faces additional thermoregulatory challenges during exercise. The lower absolute cardiac output imposes a stricter quota on partitioning of blood flows between muscle and skin as exercise progresses. Skin temperature rises more slowly during prolonged exercise, potentially limiting thermoregulatory stability. One reason for this is the overall increase in vascular resistance due to loss of elastic tissue, limiting skin blood flow. There is also evidence that ageing desensitizes the hypothalamic thermostatic centre, so that greater increments in blood temperature are required before heat loss processes are activated. Also, the gain of many autonomic reflexes falls with age, so that there may be less sudomotor discharge. Finally, atrophic changes in ageing skin are associated with decreased size of individual sweat glands.
EFFECTS OF PREGNANCY
Third, the additional heat load produced by the foetoplacental unit may result in body temperature rising more rapidly and further than normal during exercise. However, although the undoubted increase in absolute heat production during a given exercise intensity highlights potential dangers for intense exercise in hot environments, there is no good evidence of pregnant women being more susceptible to hyperthermia during moderate exercise in non-extreme environments. This is due probably to the vasodilator effect of oestrogens on skin blood vessels, together with the fact that progesterone raises the set point of the hypothalamic thermostat by around 0.5° C (0.3° F), increasing the gradient for heat diffusion into the environment except at very high ambient temperatures.
Blanchet M, Ducharme A, Racine N, et al. Effects of cold exposure on submaximal exercise performance and adrenergic activation in patients with congestive heart failure and the effects of beta-adrenergic blockade (carvedilol or metoprolol). American Journal of Cardiology. 2003;92:548-553.
Dempsey JA, Romer L, Rodman J, Miller J, Smith C. Consequences of exercise-induced respiratory muscle work. Respiratory Physiology and Neurobiology. 2006;151:242-250.
Dencker M, Thorsson O, Karlsson M, et al. Daily physical activity and its relation to physical fitness in children aged 8–11 years. European Journal of Applied Physiology. 2006;96:587-592.
Fortney SM, Nadel ER, Wenger CB, Bove JR. Effect of blood volume on sweating rate and body fluids in exercising humans. Journal of Applied Physiology. 1981;51:1594-1600.
Inbar O, Morris N, Epstein Y, Gass G. Comparison of thermoregulatory responses to exercise in dry heat among prepubertal boys, young adults and older males. Experimental Physiology. 2004;89:691-700.
Joyner MJ, Halliwill JR. Sympathetic vasodilatation in human limbs. Journal of Physiology. 2000;526:471-480.
Turley KR, Wilmore JH. Cardiovascular responses to treadmill and cycle ergometer exercise in children and adults. Journal of Applied Physiology. 1997;83:948-957.