20 Cerebellum
The cerebellum helps coordinate movement by sampling most kinds of sensory information, comparing current movements with intended movements, and issuing planning or correcting signals. The comparisons are made in a uniform, precisely organized, cerebellar cortex and the planning or correcting signals are issued through a set of deep cerebellar nuclei. Because its output is concerned with coordination of movement and not with perception, cerebellar lesions cause incoordination but no sensory changes.
The Cerebellum Can Be Divided into Transverse and Longitudinal Zones
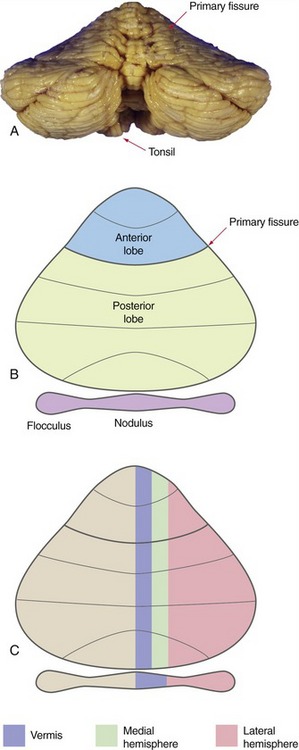
In a gross anatomical sense, the primary fissure divides the bulk of the cerebellum into anterior and posterior lobes, and another deep fissure separates the flocculus and nodulus (together forming the flocculonodular lobe) from the body of the cerebellum (Fig. 20-1). Assorted exotic names are sometimes applied to various parts of the anterior and posterior lobes, but most are of limited clinical utility. One worth remembering is the tonsil, the part of the posterior lobe nearest to the flocculus (THB6 Figure 20-2D, p. 496). The tonsil is one of the most inferior parts of the cerebellum, and expanding masses in the posterior fossa can cause it to herniate through the foramen magnum, compressing the medulla (THB6 Figure 4-19D, p. 96).
Deep Nuclei Are Embedded in the Cerebellar White Matter
The fundamental building plan of the cerebellum as a whole involves afferents that reach the cerebellar cortex, which in turn projects to deep nuclei embedded in the cerebellar white matter. The deep nuclei then give rise to the output of the cerebellum. There are a series of three deep nuclei on each side, arranged in a medial to lateral array: the fastigial (most medial), interposed, and dentate (most lateral) nuclei (THB6 Figure 20-7, p. 501).
Three Peduncles Convey the Input and Output of Each Half of the Cerebellum
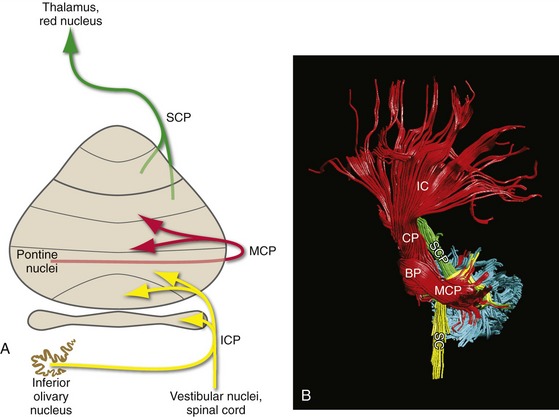
Three peduncles containing the cerebellar afferents and efferents attach the cerebellum to the brainstem (Fig. 20-2). The superior cerebellar peduncle is the major output route from its side of the cerebellum, carrying all the efferents from the dentate and interposed nuclei and some of the efferents from the fastigial nucleus. The middle cerebellar peduncle is the input route for information from the cerebral cortex, carrying the fibers from contralateral pontine nuclei. By elimination then, the inferior cerebellar peduncle is a complex bundle, carrying most of the remaining cerebellar afferents (including climbing fibers, as described a little later), as well as the remaining cerebellar efferents.
All Parts of the Cerebellum Share Common Organizational Principles
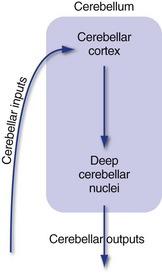
All parts of the cerebellum have a cortex with the same structure and use the same basic circuitry (Fig. 20-3; inputs → cortex → deep nuclei → outputs). This suggests that all parts of the cerebellum perform the same basic (still mysterious) operation, and that the functional differences among different cerebellar regions are simply reflections of different input sources and output targets.
Inputs Reach the Cerebellar Cortex as Mossy and Climbing Fibers
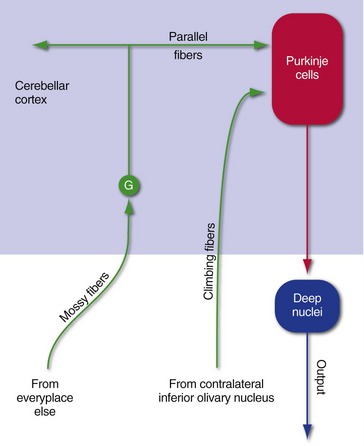
Afferents reach the cerebellar cortex in two forms: mossy fibers and climbing fibers (Fig. 20-4). Climbing fibers, all from the contralateral inferior olivary nucleus, end directly on Purkinje cells, which provide the output from cerebellar cortex. Mossy fibers, in contrast, come from many other places and end on the tiny granule cells of the cerebellar cortex; these in turn issue parallel fibers that synapse on Purkinje cells. Although mossy fibers arise on both sides of the CNS, those reaching one side of the cerebellum carry information related to the ipsilateral side of the body (e.g., see Figs. 20-6 and 20-7 later in this chapter).
Purkinje Cells of the Cerebellar Cortex Project to the Deep Nuclei
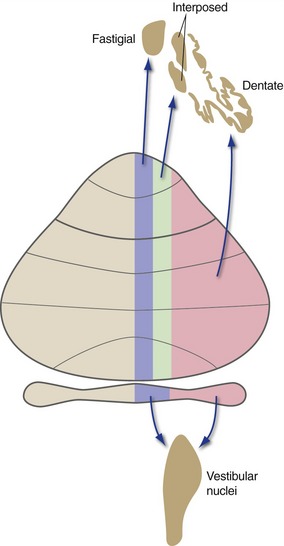
The flocculonodular lobe is primarily concerned with vestibular function (posture and eye movements), and most of its output is directed to the vestibular nuclei either directly or indirectly. The three longitudinal zones of the remainder of the cerebellum direct their outputs to three deep nuclei arranged in a corresponding medial-to-lateral array (Fig. 20-5): vermis → fastigial nucleus, medial hemisphere → interposed (globose + emboliform) nucleus, and lateral hemisphere → dentate nucleus.
Details of Connections Differ Among Zones
Each functional zone of the cerebellum is connected not only with a particular output nucleus, but also with a distinctive set of inputs (Fig. 20-6). The flocculonodular lobe primarily receives vestibular afferents. The vermis and medial hemispheres receive spinocerebellar inputs and inputs from motor cortex (via pontine nuclei). The lateral hemispheres receive inputs via the pontine nuclei from widespread cortical areas. (These are not the only inputs to these zones, but they do provide a hint about the principal function of each zone.)
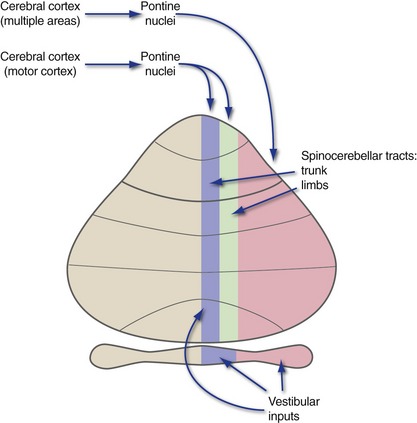
Cerebellar Cortex Receives Inputs from Multiple Sources
The Spinal Cord Projects to the Vermis and Medial Hemisphere
The vermis and particularly the medial hemisphere receive overlapping mossy fiber inputs from the ipsilateral spinal cord and trigeminal nuclei (hence, the ipsilateral side of the body) and the contralateral cerebral cortex (hence, the ipsilateral side of the body once again), in a superimposed, roughly somatotopic pattern (THB6 Figure 20-17, p. 510). Both spinal and cortical (mainly from motor cortex) inputs representing the limbs reach the medial hemisphere; those representing the trunk reach the vermis.
Each Longitudinal Zone Has a Distinctive Output
Each functional zone of cerebellar cortex has not only a distinguishing set of inputs (see Fig. 20-6), but also a distinguishing pattern of projections to the deep nuclei (see Fig. 20-5). The deep nuclei in turn have further projections that make functional sense.
The fastigial nucleus projects through the inferior cerebellar peduncle mainly to the vestibular nuclei and reticular formation. The interposed and dentate nuclei project through the superior cerebellar peduncle, most of whose fibers cross the midline in the caudal midbrain, pass through or around the red nucleus, and reach VL/VA of the thalamus (Fig. 20-7). Some fibers from the interposed nucleus end in the part of the red nucleus that gives rise to the rubrospinal tract; some from the dentate nucleus reach the (much larger) part of the red nucleus that in turn projects to the inferior olivary nucleus.
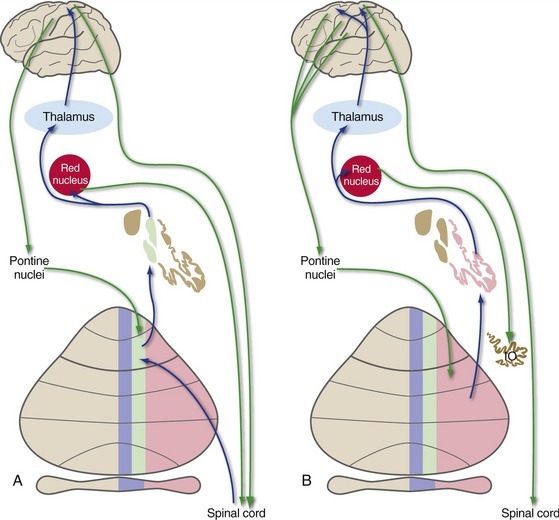
Patterns of Connections Indicate the Functions of Longitudinal Zones
The Vermis Is Involved in Postural Adjustments
Vermal inputs, from the vestibular nuclei and from spinocerebellar fibers representing the trunk, are concerned with the general orientation and posture of the body. Similarly, vermal outputs affect vestibulospinal and reticulospinal neurons that are related to antigravity and other proximal muscles, suggesting a role in adjusting posture and motor programs (e.g., walking).
The Flocculus and Vermis Are Involved in Eye Movements
As mentioned in Chapter 14 and again in Chapter 21, the vestibular system plays a major role in vestibuloocular and other kinds of smooth eye movements. The vestibular connections of the flocculus make it another important player in smooth eye movements (see Chapter 21). In addition, the vestibular/visual connections of the vermis enable it to play a role in the coordination of gaze shifts (see Chapter 21), just as the rest of the cerebellum is important in the coordination of movements generally.
Clinical Syndromes Correspond to Functional Zones
In real life, partial cerebellar damage is likely to affect either the vermis, one cerebellar hemisphere, or the flocculonodular lobe. Vermal damage causes postural instability and a broad-based, staggering gait. Damage to one hemisphere causes incoordination (ataxia) of the ipsilateral limbs, in part because of deficits in planning movements (lateral hemisphere) and in part because of deficits in correcting movements already under way (medial hemisphere). Flocculonodular damage also causes a broad-based, staggering gait and affects eye movements as well (see Chapter 21).