[level-membership-for-neurology-category]Chapter 10
Brain stem, cranial nerves, visual system, vestibular system, reticular system
Brain stem
Basic anatomy
Mid-brain
The mid-brain is approximately 1.5 cm long and extends from the pons to the mamillary bodies. The ventral surface connects to the two superior cerebellar peduncles (S2.12). The roof of the mid-brain is composed of four colliculi, the superior colliculi associated with the visual system and the inferior colliculi related to the auditory system. The red nucleus is also contained within the mid-brain. The periaqueductal grey, an area of grey matter within the mid-brain, is important in the descending modulation of pain (S3.29).
Medulla oblongata
The medulla is about 3 cm long and forms the base of the brain stem, adjoining the spinal cord at the level of foramen magnum. The descending motor tracts passing through it form two pyramids on the ventral surface of the medulla between which is the anterior median fissure. This fissure is disrupted where the tracts cross over the midline, termed the ‘decussation of the pyramids’. The inferior cerebellar peduncle attaches to the medulla and behind it is the ‘olive’. The olive is a prominent oval swelling that marks the position of the inferior olivary nucleus (S2.12).
Function of the brain stem
The three regions of the brain stem contain the nuclei of the cranial nerves which control a variety of vital functions (S2.10). The ascending (S2.15) and descending tracts (S2.14) also pass through the brain stem between the spinal cord and cerebral cortex. The reticular formation (S2.10) and the cerebellum (S2.12) are integrally linked with all three regions and have specialist roles related to movement.
Cranial nerves
Basic anatomy
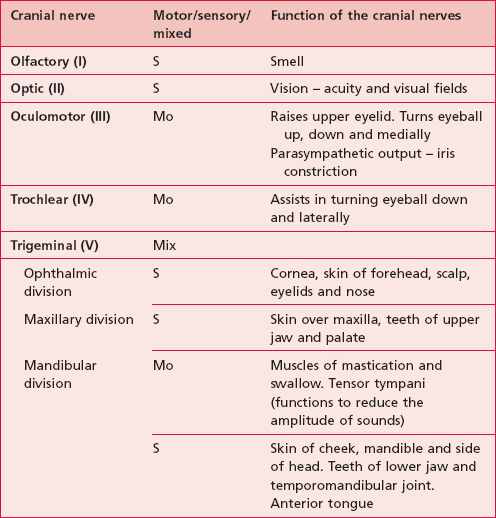
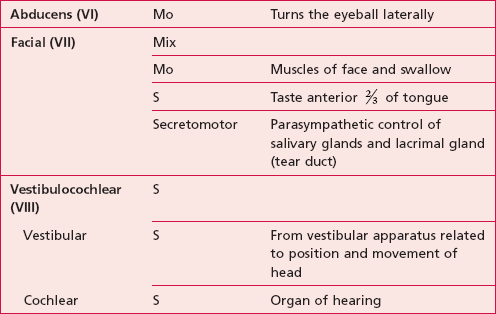
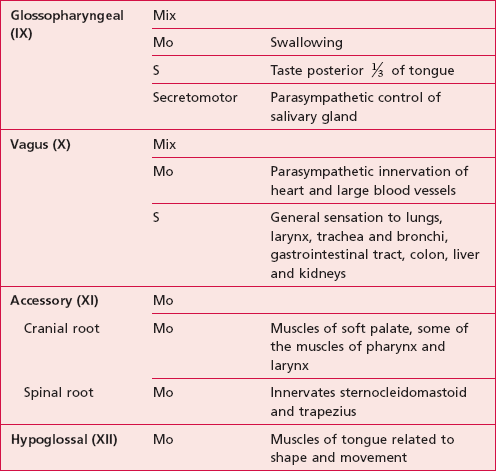
The names, numbers and functions of the cranial nerves can be seen in Table 10.1.
Motor nuclei
Five of the cranial nerves are entirely motor. Afferent input to the motor nuclei is from the cerebral cortex via the cortico-bulbar tract (S2.14). The output from the cranial nerve nuclei to the effector muscle is via a lower motor neuron.
Assessment of the cranial nerves
Traditionally, this has been carried out by the medical team on initial contact and is not covered in this text. However, it is important for the therapist to understand the findings and to identify any changes that may occur. Symptoms associated with cranial deficits may be identified by the patient themselves during the subjective assessment (S3.16) or noticed by the therapist by general observation (S3.17). In either case, any suspicions should be followed up with a referral to an experienced clinician or the medical team.
Visual system
Basic anatomy
The visual pathway
The sensory receptors of the visual system are the photosensitive rod and cone proteins of the retina. The rods and cones are most dense in the central region of the retina called the fovea. An image of the environment is focused onto the retina by alterations in the thickness of the lens. The retina converts a light pattern into a neuronal signal, which is conducted along the visual pathway to various parts of the brain to be processed (Fig. 10.1).
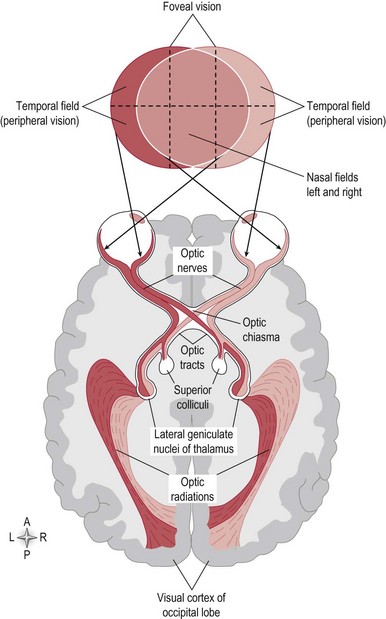
Figure 10.1 The visual pathway.
Function of the visual system
Visual acuity
Scanning (S3.27)
The ability to scan the eyes and head in a coordinated manner to maintain the object of interest or visual stimuli in the field of vision, e.g. while reading. The relevant head/neck movements are achieved via the tectospinal and tectobulbar tracts (S2.15) and eye movements by the cranial nerve nucleui (III and VI).
Visual fields (S3.27)
Information from the temporal and nasal visual fields of both eyes reaches the occipital cortex simultaneously, via the visual pathway (Fig. 10.1).
Vision and movement
The visual system plays an important part in movement and balance as it provides us with:
Vestibular system
Basic anatomy
Input to the vestibular system
Cochlear: which is associated with hearing and will not be discussed further here
Vestibule
The vestibule consists of two organs, the utricle and saccule. In the wall of the utricle and saccule is a thickened region of specialized epithelium, called a ‘macula’ (Fig. 10.2).
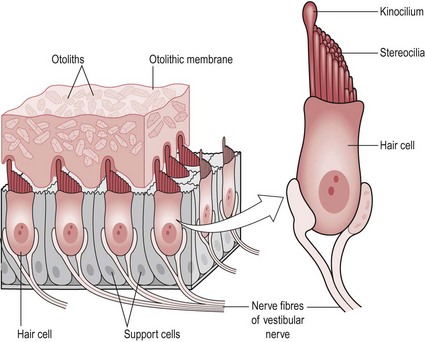
Figure 10.2 The structure of a macula.
The macula consists of two types of cells:
Supporting cells: Each supporting cell has a sensory neuron connection (vestibular branch of the vestibulocochlear nerve VIII). These cells secrete a thick gelatinous layer called the ‘otolithic membrane’ on top of which is a dense layer of calcium carbonate crystals called ‘otoliths’. This characteristic is highly relevant to the receptors function.
Hair cell bundles (Fig. 10.2): Each supporting cell holds a bundle of hair cells, stereocilia (40–80/bundle) and kinocilium (1/bundle). Each stereocilia is physically connected to the kinocilium so that when it is moved, mechanically gated ion channels are opened allowing the movement of ions. This creates a graded potential in the sensory neuron. The cell is hyperpolarized when the stereocilia moves away from the kinocilium and depolarized with movement in the opposite direction.
Semicircular canals
There are three semicircular canals which lie at 90° to each other. At the base of each canal is a thickened region termed the ‘ampulla’. Inside the ampulla is an elevation of specialized cells called the ‘crista’ (Fig. 10.3).
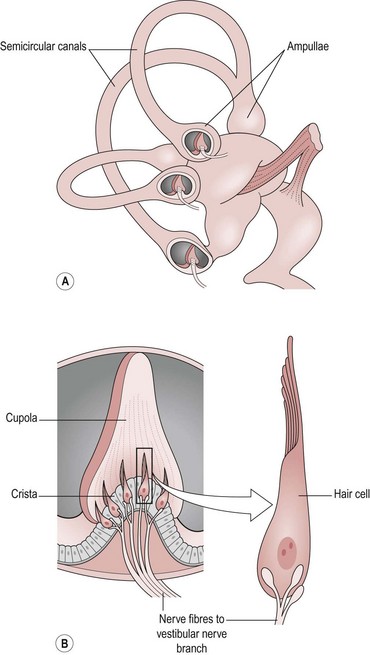
Figure 10.3 (A,B) The structure of a crista.
The crista consists of two types of cells:
Supporting cells: The supporting cells are similar in structure to those of the macula. They produce a gel which overlies the hair bundles like a cap. This is termed the ‘cupola’ and it is much deeper than the otolithic membrane of the macula so that it covers the full width of the ampulla. This means that the endolymph within the membranous labyrinth cannot pass the cupola and it is this feature that is highly relevant to its function.
Hair cell bundles: The hair bundles are in the same arrangement as for the macula and are all orientated in the same direction within one canal. However, each canal has a different orientation so that the semicircular canals can respond to movement in any plane. The orientation of the left and right sides of the brain is also different with each contralateral equivalent aligned in the opposite direction. The relevance of this will be explained below.
Output from the vestibular system
Cerebellum (S2.12) for processing and execution of smooth co-ordinated movement
Cranial nerves (III and VI) to initiate the vestibulo-ocular reflex
Descending tracts to initiate the appropriate muscle response. The muscle activation via the vestibulospinal tract will be specific to need but involving the neck, proximal limb and trunk muscles (lateral and medial vestibulospinal tract).
Function of the vestibular system
Vestibule
For example, during a forward head tilt (cervical spine flexion), the macula of the saccule will respond. The forward tilt causes the heavy mass of the otoliths and the otolithic membrane to ‘fall’ forwards moving the stereocilia in relation to the kinocilium and producing a graded potential in the sensory neuron. As the orientation of hair cells is opposite within each half of the saccule, the result will be depolarization (excitation) in one-half and hyperpolarization (inhibition) in the other (S2.6).
The action potential produced travels along the vestibular nerve (VIII) and recruits or inhibits the appropriate muscles via the vestibulospinal tract (S2.15). In this case, the forward head tilt has moved the centre of gravity forwards making the body less stable. The balance reaction required will attempt to bring the centre of gravity backwards, by recruiting extensor muscles around the pelvis/hip or trunk and simultaneously reduce the activity of the flexor muscles. This is important so that the centre of gravity is not moved further forwards and balance becomes seriously compromised. Remember also that the orientation of hair bundles in one-half of the saccule is the same in both the left and right sides of the brain and therefore the response will be bilateral. This makes sense, as the response to a linear movement needs to be symmetrical.
Reticular system
Basic anatomy
• The median column consisting of intermediate sized neurons
• The medial column containing large neurons (the median and medial columns are also known as the raphe nuclei)
Function of the reticular formation
Modulation of muscle tone
The appropriate level of resting muscle tone is regulated by modulation of the stretch reflex (S2.13) by the descending tracts. The reticulospinal tracts in combination with the vestibulospinal and reticulobulbar tracts influence muscle tone at the level of the alpha motor neuron. The two reticulospinal tracts specifically influence extensor muscle tone. The pontine/lateral reticulospinal tract terminates at the ventral horn directly on motor neurons and enhances extensor muscle tone, whereas the medullary/medial reticulospinal tract (S2.14) terminates indirectly on motor neurons and exerts an inhibitory influence on extensor muscle tone. The antagonistic effect of these tracts allows modulation and grading to set the level of extensor muscle tone appropriately for the functional need. However, the reticulospinal tracts also work in cooperation with the vestibulospinal tracts (S2.14) to specifically maintain tone of the antigravity muscles.
The reticular formation also has a role in the motor control of facial expression related to our emotional behaviour (S2.9) via its links to the limbic system and coordination of the timing and direction of eye movements via the superior colliculus and cranial nerves.
References and Further Reading
Hunziker, H-W. In the eye of the reader: foveal and peripheral perception – from letter recognition to the joy of reading. Zürich: Transmedia Stäubli Verlag; 2006.
Palmer, SM, Rosa, MG. A distinct anatomical network of cortical areas for analysis of motion in far peripheral vision. European Journal of Neuroscience. 2006; 248:2389–2405.
Purves, D, Augustine, GJ, Fitzpatrick, D, et al. Neuroscience, ed 4. Sunderland: Sinauer Associates; 2008.
Snell, RS. Clinical neuroanatomy, ed 6. Philadelphia: Lippincott Williams and Wilkins; 2006.
[/level-membership-for-neurology-category][not-level-membership-for-neurology-category]Chapter 10
Brain stem, cranial nerves, visual system, vestibular system, reticular system
Brain stem
Basic anatomy
Mid-brain
The mid-brain is approximately 1.5 cm long and extends from the pons to the mamillary bodies. The ventral surface connects to the two superior cerebellar peduncles (S2.12). The roof of the mid-brain is composed of four colliculi, the superior colliculi associated with the visual system and the inferior colliculi related to the auditory system. The red nucleus is also contained within the mid-brain. The periaqueductal grey, an area of grey matter within the mid-brain, is important in the descending modulation of pain (S3.29).
Medulla oblongata
The medulla is about 3 cm long and forms the base of the brain stem, adjoining the spinal cord at the level of foramen magnum. The descending motor tracts passing through it form two pyramids on the ventral surface of the medulla between which is the anterior median fissure. This fissure is disrupted where the tracts cross over the midline, termed the ‘decussation of the pyramids’. The inferior cerebellar peduncle attaches to the medulla and behind it is the ‘olive’. The olive is a prominent oval swelling that marks the position of the inferior olivary nucleus (S2.12).
Function of the brain stem
The three regions of the brain stem contain the nuclei of the cranial nerves which control a variety of vital functions (S2.10). The ascending (S2.15) and descending tracts (S2.14) also pass through the brain stem between the spinal cord and cerebral cortex. The reticular formation (S2.10) and the cerebellum (S2.12) are integrally linked with all three regions and have specialist roles related to movement.
Cranial nerves
Basic anatomy
The names, numbers and functions of the cranial nerves can be seen in Table 10.1.
Motor nuclei
Five of the cranial nerves are entirely motor. Afferent input to the motor nuclei is from the cerebral cortex via the cortico-bulbar tract (S2.14). The output from the cranial nerve nuclei to the effector muscle is via a lower motor neuron.
Assessment of the cranial nerves
Traditionally, this has been carried out by the medical team on initial contact and is not covered in this text. However, it is important for the therapist to understand the findings and to identify any changes that may occur. Symptoms associated with cranial deficits may be identified by the patient themselves during the subjective assessment (S3.16) or noticed by the therapist by general observation (S3.17). In either case, any suspicions should be followed up with a referral to an experienced clinician or the medical team.
