[level-membership-for-basic-science-category]
Chapter 14 Basal Ganglia
The term basal ganglia denotes a number of subcortical nuclear masses that lie in the inferior part of the cerebral hemisphere, lateral to the thalamus (Figs 14.1, 14.2). They have traditionally been regarded as including the corpus striatum, the claustrum and the amygdaloid complex. More recently, however, the working definition has been narrowed to cover only the corpus striatum and its associated structures in the diencephalon and midbrain. The reasoning behind this change is that these structures form a functional complex involved in the control of movement and motivational aspects of behaviour, whereas the claustrum is of unknown function and the amygdala is more closely related to the limbic system (Ch. 16).
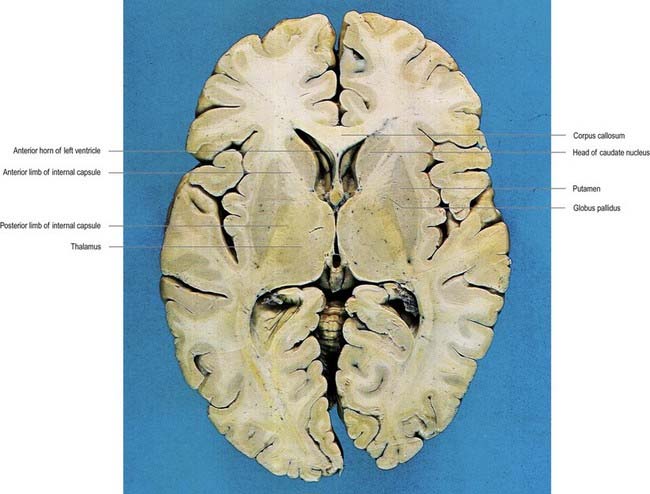
Fig. 14.1 Horizontal section of the brain.
(Dissection by E. L. Rees; photograph by Kevin Fitzpatrick on behalf of GKT School of Medicine, London.)
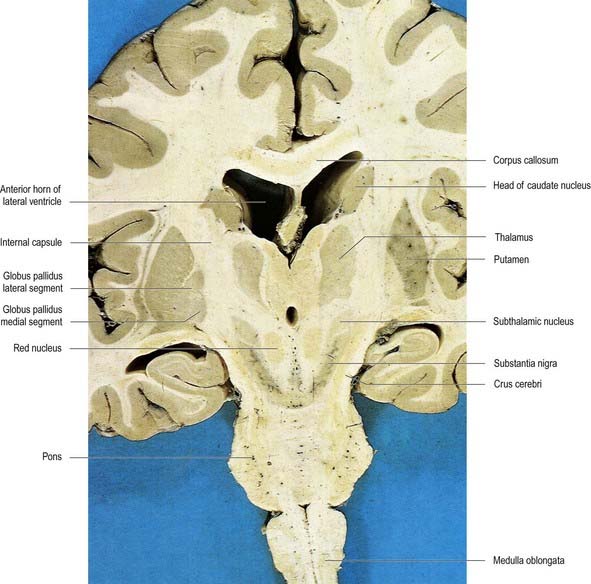
Fig. 14.2 Oblique coronal section of the brain.
(Dissection by E. L. Rees; photograph by Kevin Fitzpatrick on behalf of GKT School of Medicine, London.)
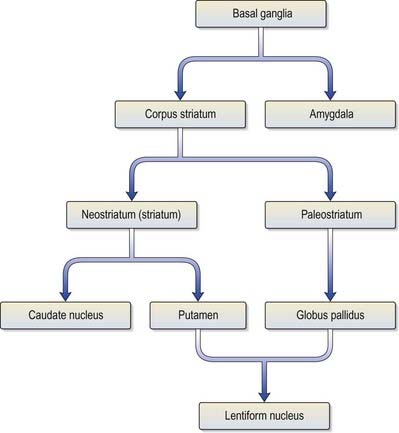
The corpus striatum consists of the caudate nucleus, putamen and globus pallidus (Fig. 14.3). Because of their close proximity, the putamen and globus pallidus were historically considered a single entity, termed the lentiform complex or nucleus. With increasing knowledge of their structure and function, however, it has become clear that the putamen is more correctly considered to be in unity with the caudate nucleus, with which it shares common chemocytoarchitecture and connections. The putamen and caudate nucleus are together referred to as the neostriatum or simply the striatum.
The striatum is considered the principal ‘input’ structure of the basal ganglia because it receives the majority of afferents from other parts of the neuraxis. Its principal efferent connections are to the globus pallidus and pars reticulata of the substantia nigra. The globus pallidus—in particular, its medial segment—together with the pars reticulata of the substantia nigra (Ch. 10) is regarded as the main ‘output’ structure because it is the source of massive efferent fibre projections, mostly directed to the thalamus.
The caudate nucleus is a curved, tadpole-shaped mass. It has a large anterior head, which tapers to a body, and a down-curving tail (Fig. 14.4). The head is covered with ependyma and lies in the floor and lateral wall of the anterior horn of the lateral ventricle, in front of the interventricular foramen. The tapering body is in the floor of the body of the ventricle, and the narrow tail follows the curve of the inferior horn, lying in the ventricular roof in the temporal lobe. Medially, the greater part of the caudate nucleus abuts the thalamus, along a junction marked by a groove, the sulcus terminalis. The sulcus contains the stria terminalis, lying deep to the ependyma (see Fig. 5.3; Fig. 14.5). The stria terminalis forms one margin of the choroid fissure of the lateral ventricle; the hippocampal fimbria and fornix form the other margin. The sulcus terminalis is especially prominent anterosuperiorly (because of the large head and body of the caudate nucleus relative to the tail), where it is accompanied by the thalamostriate vein.
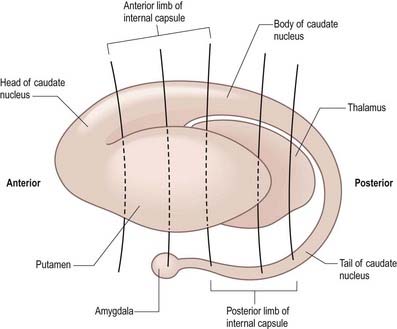
Fig. 14.4 Striatum within the left cerebral hemisphere.
(By permission from Crossman, A.R., Neary, D., 2000. Neuroanatomy, 2nd ed. Churchill Livingstone, Edinburgh.)
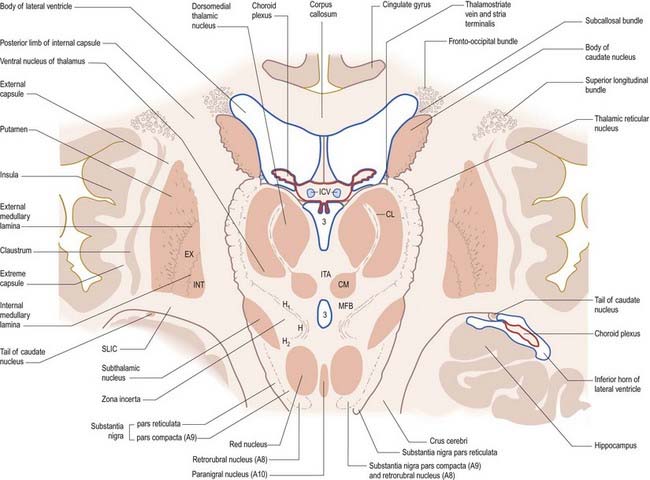
Fig. 14.5 Oblique section through the diencephalon and basal ganglia. A8, A9, A10, dopaminergic cell groups; CL, centrolateral nucleus of thalamus; CM, centromedian nucleus of thalamus; EX, external pallidal segment; H, H1, H2, subthalamic fields of Forel; ICV, internal cerebral veins in the transverse fissure; INT, internal pallidal segment; ITA, interthalamic adhesion; MFB, median forebrain bundle; SLIC, sublentiform internal capsule; 3, third ventricle.
The corpus callosum lies above the head and body of the caudate nucleus. The two are separated laterally by the fronto-occipital bundle and medially by the subcallosal fasciculus, a bundle of axons that caps the nucleus (see Fig. 14.5; Figs 14.6, 14.7). The caudate nucleus is largely separated from the lentiform complex by the anterior limb of the internal capsule (Figs 14.1, 14.6, 14.7). However, the inferior part of the head of the caudate becomes continuous with the most inferior part of the putamen immediately above the anterior perforated substance. This junctional region is sometimes known as the fundus striati (see Fig. 14.6). Variable bridges of cells connect the putamen to the caudate nucleus for most of its length. They are most prominent anteriorly, in the region of the fundus striati and the head and body of the caudate nucleus, where they break up the anterior limb of the internal capsule (see Figs 14.6, 14.7). In the temporal lobe, the anterior part of the tail of the caudate nucleus becomes continuous with the posteroinferior part of the putamen. The vast bulk of the caudate nucleus and putamen are often referred to as the dorsal striatum. A smaller inferomedial part of the rostral striatum is referred to as the ventral striatum and includes the nucleus accumbens.
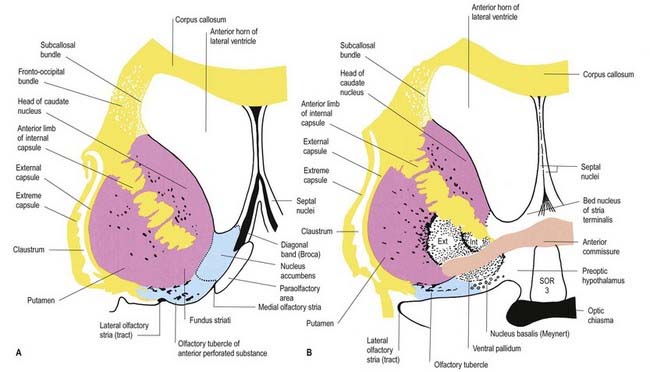
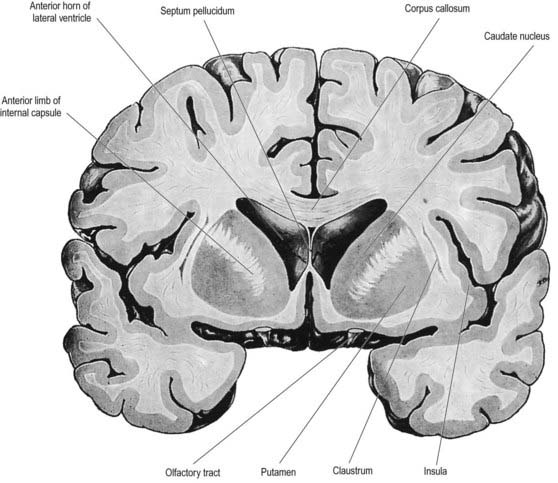
Fig. 14.7 Posterior aspect of a coronal section through the anterior horn of the lateral ventricles.
The lentiform complex lies deep to the insular cortex, with which it is roughly coextensive, although the two are separated by a thin layer of white matter and the claustrum (see Fig. 14.2; Fig. 14.8). The claustrum splits the insular subcortical white matter to create the extreme and external capsules. The latter separates the claustrum from the putamen (see Figs 16.40, 14.1, 14.2, 14.8). The lentiform complex is separated from the caudate nucleus by the internal capsule.
Inferiorly, a little behind the fundus striati, the lentiform complex is grooved by the anterior commissure, which connects inferior parts of the temporal lobes and the anterior olfactory cortex of the two sides (see Fig. 14.6). The area above the commissure is referred to as the dorsal pallidum, and that below it is the ventral pallidum.
Striatum
Neuroactive chemicals, whether intrinsic or derived from afferents, are not distributed uniformly in the striatum. For example, serotonin and glutamic acid decarboxylase concentrations are highest caudally, whereas substance P, acetylcholine and dopamine are highest rostrally. However, there is a finer-grain neurochemical organization that informs the view of the striatum as a mosaic of islands or striosomes (sometimes referred to as patches), each 0.5 to 1.5 mm across, packed into a background matrix. Striosomes contain substance P and enkephalin. During development, the first dopamine terminals from the substantia nigra are found in striosomes. Although this exclusivity does not persist after birth, striosomes in the caudate nucleus still contain a higher concentration of dopamine than the matrix does. The latter contains acetylcholine and somatostatin and is the target of thalamostriate axons. Receptors for at least some neurotransmitters are also differentially distributed. For example, opiate receptors are found almost exclusively within striosomes, and muscarinic receptors predominantly so. Moreover, the distribution of neuroactive substances within the striosomes is not uniform. In humans, the striosome–matrix patchwork is less evident in the putamen, where it appears to consist predominantly of matrix, than it is in the caudate nucleus.
The major connections of the striatum are summarized in Figure 14.9. Although the connections of the dorsal and ventral divisions overlap, a generalization can be made: the dorsal striatum is predominantly connected with motor and associative areas of the cerebral cortex, whereas the ventral striatum is connected with the limbic system and orbitofrontal and temporal cortices. For both dorsal and ventral striata, the pallidum and substantia nigra pars reticulata are key efferent structures. The fundamental arrangement is the same for both divisions. The cerebral cortex projects to the striatum, which in turn projects to the pallidum and substantia nigra pars reticulata. From these, efferents leave to influence the cerebral cortex (either the supplementary motor area or the prefrontal and cingulate cortices via the thalamus) and the superior colliculus.
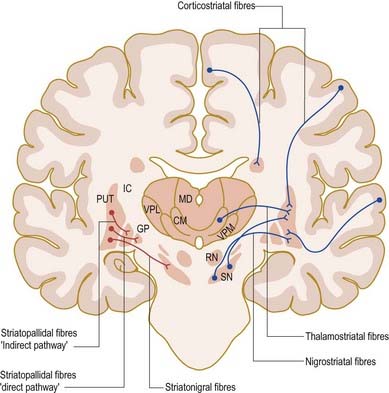
The striatum also receives afferents, which are more crudely spatially organized, from the polysensory intralaminar thalamus. The cerebelloreceptive nucleus centralis lateralis projects to the anterior striatum (especially the caudate nucleus), and the cerebello- and pallidoreceptive centromedian nucleus projects to the putamen.
The aminergic inputs to the caudate and putamen are derived from the substantia nigra pars compacta (dopaminergic group A9; Figs 14.10, 14.11), the retrorubral nucleus (dopaminergic group A8; see Fig. 14.5), the dorsal raphe nucleus (serotoninergic group B7; see Fig. 14.10) and the locus coeruleus (noradrenergic group A6). The nigrostriatal input is sometimes referred to as the ‘mesostriatal’ dopamine pathway. It reaches the striatum by traversing the H fields of the subthalamus. These aminergic inputs appear to modulate the responses of the striatum to cortical and thalamic afferent influences.
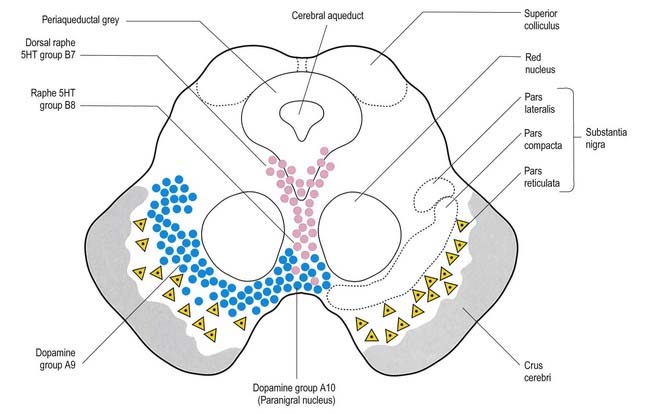
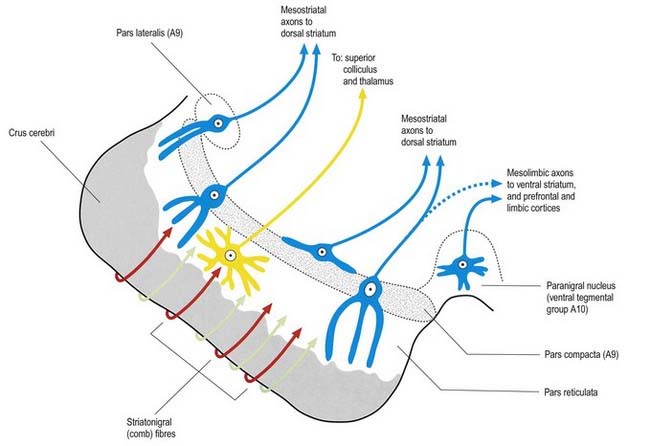
Fig. 14.11 Scheme of the organization of the substantia nigra in transverse section. Compare with Figure 14.10. Medially, there is no sharp distinction between dopaminergic cells projecting to the dorsal striatum (pars compacta, A9) and those projecting to the ventral striatum and limbic system (paranigral nucleus, A10). Dendrites of dopaminergic neurones intrude into the pars reticulata. Note the distinctive projection systems from the pars reticulata.
(From Paxinos (Ed.), 1990. The Human Nervous System, vol. 1, Webster. pp. 889–944. Modified with permission from Elsevier.)
Efferents from the striatum pass to both segments of the globus pallidus and to the substantia nigra pars reticulata, where they end in a topically ordered fashion. Fibres ending in either the lateral or medial pallidal segment originate from different striatal cells (Figs 14.9, 14.12). Those to the lateral pallidum come from neurones that co-localize GABA and enkephalin and give rise to the so-called indirect pathway. This name refers to the fact that these striatal neurones influence the activity of basal ganglia output neurones in the medial pallidum via the intermediary of the subthalamic nucleus. Other striatal neurones, which co-localize GABA and substance P/dynorphin, project directly to the medial pallidum and are therefore described as the direct pathway.
A second outflow is established from the striatum to the pars reticulata of the substantia nigra. This also has both direct and indirect components, via the lateral pallidum and subthalamic nucleus (Fig. 14.13). The axons of the direct striatonigral projection constitute the laterally placed ‘comb’ system, which is spatially quite distinct from the ascending dopaminergic nigrostriatal pathway. Striatonigral fibres end in a spatially ordered way in the pars reticulata.
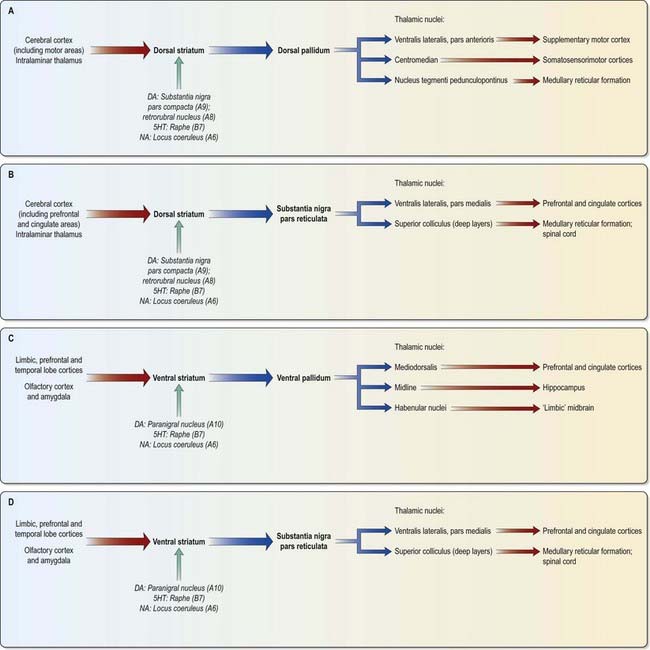
The ventral striatum is the primary target of cortical afferents from limbic cortices, including allocortex, and from limbic-associated regions (see Fig. 14.13). Thus, the hippocampus (through the fornix) and the orbitofrontal cortex (through the internal capsule) project to the nucleus accumbens, and the olfactory, entorhinal, anterior cingulate and temporal visual cortices project to both the nucleus accumbens and the olfactory tubercle in varying degrees. The tubercle also receives afferents from the amygdala. The contiguities of the cortical areas, which project to the ventral striatum and neighbouring dorsal striatum, emphasize the imprecise nature of the boundaries between the two divisions. All the cortical regions overlap and abut one another, and they project to neighbouring parts of the dorsal striatum as well as to the ventral striatum. The fundus striati and ventromedial caudate nucleus abut the olfactory tubercle and nucleus accumbens (see Fig. 14.6) and receive connections from the orbitofrontal cortex and, to a lesser extent, the lateral prefrontal and anterior cingulate cortices (which also project to the contiguous head of the caudate nucleus).
This continuity of the ventral and dorsal striata, as revealed by the arrangements of corticostriate projections, is reinforced by consideration of the aminergic inputs to the ventral striatum. They are derived from the dorsal raphe (serotoninergic group B7), the locus coeruleus (noradrenergic group A6) and the paranigral nucleus (dopamine group A10), as well as from the most medial part of the substantia nigra pars compacta (group A9; see Figs 14.10, 14.11). The dopamine projections constitute the so-called mesolimbic dopamine pathway, which also projects to the septal nuclei, hippocampus, amygdala and prefrontal and cingulate cortices through the medial forebrain bundle. The lateromedial continuity of cell groups A9 and 10 (see Fig. 14.10) is thus reflected in the relative positions of their ascending fibres in the subthalamus and hypothalamus (the H fields and median forebrain bundle, respectively), as well as in the lateromedial topography of the dorsal and ventral striata (see Fig. 14.6), which in turn have contiguous and overlapping sources of cortical afferents.
As for the dorsal striatum, efferents from the ventral striatum project to the pallidum (in this case, the ventral pallidum) and the substantia nigra pars reticulata (Figs 14.13, 14.14). In the latter case, the connection is both direct and indirect via the subthalamic nucleus. The projections from the pars reticulata are as described for the dorsal system, but axons from the ventral pallidum reach the thalamic mediodorsal nucleus (which projects to the cingulate and prefrontal association cortex) and midline nuclei (which project to the hippocampus). Ventral pallidal axons also reach the habenular complex of the limbic system.
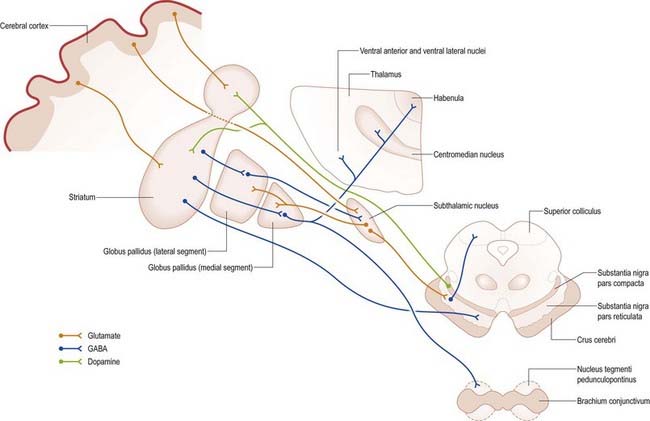
Fig. 14.14 Transverse section through the basal ganglia showing the principal connections of the globus pallidus with the thalamus and subthalamic nucleus.
CASE 1 Huntington’s Disease
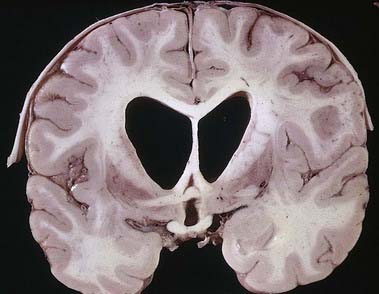
Discussion: This woman has moderately advanced Huntington’s disease (also called Huntington’s chorea), characterized anatomically by progressive atrophy of the caudate nucleus and putamen, with neuronal loss involving especially the medium spiny neurones. The major symptoms of Huntington’s disease are chorea—an irregular, non-repetitive contraction of muscles, often described as ‘dance-like’ movements—and neurocognitive changes, initially psychiatric symptoms and eventually progressing to dementia. Cortical atrophy may be advanced, and ventriculomegaly is at times striking, reflecting both cortical and caudate atrophy (Fig. 14.15). Late in the course of the disease, as the striatum is severely affected, the chorea becomes less obvious and the patient may develop a relatively akinetic state. The disease is well recognized as genetically determined, with autosomal dominant inheritance evident in a great majority of cases, characterized by expanded trinucleotide repeats (CAG) on DNA analysis.
Nucleus Accumbens
The ventral striatum consists of the nucleus accumbens and the olfactory tubercle. In front of the anterior commissure, much of the grey matter of the anterior perforated substance, and especially the olfactory tubercle, is indistinguishable from and continuous with the fundus striati in terms of cellular composition, histochemistry and interconnections. The caudate nucleus is continuous medially with the nucleus accumbens (see Fig. 14.6), which abuts the nuclei of the septum, close to the paraolfactory area, the diagonal band of Broca and the fornix.
Globus Pallidus
Striatopallidal fibres are of two main types. They project to either the lateral or medial pallidal segment. Those projecting to the lateral segment constitute the beginning of the so-called indirect pathway. They use GABA as their primary transmitter and also contain enkephalin. Efferent axons from neurones in the lateral segment pass through the internal capsule in the subthalamic fasciculus and travel to the subthalamic nucleus (see Fig. 15.18).
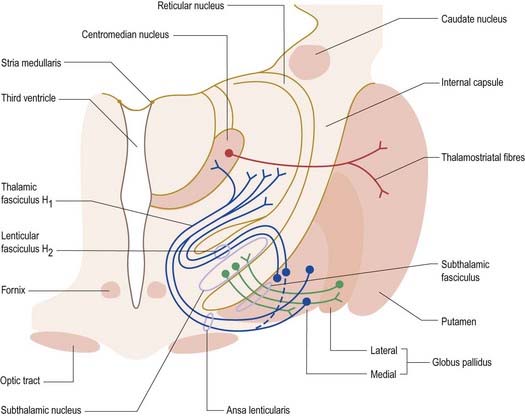
Striatopallidal axons destined for the medial pallidum constitute the so-called direct pathway. Like the indirect projection, these also use GABA as their primary transmitter, but they also contain substance P/dynorphin, rather than enkephalin. Efferent axons from the medial pallidal segment project through the ansa lenticularis and fasciculus lenticularis (see Figs 14.12, 15.18). The former runs around the anterior border of the internal capsule, and the latter penetrates the capsule directly. Having traversed the internal capsule, both pathways unite in the subthalamic region, where they follow a horizontal hairpin trajectory and turn upward to enter the thalamus as the thalamic fasciculus. The trajectory circumnavigates the zona incerta and creates the so-called H fields of Forel (see Figs 14.5, 14.14, 15.18). Within the thalamus, pallidothalamic fibres end in the ventral anterior and ventral lateral nuclei and in the intralaminar centromedian nucleus. These, in turn, project excitatory (presumed glutamatergic) fibres primarily to the frontal cortex, including the primary and supplementary motor areas. The medial pallidum also projects fibres caudally to the pedunculopontine nucleus (see Fig. 14.14). This lies at the junction of the midbrain and the pons, close to the superior cerebellar peduncle, and corresponds approximately to the physiologically identified mesencephalic locomotor region.
CASE 2 Wilson’s Disease
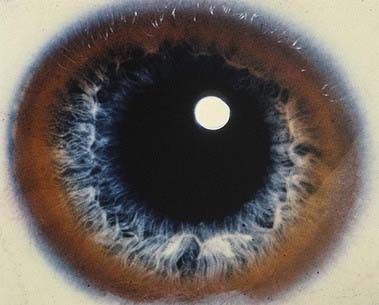
In addition to the neurological abnormalities predicted by these symptoms, ophthalmic examination demonstrates a yellowish brown Kayser–Fleischer ring involving the corneal limbus, representing a deposition of copper in Descemet’s membrane (Fig. 14.16). Imaging demonstrates symmetric ventricular enlargement with widespread atrophic changes, most pronounced in the basal ganglia and thalamus; the putamen is especially involved, with striking vacuolization.
Substantia Nigra
The substantia nigra is a nuclear complex deep to the crus cerebri in each cerebral peduncle of the midbrain. It consists of a pars compacta and a pars reticulata (see Figs 14.10, 14.11). The pars compacta, together with the smaller pars lateralis, corresponds to dopaminergic cell group A9. With the retrorubral nucleus (group A8), it makes up most of the dopaminergic neurone population of the midbrain and is the source of the mesostriatal dopamine system that projects to the striatum. The pars compacta of each side is continuous with its opposite counterpart through the ventral tegmental dopamine group A10, which is sometimes known as the paranigral nucleus. This is the source of the mesolimbic dopamine system, which supplies the ventral striatum and neighbouring parts of the dorsal striatum, as well as the prefrontal and anterior cingulate cortices. The dopaminergic neurones of the pars compacta (group A9) and paranigral nucleus (ventral tegmental group A10) also contain cholecystokinin or somatostatin.
The pars reticulata contains large multipolar cells that are very similar to those of the pallidum. Together they constitute the output neurones of the basal ganglia system. Their disc-like dendritic trees, like those of the pallidum, are oriented at right angles to afferents from the striatum, probably making en passant contacts. Like the striatopallidal axons, of which they may be collaterals, striatonigral axons use GABA and substance P or enkephalin. They distribute differentially in the pars reticulata, such that the enkephalinergic axons terminate in the medial part, whereas substance P axons terminate throughout.
Subthalamic Nucleus
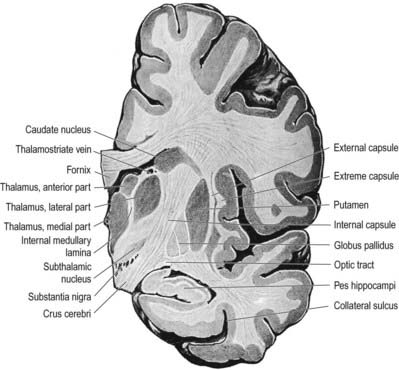
The subthalamic nucleus is a biconvex, lens-shaped nucleus in the subthalamus of the diencephalon. It lies medial to the internal capsule, immediately rostral to the level at which the latter becomes continuous with the crus cerebri of the midbrain (see Figs 14.2, 14.5, 14.8). Within its substance, small interneurones intermingle with large multipolar cells with dendrites, which extend for about one-tenth the diameter of the nucleus. It is encapsulated dorsally by axons, many of which are derived from the subthalamic fasciculus and carry a major GABAergic projection from the lateral segment of the globus pallidus as part of the indirect pathway. It also receives afferents from the cerebral cortex. The subthalamic nucleus is unique in the intrinsic circuitry of the basal ganglia, in that its cells are glutamatergic. They project excitatory axons to both the globus pallidus and the substantia nigra pars reticulata. Within the pallidum, subthalamic efferent fibres end predominantly in the medial segment, but many also end in the lateral segment.
Pathophysiology of Basal Ganglia Disorders
The normal functions of the basal ganglia are difficult to summarize succinctly. As far as their role in movement control is concerned, however, a reasonable definition is that they function to promote and support patterns of behaviour and movement that are appropriate in the prevailing circumstances and to inhibit unwanted or inappropriate behaviour and movements. This is exemplified by disorders of the basal ganglia, which are characterized, depending on the underlying pathology, by an inability to initiate and execute wanted movements (as in Parkinson’s disease) or an inability to prevent unwanted movements (as in Huntington’s disease).
CASE 5 Parkinson’s Disease
Pathological changes in Parkinson’s disease predominate in the zona compacta of the substantia nigra (Fig. 14.17), with loss of melanin-containing neurons and the appearance of intracytoplasmic Lewy bodies, representing an accumulation of the protein synuclein and a resultant degeneration of the nigrostriatal pathway and reduction in striatal dopamine. Anatomical changes may in fact be much more widespread, involving a variety of brain stem nuclei as well as the cerebral cortex.
Dopamine appears to have a dual action on medium spiny striatal neurones. It inhibits those of the indirect pathway and excites those of the direct pathway. Consequently, when dopamine is lost from the striatum, the indirect pathway becomes overactive, and the direct pathway becomes underactive (Fig. 14.18). Overactivity of the striatal projection to the lateral pallidum results in inhibition of pallidosubthalamic neurones and, consequently, overactivity of the subthalamic nucleus. Subthalamic efferents mediate excessive excitatory drive to the medial globus pallidus and substantia nigra pars reticulata. This is exacerbated by underactivity of the GABAergic, inhibitory direct pathway. Overactivity of basal ganglia output then inhibits the motor thalamus and its excitatory thalamocortical connections. Although this description is little more than a first approximation of the underlying pathophysiology, this model of the basis of Parkinsonian symptoms has led to the introduction of new neurosurgical approaches to the treatment of Parkinson’s disease, based on lesioning and deep brain stimulation of the medial globus pallidus and subthalamic nucleus (see later).
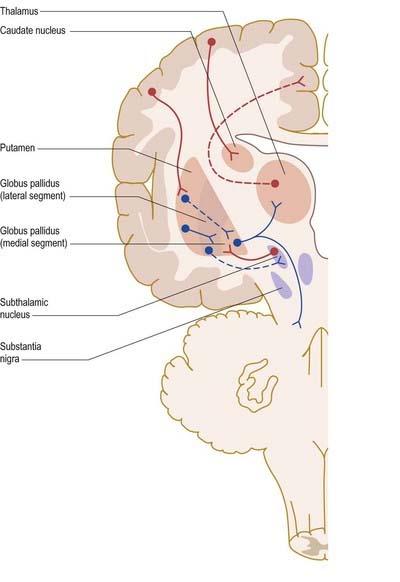
Fig. 14.18 Pathophysiology of Parkinson’s disease. Dotted lines indicate dysfunctional pathways.
(By permission from Crossman, A.R., Neary, D., 2000. Neuroanatomy, 2nd ed. Churchill Livingstone, Edinburgh.)
The current medical treatment for Parkinson’s disease relies on levodopa (L-dopa, L-dihydroxyphenylalanine), the immediate metabolic precursor of dopamine; dopamine agonists; or monoamine oxidase inhibitors. Although these drugs usually provide good symptomatic relief for many years, levodopa and, to a lesser extent, dopamine agonists eventually lead to the development of motor complications, including dyskinesias. The involuntary movements that occur as a consequence of long-term treatment of Parkinson’s disease resemble those seen in Huntington’s disease, tardive dyskinesia and ballism. Experimental evidence suggests that these may share a common neural mechanism (Fig. 14.19). Thus, the indirect pathway becomes underactive, due to the effects of dopaminergic drugs in Parkinson’s disease or the degeneration of the striatopallidal projection to the lateral pallidum in Huntington’s disease. This leads to physiological inhibition of the subthalamic nucleus by overactive pallidosubthalamic neurones. The involvement of the subthalamic nucleus explains why the dyskinetic movements of levodopa-induced dyskinesia and Huntington’s disease resemble those of ballism produced by lesion of the subthalamic nucleus. Underactivity of the subthalamic nucleus removes the excitatory drive from medial pallidal neurones, which are known to be underactive in dyskinesias. Once again, this is an oversimplification. Although it is true that underactivity of the medial globus pallidus is associated with dyskinesias, it is also known that lesions of the globus pallidus alleviate them. This suggests that the dynamic aspects of pallidal and nigral efferent activity are important factors in the generation of dyskinesia.
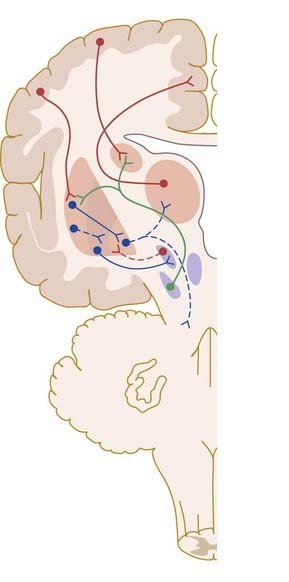
Fig. 14.19 Pathophysiology of dyskinesias. Dotted lines indicate dysfunctional pathways.
(By permission from Crossman, A.R., Neary, D., 2000. Neuroanatomy, 2nd ed. Churchill Livingstone, Edinburgh.)
There is evidence that dysfunction of the basal ganglia is involved in other complex, less well-understood behavioural disorders. In animal experiments, lesions of the basal ganglia, especially of the caudate nucleus, induce uncontrollable hyperactivity (e.g. obstinate progression, incessant pacing and other constantly repeated behaviours). Such evidence has led to the notion that the corpus striatum enables the individual to make motor choices and avoid ‘stimulus-bound’ behaviour. PET studies in humans have shown that sufferers from obsessive-compulsive disorder, which is characterized by repeated ritualistic motor behaviour and intrusive thoughts, exhibit abnormal activity in the prefrontal cortex and caudate nuclei. There have been similar observations in the hyperactive child syndrome. In this respect, it may be significant that the basal ganglia, besides receiving connections from the frontal lobe and limbic cortices, also have an ascending influence on the prefrontal and cingulate cortices through the substantia nigra pars reticulata and dorsomedial and ventromedial thalamus (see Fig. 14.13B–D).
In primates that were made Parkinsonian with the neurotoxin MPTP, lesioning the subthalamic nucleus alleviated tremor, rigidity and bradykinesia. This finding raised the possibility that the subthalamic nucleus could be used as a clinical target. Indeed, lesions of the subthalamic nucleus in humans exert a powerful effect in alleviating tremor, rigidity and bradykinesia; however, the likelihood of side effects is not trivial (the subthalamic nucleus is a small structure wrapped by fibres of passage and close to the hypothalamus and internal capsule).
The implantation of deep brain electrodes to inhibit cells in the vicinity through high-frequency pulses generated by a pacemaker has been a concept since the early 1970s, but it did not become a widespread reality until the late 1980s, as a result of technological advances. The introduction of this technique, which avoids making permanent lesions, made bilateral surgery safer. There have been numerous reports of the effectiveness of both bilateral pallidal and subthalamic nucleus stimulation (Figs 14.20, 14.21). Subthalamic nucleus stimulation is favoured by most groups because, unlike pallidal stimulation, it allows patients to reduce their anti-Parkinsonian medication.
CASE 7 Restless Legs Syndrome
A 50-year-old married woman presents with a history of insomnia, characterized by difficulty falling asleep and multiple nocturnal awakenings. It began 3 years ago and is increasing in severity; whereas it affected her once a week initially, it now affects her nightly. The resulting daytime sleepiness, anergy and fatigue have begun to interfere with her daytime functioning, including decreased memory for recent events, word-finding difficulties, avoidance of social contact, low mood and decreased job performance. Upon further questioning, she reports an irresistible urge to move both lower extremities beginning at 9 PM and intensifying significantly just after retiring. These are associated with a ‘creepy crawly’ sensation in her lower extremities, beginning with her calf muscles and generalizing into her hips and abdomen and sometimes into her arms and neck. She obtains temporary relief by stretching her extremities vigorously, rubbing them, using warm soaks or getting out of bed and walking around the house; however, the symptoms return shortly thereafter and awaken her repeatedly after sleep onset. She recalls similar symptoms, transiently, during her last pregnancy. Past medical history is negative, and she is taking no medications. She consumes six caffeinated beverages a day, the last one at 6 PM. Serum tests reveal a low ferritin level of 10 µg/l, yet there is no evidence of anemia. Polysomnography reveals high proportions of stage 1 sleep, diminished proportions of slow-wave sleep, multiple awakenings and arousals and repetitive bursts of electromyographic activity on lower limb leads, each lasting approximately 0.75 second, followed by brief arousal and separated by intervals of 20 seconds.
Alexander, DeLong, Strick, 1986Alexander G.E. DeLong M.R. Strick P.L. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu. Rev. Neurosci.. 9:1986;357-382.
Benarroch E.E. Subthalamic nucleus and its connections: anatomic substrate for the network effects of deep brain stimulation. Neurology. 2008;70:1991-1994.
Braak H., Del Tredici K. Nervous system pathology in sporadic Parkinson disease. Neurology. 2008;70:1916-1924.
Crossmann A.R. A hypothesis on the pathophysiological mechanisms that underlie levodopa- or dopamine agonist-induced dyskinesia in Parkinson’s disease: implications for future strategies in treatment. Mov. Disord.. 1990;5:100-108.
Krack et al, 2003Krack P. Batir A. Van Blercom N. et al. Five-year follow-up of bilateral stimulation of the subthalamic nucleus in advanced Parkinson’s disease. N. Engl. J. Med.. 349:2003;1925-1934.
Laitinen, Bergenheim, Hariz, 1992Laitinen L.V. Bergenheim A.T. Hariz M.I. Ventroposterolateral pallidotomy can abolish all Parkinsonian symptoms. Stereotact. Funct. Neurosurg.. 58:1992;14-21.
Key paper that ignited widespread interest in functional neurosurgery for Parkinson’s disease.
Obeso J.A., Marin C., Rodriguez-Oroz C., Blesa J., Benitiez-Temino B., Mena-Segovia J., et al. The basal ganglia in Parkinson’s disease: current concepts and unexplained observations. Ann. Neurol.. 2008;64(Suppl):S30-S46.
Penney, Young, 1986Penney J.B. Jr., Young A.B.: Striatal inhomogeneities and basal ganglia function. Mov. Disord.. 1:1986;3-15.
[/level-membership-for-basic-science-category][not-level-membership-for-basic-science-category]
Chapter 14 Basal Ganglia
The term basal ganglia denotes a number of subcortical nuclear masses that lie in the inferior part of the cerebral hemisphere, lateral to the thalamus (Figs 14.1, 14.2). They have traditionally been regarded as including the corpus striatum, the claustrum and the amygdaloid complex. More recently, however, the working definition has been narrowed to cover only the corpus striatum and its associated structures in the diencephalon and midbrain. The reasoning behind this change is that these structures form a functional complex involved in the control of movement and motivational aspects of behaviour, whereas the claustrum is of unknown function and the amygdala is more closely related to the limbic system (Ch. 16).
Fig. 14.1 Horizontal section of the brain.
(Dissection by E. L. Rees; photograph by Kevin Fitzpatrick on behalf of GKT School of Medicine, London.)
Fig. 14.2 Oblique coronal section of the brain.
(Dissection by E. L. Rees; photograph by Kevin Fitzpatrick on behalf of GKT School of Medicine, London.)
The corpus striatum consists of the caudate nucleus, putamen and globus pallidus (Fig. 14.3). Because of their close proximity, the putamen and globus pallidus were historically considered a single entity, termed the lentiform complex or nucleus. With increasing knowledge of their structure and function, however, it has become clear that the putamen is more correctly considered to be in unity with the caudate nucleus, with which it shares common chemocytoarchitecture and connections. The putamen and caudate nucleus are together referred to as the neostriatum or simply the striatum.
The striatum is considered the principal ‘input’ structure of the basal ganglia because it receives the majority of afferents from other parts of the neuraxis. Its principal efferent connections are to the globus pallidus and pars reticulata of the substantia nigra. The globus pallidus—in particular, its medial segment—together with the pars reticulata of the substantia nigra (Ch. 10) is regarded as the main ‘output’ structure because it is the source of massive efferent fibre projections, mostly directed to the thalamus.
The caudate nucleus is a curved, tadpole-shaped mass. It has a large anterior head, which tapers to a body, and a down-curving tail (Fig. 14.4). The head is covered with ependyma and lies in the floor and lateral wall of the anterior horn of the lateral ventricle, in front of the interventricular foramen. The tapering body is in the floor of the body of the ventricle, and the narrow tail follows the curve of the inferior horn, lying in the ventricular roof in the temporal lobe. Medially, the greater part of the caudate nucleus abuts the thalamus, along a junction marked by a groove, the sulcus terminalis. The sulcus contains the stria terminalis, lying deep to the ependyma (see Fig. 5.3; Fig. 14.5). The stria terminalis forms one margin of the choroid fissure of the lateral ventricle; the hippocampal fimbria and fornix form the other margin. The sulcus terminalis is especially prominent anterosuperiorly (because of the large head and body of the caudate nucleus relative to the tail), where it is accompanied by the thalamostriate vein.
Fig. 14.4 Striatum within the left cerebral hemisphere.
(By permission from Crossman, A.R., Neary, D., 2000. Neuroanatomy, 2nd ed. Churchill Livingstone, Edinburgh.)
Fig. 14.5 Oblique section through the diencephalon and basal ganglia. A8, A9, A10, dopaminergic cell groups; CL, centrolateral nucleus of thalamus; CM, centromedian nucleus of thalamus; EX, external pallidal segment; H, H1, H2, subthalamic fields of Forel; ICV, internal cerebral veins in the transverse fissure; INT, internal pallidal segment; ITA, interthalamic adhesion; MFB, median forebrain bundle; SLIC, sublentiform internal capsule; 3, third ventricle.
The corpus callosum lies above the head and body of the caudate nucleus. The two are separated laterally by the fronto-occipital bundle and medially by the subcallosal fasciculus, a bundle of axons that caps the nucleus (see Fig. 14.5; Figs 14.6, 14.7). The caudate nucleus is largely separated from the lentiform complex by the anterior limb of the internal capsule (Figs 14.1, 14.6, 14.7). However, the inferior part of the head of the caudate becomes continuous with the most inferior part of the putamen immediately above the anterior perforated substance. This junctional region is sometimes known as the fundus striati (see Fig. 14.6). Variable bridges of cells connect the putamen to the caudate nucleus for most of its length. They are most prominent anteriorly, in the region of the fundus striati and the head and body of the caudate nucleus, where they break up the anterior limb of the internal capsule (see Figs 14.6, 14.7). In the temporal lobe, the anterior part of the tail of the caudate nucleus becomes continuous with the posteroinferior part of the putamen. The vast bulk of the caudate nucleus and putamen are often referred to as the dorsal striatum. A smaller inferomedial part of the rostral striatum is referred to as the ventral striatum and includes the nucleus accumbens.
Fig. 14.7 Posterior aspect of a coronal section through the anterior horn of the lateral ventricles.
The lentiform complex lies deep to the insular cortex, with which it is roughly coextensive, although the two are separated by a thin layer of white matter and the claustrum (see Fig. 14.2; Fig. 14.8). The claustrum splits the insular subcortical white matter to create the extreme and external capsules. The latter separates the claustrum from the putamen (see Figs 16.40, 14.1, 14.2, 14.8). The lentiform complex is separated from the caudate nucleus by the internal capsule.
Inferiorly, a little behind the fundus striati, the lentiform complex is grooved by the anterior commissure, which connects inferior parts of the temporal lobes and the anterior olfactory cortex of the two sides (see Fig. 14.6). The area above the commissure is referred to as the dorsal pallidum, and that below it is the ventral pallidum.
Striatum
Neuroactive chemicals, whether intrinsic or derived from afferents, are not distributed uniformly in the striatum. For example, serotonin and glutamic acid decarboxylase concentrations are highest caudally, whereas substance P, acetylcholine and dopamine are highest rostrally. However, there is a finer-grain neurochemical organization that informs the view of the striatum as a mosaic of islands or striosomes (sometimes referred to as patches), each 0.5 to 1.5 mm across, packed into a background matrix. Striosomes contain substance P and enkephalin. During development, the first dopamine terminals from the substantia nigra are found in striosomes. Although this exclusivity does not persist after birth, striosomes in the caudate nucleus still contain a higher concentration of dopamine than the matrix does. The latter contains acetylcholine and somatostatin and is the target of thalamostriate axons. Receptors for at least some neurotransmitters are also differentially distributed. For example, opiate receptors are found almost exclusively within striosomes, and muscarinic receptors predominantly so. Moreover, the distribution of neuroactive substances within the striosomes is not uniform. In humans, the striosome–matrix patchwork is less evident in the putamen, where it appears to consist predominantly of matrix, than it is in the caudate nucleus.
The major connections of the striatum are summarized in Figure 14.9. Although the connections of the dorsal and ventral divisions overlap, a generalization can be made: the dorsal striatum is predominantly connected with motor and associative areas of the cerebral cortex, whereas the ventral striatum is connected with the limbic system and orbitofrontal and temporal cortices. For both dorsal and ventral striata, the pallidum and substantia nigra pars reticulata are key efferent structures. The fundamental arrangement is the same for both divisions. The cerebral cortex projects to the striatum, which in turn projects to the pallidum and substantia nigra pars reticulata. From these, efferents leave to influence the cerebral cortex (either the supplementary motor area or the prefrontal and cingulate cortices via the thalamus) and the superior colliculus.
The striatum also receives afferents, which are more crudely spatially organized, from the polysensory intralaminar thalamus. The cerebelloreceptive nucleus centralis lateralis projects to the anterior striatum (especially the caudate nucleus), and the cerebello- and pallidoreceptive centromedian nucleus projects to the putamen.
The aminergic inputs to the caudate and putamen are derived from the substantia nigra pars compacta (dopaminergic group A9; Figs 14.10, 14.11), the retrorubral nucleus (dopaminergic group A8; see Fig. 14.5), the dorsal raphe nucleus (serotoninergic group B7; see Fig. 14.10) and the locus coeruleus (noradrenergic group A6). The nigrostriatal input is sometimes referred to as the ‘mesostriatal’ dopamine pathway. It reaches the striatum by traversing the H fields of the subthalamus. These aminergic inputs appear to modulate the responses of the striatum to cortical and thalamic afferent influences.
Fig. 14.11 Scheme of the organization of the substantia nigra in transverse section. Compare with Figure 14.10. Medially, there is no sharp distinction between dopaminergic cells projecting to the dorsal striatum (pars compacta, A9) and those projecting to the ventral striatum and limbic system (paranigral nucleus, A10). Dendrites of dopaminergic neurones intrude into the pars reticulata. Note the distinctive projection systems from the pars reticulata.
(From Paxinos (Ed.), 1990. The Human Nervous System, vol. 1, Webster. pp. 889–944. Modified with permission from Elsevier.)
Efferents from the striatum pass to both segments of the globus pallidus and to the substantia nigra pars reticulata, where they end in a topically ordered fashion. Fibres ending in either the lateral or medial pallidal segment originate from different striatal cells (Figs 14.9, 14.12). Those to the lateral pallidum come from neurones that co-localize GABA and enkephalin and give rise to the so-called indirect pathway. This name refers to the fact that these striatal neurones influence the activity of basal ganglia output neurones in the medial pallidum via the intermediary of the subthalamic nucleus. Other striatal neurones, which co-localize GABA and substance P/dynorphin, project directly to the medial pallidum and are therefore described as the direct pathway.
[/not-level-membership-for-basic-science-category]
