Chapter 9A Cartilage Biopsy Handling
Formation of the Synovial Joint
The understanding of embryonic tissue formation will enable us to understand and control the processes of repair and regeneration in adult tissue. The first sign of joint formation in the embryo is the appearance of an interzone where the cells gives rise to the articular layer of the future long bones. It has been unclear whether the interzone cells derive from transdifferentiation of local prechondrocytes into interzone cells or from migration of mesenchymal cells into the joint site, or a combination.1 The prechondrocytes in the interzone differentiate to early chondroblasts, thereafter becoming and remaining articular chondrocytes. However, recently studies have shred new light of the role of this population of cells expressing GDF5 (growth and differentiation factor 5; also known as CDMP-1 [cartilage-derived morphogenic protein 1]), a member of the transforming growth factor-beta family. The role of GDF5 expressing cells was demonstrated in ROSA-LacZ-reporter mice where GDF5 reporter expressing cells from the interzone from E13.5 were followed postnatally and remain present in structures that appear to arise from the interzone, namely the articular cartilage, synovium, and other joint tissues.2
The Adult Articular Hyaline Cartilage
The articular cartilage tissue is an avascular, noninnervated, and alymphatic tissue where the nutrition of the chondrocytes comes from passive diffusion. The only cell type in cartilage is the chondrocyte, constituting about 2% to 5% of the tissue. The function of chondrocytes is to build, maintain, and remodel the extracellular matrix composed of collagens, proteoglycans, noncollagenous proteins, and water. The articular cartilage has specialized load-bearing properties and the ability to withstand compressive, tensile, and shear forces because of the composition and structural integrity of its extracellular matrix (ECM), which consists of fibrillar and nonfibrillar macromolecules. The principal fibrillar component is collagen type II, which accounts for 90% to 95% of the collagen in articular cartilage; it forms a three-dimensional cross-linked network (together with smaller amounts of other minor collagens such as collagen types IX and XI). The collagen fibrils on the surface are oriented tangentially to create maximal strength, whereas deeper in the cartilage the fibrils are more vertically oriented. Collagen type X is exclusively produced by prehypertrophic and hypertrophic chondrocytes in the calcified layer.
Articular cartilage also consists of noncollagenous matrix proteins, which are important for the interaction and assembly of the various macromolecules. Cartilage oligomeric protein (COMP) is a glycoprotein belonging to the thrombospondin family, also named thrombospondin 5. COMP interacts with collagen types I, II and IX, and it has been used as a diagnostic marker in serum for the progress of matrix degradation.3
Depending on matrix composition and cellular appearance, the articular cartilage is divided into several zones with different functional roles: superficial, transitional, radial, and calcified zones. Facing the joint cavity is the tangential layer/superficial zone with small, flattened cells parallel to the surface and mainly collagen type I fibers arranged tangentially to the articular surface and small amounts of proteoglycans. The surface is covered with a thin sheet of fine fibrils and cells lubricated by a thin layer of synovial fluid, sometimes called the “lamina splendens.” The production of the lubricin protein providing the frictionless surface of articular cartilage is a specific property for the cells at the surface of cartilage.4 Below the superficial zone is the transitional zone with larger rounded chondrocytes arranged in groups and as single cells. The matrix is composed almost entirely of proteoglycan and aggrecan, and is rich in collagens. The collagen fibrils are more randomly arranged than in the superficial zone. This zone displays the typical morphological features of hyaline cartilage. The zone is followed by the radial zone of the articular cartilage, where the cell density is lowest and the large chondrocytes form radial columns and produce a matrix rich in proteoglycans, especially aggrecan. The content of collagen is low. The tidemark is the zone separating the cartilage from the underlying bone, and below the tidemark is the calcified cartilage layer, with chondrocytes demonstrating hypertrophic phenotype and a matrix rich in type X collagen but without proteoglycans.
The growth properties of chondrocytes from different layers are poorly studied. The superficial cells express markers of stem cells or progenitor cells (e.g., the notch receptor). Deeper layers are more differentiated but the cloning capacity of cells in the middle or deep layer is not negligible. Based on these uncertainties a full thickness cartilage biopsy is preferred. Several limiting factors are associated with the use of chondrocytes from osteoarthritis (OA) joints, including the number of cells that can be obtained from a diseased tissue, the capacity of cells to proliferate in vitro, and the responsiveness to growth factors necessary to trigger redifferentiation process. Chondrocyte numbers are decreased by 38% in OA cartilage as assessed histomorphometrically and via the number of isolated cells. Whereas the proliferative capacity of chondrocytes in OA cartilage is increased in vitro and may account for chondrocyte clustering observed in vivo, results in vitro are still inconclusive given that both lower and higher proliferative rates have been reported. Despite all the identified differences, recent data indicate that OA chondrocytes retain their differentiation potential upon isolation and proliferation in vitro.5 In the micromass pellet cultures, OA chondrocytes continued to proliferate for 14 days, thus increasing the pellet size in contrast to normal chondrocytes. The proteoglycan production was comparable to normal chondrocytes, and the collagen-rich matrix was present, although the total collagen was significantly lower. Additionally, in a 3D-scaffold based on hyaluronic acid, OA chondrocytes were also able to produce cartilage-specific matrix proteins. These results raise hope that despite their differences in comparison to normal chondrocytes, OA chondrocytes could be employed as a cell source for tissue engineering (TE) treatment providing that the disease can be controlled. Recent data using human chondrocytes from patients with the history of trauma demonstrated that cells exposed to a hyaluronan-based scaffold reduced apoptosis and decreased gene expression as well as secretion of degradation cytokines, namely, MMP-1, and MMP-13. At the same time, the expression of cartilage-specific genes SOX9, collagen type II, and aggrecan indicated differentiation toward chondrogenesis.6
Instructions for Cartilage Sampling
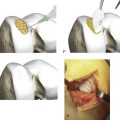
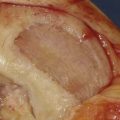
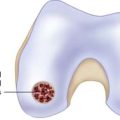
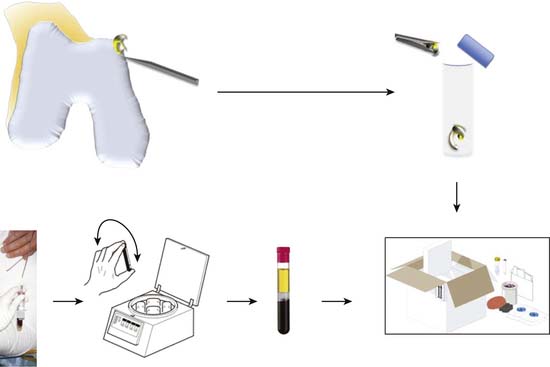
If required the intercondylar notch can be used.
Obtain a minimum amount of cartilage with a ring curette or a periosteal elevator.
The average weight recorded in our series (1235 biopsies) was 221 mg.
Leave a thin attachment with the neighboring cartilage.
Then use a grasper forceps to remove the biopsy through the portal.
Put the tube in the plastic cylinder.
Send the cartilage biopsy to the laboratory for isolation and expansion of chondrocytes.
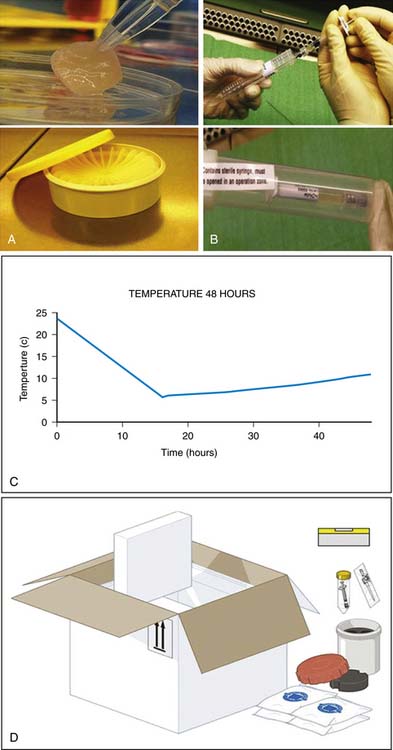
The chondrocytes can be frozen to be used later (Fig. 9A-1).
Autologous Blood Sampling
Instructions
Mark the tubes with the patient’s name.
Perform vein puncture in the usual way, and fill the tubes with blood samples.
Turn the tubes upside down five times so that the coagulation activator is mixed with the blood.
Let the blood coagulate at room temperature for 30 minutes with the tubes in a vertical position.
Centrifuge one more time if needed.
Put the tubes in the plastic cylinder for transportation (Fig. 9A-1).
Cell Seeded Scaffold
After expansion of the cells for two passages, the cells are seeded on human serum precoated Hyaff 11 scaffolds (thickness 2 and 4 mm;) using 2 × 106 cells per cm2. After incubation overnight at 37°C in 7% CO2/92% air, the scaffolds are cultured using chondrogenic media for 14 to 21 days (Fig. 9A-2, A-D).
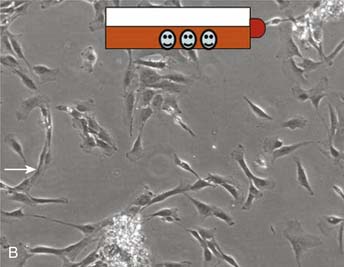

It is important to keep the membrane wet before implantation. Use 0.9% NaCl if necessary (Fig. 9A-3, A-D).
Cell Suspension Delivery
Let a nonsterile person open the outer container and drop the syringe onto the operating table (Fig. 9A-3).
1. Archer C.W., Dowthwaite G.P., Francis-West P. Development of synovial joints. Birth Defects Res C Embryo Today. 2003 May;69(2):144-155.
2. Koyama E., Shibukawa Y., Nagayama M., Sugito H., Young B., Yuasa T., Okabe T., Ochiai T., Kamiya N., Rountree R.B., Kingsley D.M., Iwamoto M., Enomoto-Iwamoto M., Pacifici M. A distinct cohort of progenitor cells participates in synovial joint and articular cartilage formation during mouse limb skeletogenesis. Dev Biol. 2008 Apr 1;316(1):62-73. Epub 2008 Jan 26
3. Saxne T., Heinegård D. Cartilage oligomeric matrix protein: a novel marker of cartilage turnover detectable in synovial fluid and blood. Br J Rheumatol. 1992 Sep;31(9):583-591. Erratum in: Br J Rheumatol 1993 Mar;32(3)
4. Jay G.D., Torres J.R., Warman M.L., Laderer M.C., Breuer K.S. The role of lubricin in the mechanical behavior of synovial fluid. Proc Natl Acad Sci USA. 2007 Apr 10;104(15):6194-6199. Epub 2007 Apr.
5. Tallheden T., Bengtsson C., Brantsing C., Sjögren-Jansson E., Carlsson L., Peterson L., Brittberg M., Lindahl A. Proliferation and differentiation potential of chondrocytes from osteoarthritic patients. Arthritis Res Ther. 2005;7(3):R560-R568. Epub 2005 Mar 3
6. Grigolo B., De Franceschi L., Roseti L., Cattini L., Facchini A. Down regulation of degenerative cartilage molecules in chondrocytes grown on a hyaluronan-based scaffold. Biomaterials. 2005 Oct;26(28):5668-5676. Epub 2005 Apr 12