CHAPTER 34 Microtubules and Centrosomes
Microtubules are stiff, cylindrical polymers of α- and β-tubulin (Fig. 34-1) that provide support for a variety of cellular components and tracks for movements powered by motor proteins called kinesins and dyneins. Microtubules are 25nm in diameter and can grow longer than 20μm in cells and 3mm in vitro. The head-to-tail arrangement of dimers of α- and β-tubulin in the wall of microtubules give the polymer a molecular polarity. The “plus” (β-tubulin) end grows faster than the “minus” end.
A great simplifying principle is that microtubules have a radial organization in many types of cells (Fig. 34-2A). Typically, the plus end is peripheral, and the minus end is anchored in a microtubule-organizing center. One exception to this radial organization is found in dendrites of nerve cells, where about 40% of the microtubules are oriented with the minus end away from the cell body.
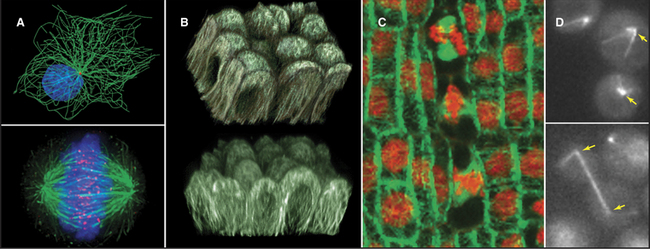
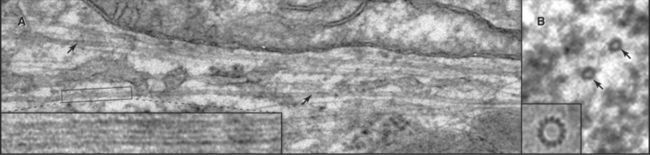
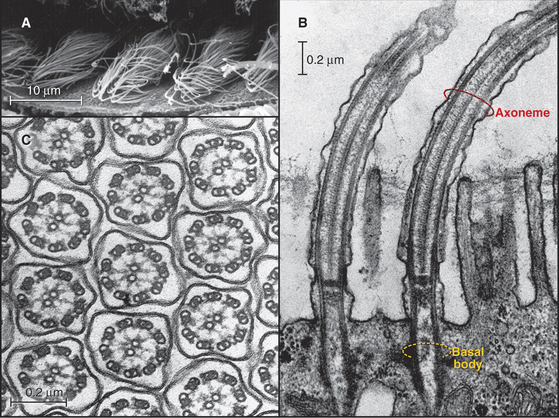
In most animal cells, the organizing center for cytoplasmic microtubules is the centrosome. Centrosomes consist of centrioles and a surrounding matrix containing the active component in microtubule nucleation—a complex of proteins including the specialized tubulin isoform γ-tubulin. The microtubule-organizing center is more diffuse in columnar epithelial cells, where microtubules originate from a broad zone containing γ-tubulin near the apex of the cell (Fig. 34-2B). In plant cells, γ-tubulin and microtubules are found throughout the cortex rather than in a discrete array, although they form a bipolar mitotic spindle during cell division (Fig. 34-2C). Microtubules in fungi grow from a spindle pole body, an organizing center containing γ-tubulin that is associated with the nuclear envelope (Fig. 34-2D). Animals and protozoa with cilia and flagella use basal bodies (Fig. 34-3B) to nucleate the assembly of microtubules for their motile structure, called an axoneme.
Microtubules vary considerably in stability. Those that form the axonemes in eukaryotic cilia and flagella are stable for days to weeks. Cytoplasmic microtubules turn over much more rapidly, within minutes in the case of the interphase array of microtubules and within tens of seconds for mitotic spindle microtubules. These dynamic microtubules randomly undergo rapid depolymerization and then regrow over a period of seconds to minutes. This “dynamic instability” helps to remodel the network of microtubules in cytoplasm and contributes to some forms of motility, including the assembly of the mitotic spindle and movements of chromosomes during mitosis (see Fig. 44-7).
Because the same tubulin dimers can form dynamic single microtubules in cytoplasm and stable doublet microtubules in axonemes, it is believed that accessoryproteins specify both the stability and diverse structures assembled from tubulin. The various families of microtubule-associated proteins (MAPs) bind tubulin dimers, stabilize polymers, associate with microtubule ends, or sever cytoplasmic microtubules. In the axonemes of cilia and flagella, more than 100 accessory proteins organize and stabilize the regular array of nine outer doublet microtubules and two central single microtubules (see Fig. 38-16).
Microtubule motor proteins (see Figs. 36-13 and 36-14) power movements ranging from the slow movements of chromosomes on the mitotic spindle (see Fig. 44-14) to the rapid beating of cilia and flagella (see Fig. 38-14). Different motors move toward the plus and minus ends of microtubules. The main minus-end-directed motor, dynein, drives the beating of cilia and flagella. Dynein and the kinesin family of plus-end motors move membrane-bound organelles, RNA particles, viruses, and other cargo along microtubules (see Fig. 37-7). These active movements determine, to a great extent, the distribution of cellular organelles and the shape of cells.
Tubulin Structure
The tubulin molecule is a heterodimer of a and β subunits that share a common fold and 40% identical residues (Fig. 34-4). Dimers of α- and β-tubulin are stable and rarely dissociate at the 10- to 20-mM concentr-tions of tubulin found in cells. γ-tubulins have the same fold.
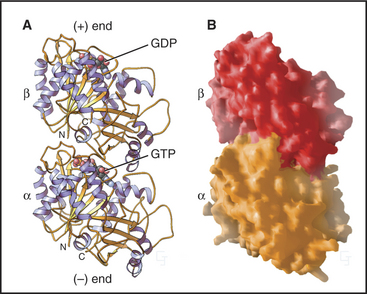
Each tubulin subunit binds a guanine nucleotide, either guanosine triphosphate (GTP) or guanosine diphosphate (GDP). The fold and the GTP-binding site of tubulin do not resemble those of other GTP-binding proteins (see Fig. 25-7). GTP on α-tubulin is buried in the dimer, so it does not exchange with solution GTP; hence, it is called the nonexchangeable N-site. GTP on β-tubulin is exposed in the dimer and exchanges slowly (Kd = 50nM), so this is known as the exchangeable site, or E-site. When incorporated into a microtubule, contacts with the adjacent a subunit bury the GTP on the β subunit and promote its hydrolysis. Neither bound guanine nucleotide can exchange when tubulin is buried in the wall of a microtubule. As is explained later in this chapter, the nature of the nucleotide on the β subunit profoundly affects microtubule assembly.
Most Archaea and Bacteria have a protein called FtsZ with the same fold as tubulin. The two proteins are most likely to have had a common ancestor, but their sequences have diverged considerably. FtsZ also forms polymers and is required for cytokinesis of prokaryotes (see Fig. 44-21). One group of Bacteria lost their FtsZ but have acquired genes for both α- and β-tubulin, most likely by lateral transfer from an eukaryote.
Tubulins are modified in a number of different ways in cells. Over time, stable microtubules accumulate two modifications—acetylation of lysine-40 and removal of the C-terminal tyrosine of α-tubulin—but neither modification is responsible for the stability. The enzyme carboxypeptidase B removes the tyrosine, leaving a glutamic acid exposed. Another enzyme, tyrosine-tubulin ligase, can replace the tyrosine. Other microtubules are modified by the addition of a polymer of up to six glutamic acid residues to the γ-carboxyl groups of glutamic acid residues of both α-tubulin and β-tubulin. Addition of one or more glycines to the γ-carboxyl of other glutamate residues stabilizes the central pair of microtubules in axonemes (see Fig. 38-16).
Structure of Microtubules
Microtubules are cylinders constructed of longitudin-ally oriented protofilaments with a 4-nm longitudinal repeat arising from the tubulin subunits (Fig. 34-5). Most cytoplasmic microtubules have 13 protofilaments, but microtubules in some cells have 11, 15, or 16 protofilaments. Microtubules assembled in vitro can have 11 to 15 protofilaments, but 13 is the favored number. For years, microtubules were thought to be true helices, but microtubules with 13 protofilaments have a longitudinal seam or discontinuity between two of the protofilaments, breaking up perfect helical packing of the subunits (Fig. 34-5D). The other protofilaments are aligned identically with respect to their neighbors.
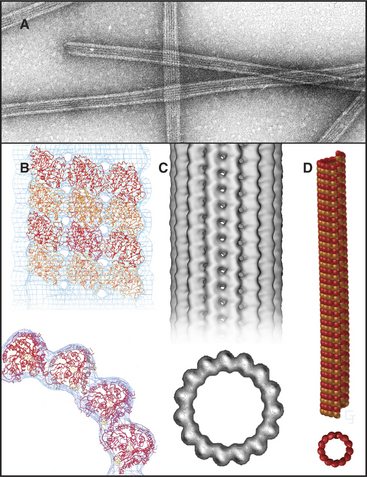
The stiffness, length, and polarity of microtubules make them valuable both for cytoskeletal support and as tracks for microtubule-based motors. Because they resist compression, microtubules are called on more frequently than actin filaments or intermediate filaments to support asymmetrical cellular structures, including axonemes, the mitotic spindle, and elaborate surface processes of some protozoa (see Fig. 38-4). Plants provide a spectacular example of the influence of microtubules on morphology: Point mutations in tubulin influence whether climbing plants wrap in a left-handed or right-handed helix around their supports.
Microtubule Assembly from GTP Tubulin
Microtubules assemble from pure GTP-tubulin subunits much like actin filaments do using subunits with bound ATP (see Figs. 5-6 and 33-8). Superficially, the assembly of microtubules is a simple bimolecular reaction of tubulin dimers with the ends of the polymer. Association and dissociation of tubulin occurs only at the ends, not from the walls of microtubules. Growth is faster at the plus end than at the minus end (Fig. 34-6), and assembly of GTP-tubulin is much more favorable than that of GDP-tubulin. The rate of elongation is proportional to the concentration of GTP-tubulin dimers above the critical concentration at each end. The rate constants measured from the slopes and intercepts in elongation experiments (Table 34-1) are similar to those for actin (see Fig. 33-8), although this similarity might be misleading, as the number of sites growing at the end of a microtubule is unknown and is most likely to be greater than one as on actin filaments.
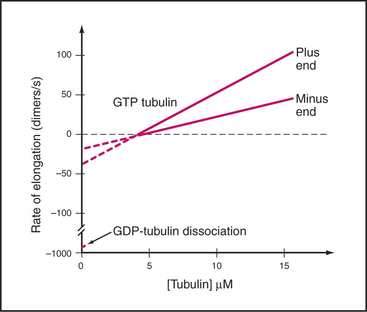
Table 34-1 RATE CONSTANTS FOR THE ASSEMBLY OF MICROTUBULES IN VITRO AND IN CELLS
| Reaction | Plus End | Minus End |
|---|---|---|
| In Vitro Elongation of Purified Tubulin | ||
| Association of GTP tubulin | 9 μM−1s−1 | 4 M−1s−1 |
| Dissociation of GTP tubulin | 44 s−1 | 23 s−1 |
| Association of GDP tubulin | Unknown | Unknown |
| Dissociation of GDP tubulin | 733 s−1 | 915 s−1 |
| Steady-State Dynamic Instability | ||
| Frequency of catastrophe in vitro at 7 μM tubulin | 0.0045 s−1 | 0.003 s−1 |
| Frequency of rescue in vitro at 7 μM tubulin | 0.02 s−1 | 0.06 s−1 |
| Microtubule Dynamic Instability in Live Cells | ||
| Frequency of catastrophe in vivo (interphase) | 0.014 s−1 | |
| Frequency of catastrophe in vivo (mitosis) | 0.017 s−1 | |
| Frequency of rescue in vivo (interphase) | 0.046 s−1 | |
| Frequency of rescue in vivo (mitosis) | 0 | |
Data from Walker RA, O’Brien ET, Pryer NK, et al: Dynamic instability of individual microtubules. J Cell Biol 107:1437–1448, 1988.
Drugs (Box 34-1) and environmental conditions can depolymerize microtubules. Pioneering light microscopic observations of live cells established that mitotic spindle fibers (later shown to be microtubules) are sensitive to both cold and high hydrostatic pressure. This makes it possible to purify microtubules and tightly associated proteins by cycles of depolymerization in the cold and repolymerization at higher temperatures. Dissociated microtubule components in ice-cold cellular extracts polymerize when rewarmed to body temperature. Pelleting in a centrifuge separates microtubules and any associated proteins from soluble components. Resuspension in cold buffer with GTP depolymerizes the microtubules. The cycle is repeated until the desired degree of purity is achieved. Adaptations of tubulins or accessory proteins allow cold-water organisms to assemble microtubules at temperatures near freezing.
Microtubule-Organizing Centers
Instead of spontaneous nucleation, virtually all cellular microtubules arise from microtubule-organizing centers. Microtubules in cilia and flagella grow directly from basal bodies (see Fig. 38-17), but other microtubules originate in the pericentrosomal material surrounding centrioles in the centrosome (Figs. 34-2A and 34-18), from fungal spindle pole bodies (Fig. 34-19) or nuclear envelopes and less-organized material in the cortex of some epithelial cells (Fig. 34-2B) and plant cells (Fig. 34-2C). As is described in the section on centrosomes later in this chapter, these organizing centers use assemblies of γ-tubulin to initiate microtubules.
Steady-State Dynamics of Microtubules in Vitro
If the GTP-tubulin reactions were all that contributed to assembly, microtubules would grow until the concentration of free tubulin dimers decreased to the critical concentration, after which polymers would be relatively stable, since the critical concentrations are similar at the two ends. Under these conditions, assembly at one end would be balanced by disassembly at the other. However, this is not what happens at steady state in either test tubes or cells. In vitro, the overall microtubule polymer and monomer concentrations are stable over time, but the microtubule number declines as some microtubules disappear, and the survivors grow longer. Direct observation by light microscopy (Fig. 34-7A)shows that this behavior is attributable to random, rapid fluctuations in the length of microtubules. Amazingly, growing and shrinking microtubules coexist at steady state; some shrinking microtubules disappear while others grow. This behavior is called dynamic instability.






